
植物生态学报 2010, 34 (1): 2–6 doi: 10.3773/j.issn.1005-264x.2010.01.002
Chinese Journal of Plant Ecology http:// www. https://www.doczj.com/doc/4a11865576.html,
——————————————————
收稿日期Received: 2009-11-24 接受日期Accepted: 2009-12-15
生态化学计量学: 探索从个体到生态系统的统一化理论
贺金生1,2 韩兴国3
1
北京大学城市与环境学院生态学系, 北京 100871; 2中国科学院西北高原生物研究所, 西宁 810008; 3
中国科学院植物研究所, 北京 100093
Ecological stoichiometry: Searching for unifying principles from individuals to ecosystems
HE Jin-Sheng 1,2 and HAN Xing-Guo 3
1
Department of Ecology, College of Urban and Environmental Sciences, Peking University, Beijing 100871, China; 2Northwest Institute of Plateau Biology, Chinese Academy of Sciences, Xining 810008, China; and 3Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China
从1909年丹麦哥本哈根大学Johannes Warming (1841–1924)出版第一本生态学教科书《植物生态学》到现在, 生态学经历了100年的发展。作为研究生物与生物、生物与环境相互关系的学科, 生态学具有高度的综合性和交叉性, 研究的问题也具有复杂性和多尺度的特点。正因为这样的学科特点, 传统上生态学家多强调研究对象的特殊性、研究问题的局域性, 缺乏统一的理论框架。进入21世纪以后, 生态学在统一化理论方面出现了一些尝试, 代表性的有生态学的代谢理论(metabolic theory of ecology) (Brown et al ., 2004)和生态化学计量学(ecological stoichiometry) (Sterner & Elser, 2002)。
生态化学计量学综合生物学、化学和物理学的基本原理, 利用生态过程中多重化学元素的平衡关系, 为研究C 、N 、P 等元素在生态系统过程中的耦合关系提供了一种综合方法。1958年, 哈佛大学的A. Redfield 首次证明: (1)海洋浮游生物的C 、N 、P 有特定的组成(摩尔比106:16:1, 该比率后被称为Redfield 比率); (2)这一比率受海洋环境和生物相互作用的调节(Redfield, 1958)。这一开创性的研究成为以后生态化学计量学的奠基之作。从Elser 和Hassett (1994)以及Elser 等(2000)的重要文章, 到以后Sterner 和Elser (2002)的专著《Ecological Stoichi- ometry 》的出版, 标志着生态化学计量学理论的系统化和逐步成熟。
2004年著名生态学杂志《Ecology 》 (85卷1 177页)和2005年《Oikos 》(109卷3页)分别出版了“Ecological stoichiometry”专辑, 进一步推动了这一领域的发展。本刊也于2005年发表了曾德慧和
陈广生(2005)的综述性文章, 系统介绍了生态化学计量学的理论与进展。
1 生态化学计量学为从个体到生态系统统一化理论的构建提供了新思路
生态化学计量学主要研究生态过程中化学元素的比例关系, 因此跨越了个体、种群、群落、生
态系统、景观和区域各个层次。生态化学计量学目前主要集中在C 、N 、P 元素的计量关系。这是因为, C 、N 、P 是重要的生命元素, 它们是地球上所有生命化学组成的基础。一般来说, 组成地球上有机体蛋白质的16%是N, 核酸组成的9.5%是P, 这两个比例在不同来源的生物中相对稳定。而有机体干物质的50%左右是C, 这一比例在生物的不同类群中随细胞的结构组成发生变化。生物在长期进化过程中, 形成了一定的内稳态机制(homeostatic mecha-nism), 即生物在变化的环境(包括食物)中具有保持其自身化学组成相对恒定的能力, 它是生态化学计量学存在的前提(Sterner & Elser, 2002)。现在还不清楚生物组成的其他元素是否存在相对稳定的计量关系, 但从生态化学计量学角度, 对其他化学元素的探索是必要的, 特别是S 的作用。
无论是植物个体水平, 还是生态系统水平, C 、N 、P 都是相互作用的。研究其中一个元素在生态学过程中的作用, 必须同时考虑其他元素的影响。生态化学计量学为研究C 、N 、P 等主要元素的生物地球化学循环和生态学过程提供了一种新思路 (Sterner & Elser, 2002; Güsewell, 2004)。在植物的个
贺金生等: 生态化学计量学: 探索从个体到生态系统的统一化理论 3
doi: 10.3773/j.issn.1005-264x.2010.01.002
体水平上, C 、N 、P 的组成及分配是相互联系、不可分割的一个整体, 它们的相互作用及与外界环境的关系共同决定着植物的营养水平和生长发育过程(Bazzaz & Grace, 1997; Güsewell, 2004)。通常可以把C 或N 作为植物资源分配的“货币(currency)”来看待, 不同器官和组织之间相互作用的结果就是分配多少C 或N 到特定部位, 以协调整体的生长发育过程(Grace, 1997)。例如, 植物的光合作用与光合器官(通常是叶片)中的N 含量密切相关(Field & Mooney, 1986), 而光合器官中的氮素又依赖于植物根系对N 的吸收和向叶片的运输, 这些过程都需要植物的光合作用提供能量。因此, 植物要获得C 首先需要投资N 到同化器官。同样, 为了获得N, 植物要投资同化的有机物到根系。在个体水平上, 植物的生长速率随叶片N:P 比率的降低而增加, 即所谓的生长速率假说(growth rate hypothesis, Sterner & El-ser, 2002)。
在生态系统水平上, 生产者、消费者、分解者及土壤等环境的C 、N 、P 组成决定了生态系统的主要过程, 如能量流动和物质循环(Chapin, 1980; Tilman, 1982; Aerts & Chapin, 2000; Moe et al ., 2005)。例如, 群落冠层叶片氮素水平在一定程度上代表其光合能力和生态系统的生产力, 凋落物的分解速率也与其C:N 比率呈负相关关系, 而土壤的C:N 比率与有机质的分解、土壤呼吸等密切相关 (Schlesinger & Andrews, 2000; Yuste et al ., 2007), 土壤及植物的N 和P 共同决定着生态系统的生产力(Treseder & Vitousek, 2001; Chapin et al ., 2002)。因此, 在生态系统水平上, C 、N 、P 的耦合作用制约了
生态系统的主要过程。
2 化学计量学是当前生态学研究的前沿领
域之一
化学计量学目前的发展主要集中在以下几个方面:
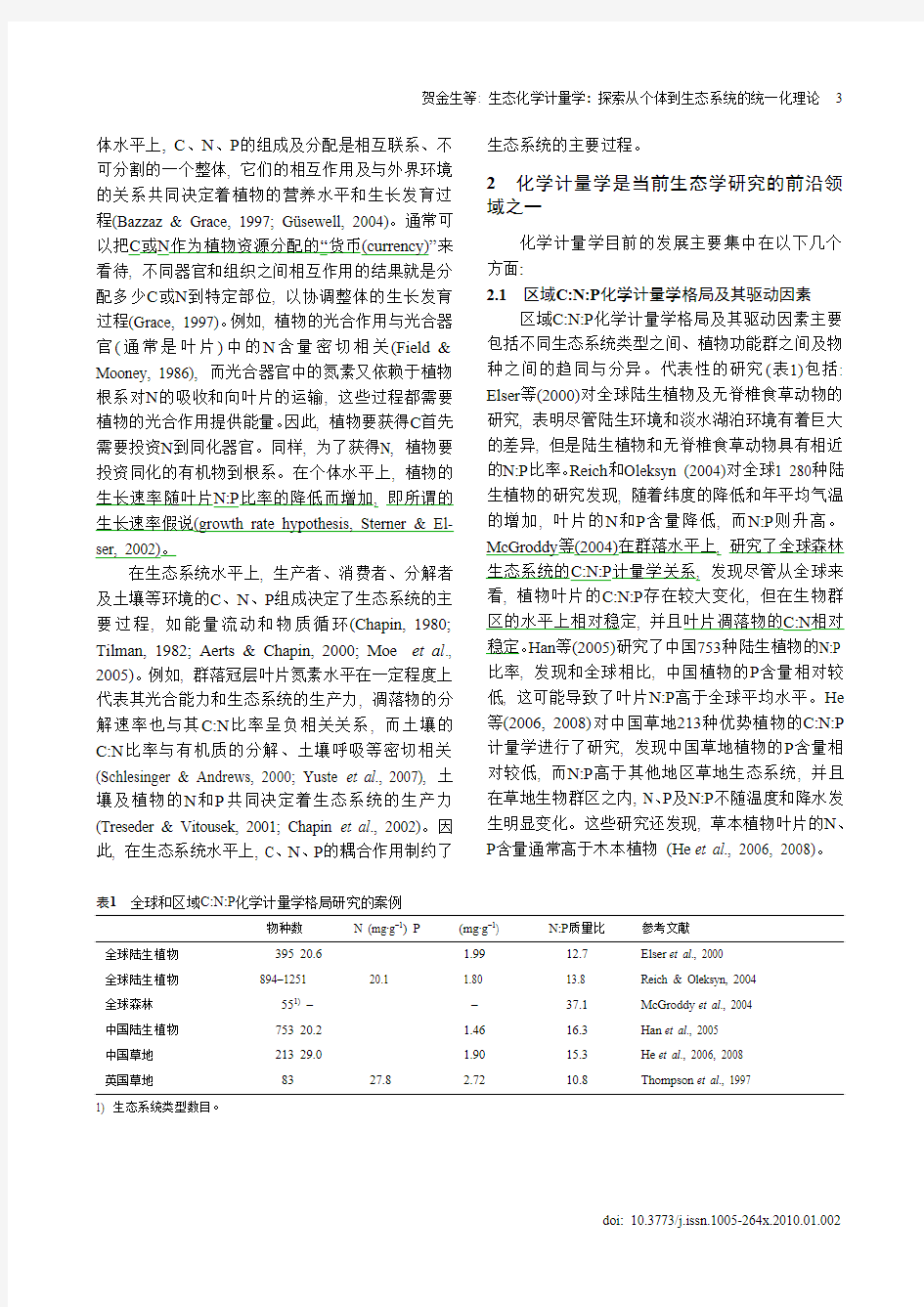
2.1 区域C:N:P 化学计量学格局及其驱动因素
区域C:N:P 化学计量学格局及其驱动因素主要包括不同生态系统类型之间、植物功能群之间及物种之间的趋同与分异。代表性的研究(表1)包括: Elser 等(2000)对全球陆生植物及无脊椎食草动物的研究, 表明尽管陆生环境和淡水湖泊环境有着巨大的差异, 但是陆生植物和无脊椎食草动物具有相近的N:P 比率。Reich 和Oleksyn (2004)对全球1 280种陆生植物的研究发现, 随着纬度的降低和年平均气温的增加, 叶片的N 和P 含量降低, 而N:P 则升高。McGroddy 等(2004)在群落水平上, 研究了全球森林生态系统的C:N:P 计量学关系, 发现尽管从全球来看, 植物叶片的C:N:P 存在较大变化, 但在生物群区的水平上相对稳定, 并且叶片凋落物的C:N 相对稳定。Han 等(2005)研究了中国753种陆生植物的N:P 比率, 发现和全球相比, 中国植物的P 含量相对较低, 这可能导致了叶片N:P 高于全球平均水平。He 等(2006, 2008)对中国草地213种优势植物的C:N:P 计量学进行了研究, 发现中国草地植物的P 含量相对较低, 而N:P 高于其他地区草地生态系统, 并且在草地生物群区之内, N 、P 及N:P 不随温度和降水发生明显变化。这些研究还发现, 草本植物叶片的N 、P 含量通常高于木本植物 (He et al ., 2006, 2008)。
表1 全球和区域C:N:P 化学计量学格局研究的案例
物种数
N (mg·g –1) P (mg·g –1) N:P 质量比 参考文献
全球陆生植物 395 20.6 1.99 12.7 Elser et al ., 2000 全球陆生植物 894–1251
20.1
1.80
13.8
Reich & Oleksyn, 2004
全球森林 551) – – 37.1
McGroddy et al ., 2004 中国陆生植物 753 20.2 1.46 16.3 Han et al ., 2005 中国草地 213 29.0 1.90 15.3 He et al ., 2006, 2008 英国草地
83
27.8
2.72
10.8
Thompson et al ., 1997
1) 生态系统类型数目。
4 植物生态学报Chinese Journal of Plant Ecology 2010, 34 (1): 2–6
https://www.doczj.com/doc/4a11865576.html,
2.2 C:N:P 计量关系与植物个体生长发育、种群增长、群落动态和生态系统过程的联系
生态化学计量学的基本原理之一, 就是种内不同发育阶段之间, 以及群落和生态系统不同组成物种之间对C 、N 、P 等多种元素要求的差异, 这种差异引起不同层次上资源“供应-需求”之间的错配(mismatch)或矛盾, 从而调节生理和生态学过程 (Anderson et al., 2004b; Moe et al., 2005)。例如植物和养分之间、食草动物和植物之间、食碎屑者和碎屑之间(detritivore vs. detritus)都可能发生错配现象, 因此可以影响到生态学的各个过程, 包括个体的生长(Elser et al., 1996; Anderson et al., 2004b; Vrede et al., 2004)、种群的增长(Urabe & Sterner, 1996)、元素循环和物种共存(Urabe et al., 2002; Hessen et al., 2004; Zhang et al., 2004)。到目前为止, 有关元素错配的生态学效应研究, 多集中在个体水平上的生理生态过程, 而对于种群和群落学的作用研究很少 (Anderson et al., 2004a; Frost et al., 2005)。 2.3 不同营养级之间对C:N:P 化学计量学的调节
对于生态系统的食物链来说, 这个过程包括食物的C:N:P 比率如何通过代谢过程, 传递(translate) 到下一营养级, 即在生态系统不同组分中的调节 (Sterner & Elser, 2002; Raubenheimer & Simpson, 2004)。具体到植物生态学, 相关研究主要集中到C:N:P 如何从高C:N 和C:P (土壤)到低C:N 和C:P (植物叶片和根系)的传递过程及其机理。在C:N:P 计量学研究中, 一个重要的例子就是可以根据植物叶片的N:P 比率来判断土壤养分状况。例如, Koerselman 和Meuleman (1996)通过对欧洲湿地植物施肥作用的评估发现, 当植物N:P < 14时, 表现为受N 的限制; 当N:P > 16时, 表现为受P 的限制。内蒙古羊草草原的施肥试验表明, N:P > 23时是P 限制, 而N:P < 21时是N 限制(Zhang et al., 2004)。最近通过对内蒙古温带草地、青藏高原高寒草地, 以及新疆山地草地199个取样地点213个物种的化学计量学分析发现, 植物叶片N:P 比率主要受P 含量的影响(He et al., 2006, 2008)。
2.4 环境要素和生物组成元素之间的计量关系
除了生物组成元素之间的计量关系, 目前发现环境要素如光照可能影响植物的C:N:P 计量关系。近年来的一些研究表明, “光:养分(light : nutrient)”
可能存在计量关系, 特别是在浮游生物中。如Striebel 等(2008)发现, 光照可以改变浮游植物的C:P 计量关系, 随着辐射增强, 藻类群落生物量和C:P 增长加快, 并且不管是贫营养还是富营养的湖泊中, 水蚤生物量的增长都是中等光强时最大。Dickman 等(2008)把光:养分比率关系应用到鱼类的实验中, 发现光、营养和食物链长度都能影响能量传递效率。这些研究表明, 除了考虑C:N:P 计量关系, 光照等环境要素也是一个重要因素。
3 本“生态化学计量学专题”旨在促进生态化
学计量学在国内的发展
生态化学计量学近年来在国内发展较快。这些研究主要集中在区域C:N:P 化学计量学格局及其驱动因素方面(Han et al., 2005; He et al., 2006; 任书杰等, 2007; Zheng & Shangguan, 2007; He et al., 2008), 也有施肥对群落N:P 比率的影响(Zhang et al., 2004), 不同演替阶段优势植物的N:P 比率的变化(高三平等, 2007)。另外, 曾德慧和陈广胜(2005)综述了生态化学计量学的研究进展, 王绍强和于贵瑞(2008)重点介绍了生态化学计量学在土壤学中的应用。
本专题收录了8篇论文, 分别对森林、草地和湿地生态系统的植物、土壤C:N:P 化学计量特征进行了研究。这些研究不仅包括了不同生态系统类型之间(阎恩荣等, 2010; 吴统贵等, 2010a, 2010b)、不同演替阶段(刘兴诏等, 2010; 银晓瑞等, 2010)植物C:N:P 化学计量特征的差异, 还包括了植物叶片N 、P 化学计量学特征的季节变化、植物叶片和细根不同器官之间计量特征的关联(周鹏等, 2010)以及这种关联在物种之间和物种内的差异(徐冰等, 2010)。以往的研究多集中在物种水平, 而杨阔等(2010)则探讨了群落水平的化学计量学特征及随环境的变化, 这无疑是以后研究的一个重要内容。
显然, 这些研究只是国内方兴未艾的生态化学计量学研究的一小部分, 仅能部分反映出国内外研究的热点和关键的科学问题。生态化学计量学是一种理论、一种思维, 也是一种工具, 对立志提高生态学的理论性和预测能力的研究者来说, 无疑是一个机遇, 但也是挑战。我们相信, 随着生态化学计量学渗透到生态学的各个方面, 一定会有更新的成
贺金生等: 生态化学计量学: 探索从个体到生态系统的统一化理论 5
doi: 10.3773/j.issn.1005-264x.2010.01.002
果不断出现。 参考文献
Aerts R, Chapin FS III (2000). The mineral nutrition of wild
plants revisited: a re-evaluation of processes and patterns. Advances in Ecological Research, 30, 1–67.
Anderson T, Elser JJ, Hessen DO (2004a). Stoichiometry and
population dynamics. Ecology Letters, 7, 884–900.
Anderson TR, Boersma M, Raubenheimer D (2004b). Stoichi-
ometry: linking elements to biochemicals. Ecology, 85, 1193–1202.
Bazzaz FA, Grace J (1997). Plant Resource Allocation . Aca-demic Press, San Diego, CA, USA.
Brown JH, Gillooly JF, Allen AP, Savage VM, West GB
(2004). Toward a metabolic theory of ecology. Ecology, 85, 1771–1789.
Chapin FS III (1980). The mineral nutrition of wild plants.
Annual Review of Ecology and Systematics, 11, 233–260. Chapin FS III, Matson PA, Mooney H (2002). Principles of
Terrestrial Ecosystem Ecology. Springer-Verlag, New York.
Dickman EM, Newell JM, González MJ, Vanni MJ (2008).
Light, nutrients, and food-chain length constrain plank-tonic energy transfer efficiency across multiple trophic levels. Proceedings of the National Academy of Sciences of the United States of America, 105, 18408–18412.
Elser JJ, Dobberfuhl DR, MacKay NA, Schampel JH (1996).
Organism size, life history, and N:P stoichiometry. Bio-Science, 46, 674–684.
Elser JJ, Fagan WF, Denno RF, Dobberfuhl DR, Folarin A,
Huberty A, Interlandi S, Kilham SS, McCauley E, Schulz KL, Siemann EH, Sterner RW (2000). Nutritional con-straints in terrestrial and freshwater food webs. Nature, 408, 578–580.
Elser JJ, Hassett RP (1994). A stoichiometric analysis of the
zooplankton-phytoplankton interaction in marine and freshwater ecosystems. Nature, 370, 211–213.
Field C, Mooney HA (1986). The photosynthesis-nitrogen rela-tionship in wild plants. In: Givnish TJ ed. On the Economy of Plant form and Function . Cambridge University Press, Cambridge, UK. 25–55.
Frost PC, Evans-White MA, Finkel ZV, Jensen TC, Matzek V
(2005). Are you what you eat? Physiological constraints on organismal stoichiometry in an elementally imbalanced world. Oikos, 109, 18–28. Gao SP (高三平), Li SX (李俊祥), Xu MC (徐明策), Chen X
(陈熙), Dai J (戴洁) (2007). Leaf N and P stoichiometry of common species in successional stages of the evergreen broad-leaved forest in Tiantong National Forest Park, Zhejiang Province, China. Acta Ecologica Sinica (生态学报), 27, 947–952. (in Chinese with English abstract) Grace J (1997). Towards models of resource allocation by
plants. In: Bazzaz FA, Grace J eds. Plant Resource Allo-cation . Academic Press, San Diego, CA, USA. 279–291. Güsewell S (2004). N:P ratios in terrestrial plants: variation
and functional significance. New Phytologist, 164, 243– 266.
Han W, Fang JY, Guo D, Zhang Y (2005). Leaf nitrogen and
phosphorus stoichiometry across 753 terrestrial plant spe-cies in China. New Phytologist, 168, 377–385.
He JS, Fang JY, Wang ZH, Guo D, Flynn DFB, Geng Z
(2006). Stoichiometry and large-scale patterns of leaf car-bon and nitrogen in the grassland biomes of China. Oecologia, 149, 115–122.
He JS, Wang L, Flynn DFB, Wang X, Ma W, Fang J (2008).
Leaf nitrogen : phosphorus stoichiometry across Chinese grassland biomes. Oecologia, 155, 301–310.
Hessen DO, Agren GI, Anderson TR, Elser JJ, de Ruiter PC
(2004). Carbon, sequestration in ecosystems: the role of stoichiometry. Ecology, 85, 1179–1192.
Koerselman W, Meuleman AFM (1996). The vegetation N:P
ratios: a new tool to detect the nature of nutrient limita-tion. Journal of Applied Ecology, 33, 1441–1450. Liu XZ (刘兴诏), Zhou GY (周国逸), Zhang DQ (张德强),
Liu SZ (刘世忠), Chu GW (褚国伟), Yan JH (闫俊华) (2010). N and P stoichiometry of plant and soil in lower subtropical forest successional series in southern China. Chinese Journal of Plant Ecology (植物生态学报), 34, 64–71. (in Chinese with English abstract)
McGroddy ME, Daufresne T, Hedin LO (2004). Scaling of
C:N:P stoichiometry in forests worldwide: implications of terrestrial Redfield-type ratios. Ecology, 85, 2390–2401. Moe SJ, Stelzer RS, Forman MR, Harpole WS, Daufresne T,
Yoshida T (2005). Recent advances in ecological stoichiometry: insights for population and community ecology. Oikos, 109, 29–39.
Raubenheimer D, Simpson SJ (2004). Organismal stoichiome-try: quantifying non-independence among food compo-nents. Ecology, 85, 1203–1216.
Redfield AC (1958). The biological control of chemical factors
in the environment. American Scientist, 46, 205–221.
Reich PB, Oleksyn J (2004). Global patterns of plant leaf N and
P in relation to temperature and latitude. Proceedings of the National Academy of Sciences of the United States of America, 101, 11001–11006. Ren SJ (任书杰), Yu GR (于贵瑞), Tao B (陶波), Wang SQ
(王绍强) (2007). Leaf nitrogen and phosphorus stoichio- metry across 654 terrestrial plant species in NSTEC. Chi-nese Journal of Environmental Science (环境科学), 28, 2665–2667.
Schlesinger WH, Andrews JA (2000). Soil respiration and the
global carbon cycle. Biogeochemistry, 48, 7–20.
Sterner RW, Elser JJ (2002). E cological Stoichiometry: the
Biology of Elements from Molecules to the Biosphere.
6 植物生态学报Chinese Journal of Plant Ecology 2010, 34 (1): 2–6
https://www.doczj.com/doc/4a11865576.html,
Princeton University Press, Princeton.
Striebel M, Sp?rl G, Stibor H (2008). Light-induced changes of
plankton growth and stoichiometry: experiments with natural phytoplankton communities. Limnology and Oceanography, 53, 513–522.
Thompson K, Parkinson JA, Band SR, Spencer RE (1997). A
comparative study of leaf nutrient concentrations in a re-gional herbaceous flora. New Phytologist, 136, 679–689. Tilman D (1982). Resource Competition and Community
Structure . Princeton University Press, Princeton.
Treseder KK, Vitousek PM (2001). Effects of soil nutrient
availability on investment in acquisition of N and P in Hawaiian rain forests. Ecology, 82, 946–954.
Urabe J, Elser JJ, Kyle MM, Sekino T, Kawabata Z (2002).
Herbivorous animals can mitigate unfavourable ratios of energy and material supplies by enhancing nutrient recy-cling. Ecology Letters, 5, 177–185.
Urabe J, Sterner RW (1996). Regulation of herbivore growth
by the balance of light and nutrients. Proceedings of the National Academy of Sciences of the United States of America, 93, 8465–8469.
Vrede T, Dobberfuhl DR, Kooijman S, Elser JJ (2004). Fun-damental connections among organism C:N:P stoichiome-try, macromolecular composition, and growth. Ecology, 85, 1217–1229. Wang SQ (王绍强), Yu GR (于贵瑞) (2008). Ecological
stoichiometry characteristics of ecosystem carbon, nitro-gen and phosphorus elements. Acta Ecologica Sinica (生态学报), 28, 3937–3947.
Wu TG (吴统贵), Chen BF (陈步峰), Xiao YH (肖以华), Pan
YJ (潘勇军), Chen Y (陈勇), Xiao JH (萧江华) (2010a). Leaf stoichiometry of trees in three forest types in Pearl River Delta, South China. Chinese Journal of Plant Ecology (植物生态学报), 34, 58–63. (in Chinese with English abstract)
Wu TG (吴统贵), Wu M (吴明), Liu L (刘丽), Xiao JH (萧江
华) (2010b). Seasonal variations of leaf nitrogen and phosphorus stoichiometry of three herbaceous species in Hangzhou Bay coastal wetlands, China. Chinese Journal of Plant Ecology (植物生态学报), 34, 23–28. (in Chinese with English abstract)
Xu B (徐冰), Cheng YX (程雨曦), Gan HJ (甘慧洁), Zhou WJ
(周文嘉), He JS (贺金生) (2010). Correlations between leaf and fine root traits among and within species of typical temperate grassland in Xilin River Basin, Inner Mongolia, China. Chinese Journal of Plant Ecology (植物生态学报), 34, 29–38. (in Chinese with English abstract) Yan ER (阎恩荣), Wang XH (王希华), Guo M (郭明), Zhong
Q (仲强), Zhou W (周武) (2010). C:N:P stoichiometry across evergreen broad-leaved forests, evergreen coniferous forests and deciduous broad-leaved forests in the Tiantong region, Zhejiang Province, eastern China. Chinese Journal of Plant Ecology (植物生态学报), 34, 48–57. (in Chinese with English abstract)
Yang K (杨阔), Huang JH (黄建辉), Dong D (董丹), Ma WH
(马文红), He JS (贺金生) (2010). Canopy leaf N and P stoichiometry in grassland communities of Qinghai- Tibetan Plateau, China. Chinese Journal of Plant Ecology (植物生态学报), 34, 17–22. (in Chinese with English abstract)
Yin XR (银晓瑞), Liang CZ (梁存柱), Wang LX (王立新),
Wang W (王炜), Liu ZL (刘钟龄) , Liu XP (刘小平) (2010). Ecological stoichiometry of plant nutrients at different restoration succession stages in typical steppe of Inner Mongolia, China. Chinese Journal of Plant Ecology (植物生态学报), 34, 39–47. (in Chinese with English abstract)
Yuste JC, Baldocchi DD, Gershenson A, Goldstein A, Misson
L, Wong S (2007). Microbial soil respiration and its dependency on carbon inputs, soil temperature and moisture. Global Change Biology, 13, 1–8. Zeng DH (曾德慧), Chen GS (陈广生) (2005). Ecological
stoichiometry: a science to explore the complexity of living systems. Acta Phytoecologica Sinica (植物生态学报), 29, 1007–1019. (in Chinese with English abstract) Zhang LX, Bai YF, Han XG (2004). Differential responses of
N:P stoichiometry of Leymus chinensis and Carex korshinskyi to N additions in a steppe ecosystem in Nei Mongol. Acta Botanica Sinica, 46, 259–270.
Zheng S, Shangguan Z (2007). Spatial patterns of leaf nutrient
traits of the plants in the Loess Plateau of China. Trees, 21, 357–370. Zhou P (周鹏), Geng Y (耿燕), Ma WH (马文红), He JS (贺金
生) (2010). Linkages of functional traits among plant organs in the dominant species of the Inner Mongolia grassland, China. Chinese Journal of Plant Ecology (植物生态学报), 34, 7–16. (in Chinese with English abstract)
责任编辑: 王 葳
植物生态学报 2010, 34 (1): 7–16 doi: 10.3773/j.issn.1005-264x.2010.01.003
Chinese Journal of Plant Ecology https://www.doczj.com/doc/4a11865576.html,
——————————————————
收稿日期Received: 2008-05-05 接受日期Accepted: 2008-07-23 * 通讯作者Author for correspondence (E-mail: jshe@https://www.doczj.com/doc/4a11865576.html,)
温带草地主要优势植物不同器官间功能性状的关联
周 鹏1 耿 燕1 马文红2 贺金生1*
1
北京大学城市与环境学院生态学系, 北京 100871; 2北京大学深圳研究生院环境与城市学院, 深圳 518055
摘 要 理解植物各器官间功能性状的关联, 有助于确定控制功能性状的内在机制以及性状间的比例关系。基于内蒙古温带草地19个地点、42种优势草本植物的野外观测, 分析了叶片、茎、生殖器官、细根和粗根间功能性状(N 、P 含量、N:P 、比叶面积、比根长以及叶片和细根的组织密度)的关联。主要结果如下: 在种群和物种水平上, 各器官的N 和P 含量都显著正相关, 比叶面积与叶片N 、P 含量和组织密度在种群水平上显著负相关, 而在物种水平上没有显著的相关关系; 而比根长仅在种群水平上与细根的组织密度显著负相关。N 、P 含量以及N:P 在各器官之间一致呈显著正相关, 而比叶面积和比根长没有显著的相关关系。叶片和细根的组织密度在种群水平上显著负相关, 而在物种水平上没有显著的相关关系。非禾草比禾草相应器官(除茎外)的N 、P 含量高, 但二者茎的N 、P 含量没有显著的差异; 豆科植物比非豆科植物相应器官的N 含量高, 而P 含量没有显著的差异。 关键词 生态计量学, 植物器官, 比叶面积, 比根长, 温带草地, 组织密度
Linkages of functional traits among plant organs in the dominant species of the Inner Mongo-lia grassland, China
ZHOU Peng 1, GENG Yan 1, MA Wen-Hong 2, and HE Jin-Sheng 1*
1
Department of Ecology, College of Urban and Environmental Sciences, Peking University, Beijing 100871, China; and 2School of Environment & Urban Study, Peking University Shenzhen Graduate School, Shenzhen 518055, China
Abstract
Aims The temperate grasslands in Inner Mongolia, representing a great diversity in vegetation types (desert steppe, typical steppe and meadow steppe) and function groups (grass/herb, legume/non-legume), are ideal places to test the hypothesized functional trait relationships among plant organs. Our main objective in this study was to test whether plant functional traits vary in a coordinated fashion both within and across organs.
Methods Based on the field observation during July and August in both 2006 and 2007, we measured suites of ecophysiological traits of 42 grassland species from 19 sites in Inner Mongolia. The longitude of the study region ranges from 112.82° to 120.12° (E), and the latitude ranges from 41.76° to 49.89° (N). N and P concentrations, N:P ratios of leaves, stems, reproductive structures, fine roots (diameter < 1 mm) and coarse roots (diameter > 1 mm) as well as tissue density of leaf and fine root and specific leaf area/specific root length were determined. Important findings At both population and interspecific level, N and P concentrations were positively correlated within each organ. Specific leaf area was negatively correlated with leaf N and P concentrations and tissue density at the population level but not at the interspecific level. Specific root length was negatively related to fine root tissue density at population level. Plants with low leaf or fine root tissue density had leaves or fine roots with high N concentrations and large specific leaf area or specific root length. N and P concentrations as well as N: P ratios were also consistently correlated across all organs, but no correlation between specific leaf area and specific root length was observed. At the population level there existed a weak negative correlation between leaf and fine root tissue density while at interspecific level this relationship disappeared. Grasses had lower N and P concentrations than herbs in leaves, reproductive structures and roots, but not in stems. Legumes had higher N concentrations than non-legumes in all organs, but they showed no significant differences in P concentrations.
Key words ecological stoichiometry, plant organ, specific leaf area, specific root length, temperate grassland, tissue density
阐明不同器官结构和功能属性的关系是理解物种性状演化的关键(Eviner & Chapin, 2003), 这有
助于确定控制功能性状的内在机制以及性状间的比例关系(Tjoelker et al ., 2005), 并将在很大程度上
8 植物生态学报Chinese Journal of Plant Ecology 2010, 34 (1): 7–16
https://www.doczj.com/doc/4a11865576.html,
改进全球生态系统机理模型(Norby & Jackson, 2000; Reich et al ., 2001; Westoby et al ., 2002)。植物功能性状的变化通常被描述为主要器官(叶片、茎、根和生殖结构)间营养元素分配及形态结构的差异(Westoby et al ., 2002; Wright et al ., 2004)。不同器官的功能、生长和周转速率以及植物生活史对策与养分含量密切相关(Jackson et al ., 1997)。同时, 单个器官并不是严格地独立于其他器官(Ackerly & Donoghue, 1998), 其功能性状可能受到器官间生理作用和系统发育的共同影响(Kerkhoff et al ., 2006)。
近年来研究表明, 植物不同器官内功能性状表现趋同。例如, 植物叶片(Han et al., 2005; He et al., 2008)、根(Kerkhoff et al., 2006)的N 和P 含量都正相关; 叶片N 含量与比叶面积(单位质量的叶面积, SLA)正相关(Reich et al., 1997; Wright et al ., 2004; Wright & Westoby, 2004; He et al ., 2006a), 而与叶片的组织密度负相关(Craine et al., 2005); 根N 含量与比根长(单位质量的根长, SRL)正相关(Pregitzer et al., 1998; Reich et al., 1998), 而与根的组织密度负相关(Craine et al., 2005)。同样, 植物不同器官间功能性状表现出一致的联系。例如, 叶和根的N 含量(Craine & Lee, 2003; Craine et al ., 2005; Tjoelker et al ., 2005)、P 含量(Kerkhoff et al., 2006)以及组织密度(Craine & Lee, 2003)都正相关。并且, 有研究表明上述关系不随物种(Kerkhoff et al., 2006)、土壤(Craine et al ., 2001)以及气候(Ryan, 1991; Wright et al ., 2004)的变化而变化。
目前, 有关植物功能性状的研究大多集中在光合器官(Reich et al ., 1997; Wright et al ., 2004; Wright & Westoby, 2004; He et al ., 2006a, 2006b), 近年来逐渐深入到地下部分(Pregitzer et al ., 1998, 2002; Eissenstat et al ., 2000; Gordon & Jackson, 2000; Wahl & Ryser, 2000), 但很少有人考虑到植物不同器官间功能性状的联系。Kerkhoff 等(2006)研究了152科1 287种草本和木本植物, 发现N 、P 含量和N:P 在不同器官(叶片、茎、生殖结构和根)内、之间一致相关。然而, 国际上至今仍缺乏在生态系统水平上的大量野外观测数据(张林和罗天祥, 2004)来定量描述植物不同器官间功能性状的相关关系。目前国内有关植物功能属性在不同器官间的关联的研究基本尚属空白。
内蒙古温带草原跨越了多种草地类型(荒漠草
原、典型草原和草甸草原), 为研究不同功能群(如多年生禾草/非禾草, 豆科/非豆科)植物结构和功能的联系提供了丰富的资源。我们将植物器官具体划分如下: 叶片、茎、生殖结构、细根(直径 < 1 mm)和粗根(直径 > 1 mm)。通过分析上述器官间功能性状(N 、P 含量, N:P 以及叶片和细根的组织密度, 比叶面积/比根长)的数量关系, 试图揭示: 1)温带草地草本植物各功能性状在不同器官内是否具有一致的关系; 2)这些性状在不同器官间是否存在一致的相关关系。
1 数据和方法
1.1 样地概况
2006–2007年7–8月, 在内蒙古草原沿着气候梯度从西向东进行野外采样, 研究区的范围为112.82°–120.12° E, 41.76°–49.89° N, 年平均气温为–2.5– 4.2 ℃, 年降水量为141–429 mm (表1)。研究区自西向东分布着3种草地类型: 荒漠草原、典型草原和草甸草原。其中, 荒漠草原优势物种为小针茅(Stipa klemenzii )和沙生针茅(S. glareosa ), 并生长有葱属(Allium )植物, 如蒙古葱(A. mongolicum )和多根葱(A. polyrhizum ); 典型草原优势物种为大针茅(S. grandis )和羊草(Leymus chinensis ); 草甸草原优势物种为贝加尔针茅(S. baicalensis )和线叶菊(Filifolium sibiricum ), 并生长有多年生杂草, 如柴胡(Bupleurum chinense )、扁蓿豆(Melilotoides ruthe- mica )和地榆(Sanguisorba officinalis )。 1.2 取样方法
沿着温度和水分梯度在研究区选择人类活动影响较小的19个样地: 荒漠草原6个样地, 典型草原7个样地, 草甸草原6个样地。一共调查了42个物种, 隶属于13科27属。每个样地选取优势物种, 每种植物取5–10株。在每株植物的茎基部10–20 cm 范围内用铁铲挖取20–30 cm 深的土块, 取出包括根在内的完整植株, 小心清理掉根表面的土壤和杂质。取样过程中尽量避免末端根的损失, 以保证根系构型的完整性。将取出的完整植株放入装有冰块的冷贮藏箱内(<5 ℃), 临时保存并迅速运回实验室。
从每种植物摘取5–10片成熟完整的叶片, 用电子游标卡尺测量叶厚度, 并用便携式叶面积仪
(AM200, ADC Bioscientific Limited, Herts, UK)测定
周鹏等: 温带草地主要优势植物不同器官间功能性状的关联 9
doi: 10.3773/j.issn.1005-264x.2010.01.003
表1 研究区概况
Table 1 Study site descriptions
地点 Site
经度
Longitude (° E)
纬度
Latitude (° N)
海拔 Altitude (m)
草地类型 Grassland type
1 120.20 49.35 669 草甸草原 Meadow steppe
2 119.99 49.89
730
草甸草原 Meadow steppe
3 119.56 46.58 1 213
草甸草原 Meadow steppe 4 118.66 46.53 997
草甸草原 Meadow steppe 5 118.66 45.75 879 典型草原 Typical steppe 6 118.18 46.15
970
典型草原 Typical steppe
7 118.11 44.36 1 136 草甸草原 Meadow steppe 8 117.36 44.51 1 062 典型草原 Typical steppe 9 117.13 45.32 865 典型草原 Typical steppe 10 116.82 43.51
1 433 草甸草原 Meadow steppe 11 116.74 43.60 1 213 典型草原 Typical steppe
12 116.74 43.60 1 217 典型草原 Typical steppe 13 116.28 44.07 1 075 典型草原 Typical steppe 14 113.40 43.81
1 003
荒漠草原 Desert steppe
15 112.73 42.95 1 037 荒漠草原 Desert steppe 16 112.59 42.84 1 087 荒漠草原 Desert steppe 17 112.23 43.31 1 018 荒漠草原 Desert steppe 18 111.99 43.61
943
荒漠草原 Desert steppe
19 111.82 41.76
1 435 荒漠草原 Desert steppe
叶面积后, 将叶片在60 ℃下烘干48–72 h 后称重, 得到比叶面积(叶面积/质量, cm 2·g –1)和组织密度(质量/总体积, g·cm –3)。然后, 将叶片、茎和生殖器官从每株植物分离出来, 在60 ℃下烘干48–72 h 后称重。
根系样品在实验室冷冻( < –15 ℃)保存。将植物根上附着的土用水冲洗干净并分离出细根(直径 < 1 mm)和粗根(直径 > 1 mm)。选取其中25个优势物种(每种植物5株)的细根, 用数字图像分析系统(WinRhizo, Régent Instruments, Inc. Quebec City, Quebec, Canada)扫描, 获得细根的总长度和总体积。然后, 将细根在60 ℃下烘干48–72 h 后称重, 得到比根长(总长度/质量, m·g –1)和组织密度(质量/总体积, g·cm –3)。
烘干的各器官样品粉碎后, 用C/N 元素分析仪(2400II CHNS/O Elemental Analyzer, Perkin-Elmer, USA)测定总N 含量, 并用磷钼蓝比色法测定总P 含量(Kuo, 1996)。 1.3 数据分析
首先, 基于单因素方差分析(One-Way ANO- V A), 比较了不同草地类型和功能群之间植物各器
官的功能性状的差异; 其次, 用Pearson 相关分析在种群(n = 106)和物种(n = 42)水平上检验各器官内、之间功能性状的相关关系。上述统计分析在统计软件SPSS 13.0 (2001, v. 13.0; SPSS Inc., USA)下完成。最后, 应用统计软件SMATR 2.0 (Standardized Ma-jor Axis Tests & Routines Version 2.0)在上述两个水平上对叶片和细根的功能性状进行了II 类线性回归分析。原始数据经对数变换以符合标准化正态分布。
2 结果
2.1 不同功能群和草地类型植物各器官功能性状的比较
内蒙古温带草地植物各器官的N 、P 含量和N:P 在不同功能群以及草地类型之间基本上呈一致的格局(图1)。非禾草比禾草相应器官(叶片、生殖结构、细根和粗根)具有更高的N 、P 含量(p < 0.05), 而茎的N 、P 含量在禾草和非禾草之间没有显著的差异(p > 0.05)。豆科比非豆科相应器官具有更高的N 含量(p < 0.05), 而P 含量没有显著的差异(p > 0.05)。与典型草原和草甸草原相比, 荒漠草原植物叶片和生
10 植物生态学报Chinese Journal of Plant Ecology 2010, 34 (1): 7–16
https://www.doczj.com/doc/4a11865576.html,
图1 内蒙古温带草地不同功能群和草地类型植物叶片(Le)、茎(St)、生殖器官(Re)、细根(Fr)和粗根(Cr)平均N 含量(A )、P 含量(B )、N:P (C )、比叶面积和比根长(D )以及叶片和细根的组织密度(E )的比较。用单因素双差分析(Tukey post hoc)检验不同功能群和草地类型之间的差异; 显著性差异(p < 0.05)用不同字母标记。误差棒为标准误差。
Fig. 1 Comparisons of leaf (Le), stem (St), reproductive structure (Re), fine root (Fr) and coarse root (Cr), mean N concentrations (A ), P concentrations (B ), N: P ratios (C ), specific leaf area, specific root length (D ) and tissue density of leaves and fine roots (E ) among different function groups and vegetation types in Inner Mongolia grassland. Differences between each group were tested us-ing a One-Way ANOV A with a Tukey post hoc test of significance; significant differences at p < 0.05 are indicated by different letters. Errors bars are standard errors. SLA, specific leaf area; SRL, specific root length.
殖结构的N 、P 含量更高, 而叶片、细根和粗根的N:P 在3种草地类型间没有表现出明显的变化格局(p > 0.05)。荒漠草原的比叶面积更低(p < 0.05), 不同植被类型的比根长没有差异(p > 0.05)。 2.2 不同器官功能性状的关联
在种群和物种水平上, 所有植物不同器官(叶
片、茎、生殖结构、细根和粗根)的N 和P 含量都显著正相关(表2)。比叶面积与叶片N 、P 含量和组织密度在种群水平上显著负相关, 而在物种水平上没有显著的相关关系; 在种群和物种水平上, 比叶面积与叶片的N:P 都没有显著的相关关系(图2)。除了在种群水平上比根长与细根的组织密度显著负相
周鹏等: 温带草地主要优势植物不同器官间功能性状的关联 11
doi: 10.3773/j.issn.1005-264x.2010.01.003
表2 内蒙古温带草地不同器官内功能性状的相关系数
Table 2 Pearson correlation coefficients for functional traits within different organs in Inner Monglolia grassland
种群水平 Population level
物种水平 Interspecific level N P
N: P
SLA/SRL
N P
N: P
SLA/SRL
叶片 P 0.79 P 0.81 Leaf
N : P
0.19
– 0.46 N: P 0.31 – 0.30 SLA – 0.21 – 0.30 0.19 SLA – 0.22 – 0.26 0.07 TD – 0.21 – 0.24 0.08 – 0.37 TD – 0.08 – 0.23 0.26 – 0.35 茎 P 0.73 P 0.77 Stem N : P 0.50 – 0.23 N: P 0.59 – 0.06 生殖器官
P 0.87 P 0.83 Reproductive structure N: P 0.24 – 0.27 N: P 0.43 – 0.15 细根 P 0.69 P 0.69 Fine root N: P 0.60 – 0.14 N: P 0.65 – 0.01 SRL – 0.03
– 0.07 – 0.04 SRL – 0.14 – 0.04 – 0.20 TD – 0.35
– 0.13 – 0.41 – 0.29 TD – 0.41 – 0.18 – 0.42 – 0.14 粗根 P 0.46 P 0.49 Coarse root
N: P
0.58
– 0.43
N: P
0.64
– 0.35
SLA, 比叶面积; SRL, 比根长; TD, 组织密度; Re: 生殖器官。黑体为显著相关(p < 0.05)。
SLA, specific leaf area; SRL, specific root length; TD, tissue density. Bold coefficients denote statistical significance (p < 0.05).
关外, 比根长与细根的N 、P 含量、N:P 和组织密度都没有显著的相关关系(图3)。
在种群和物种水平上, 除了细根和茎的P 含量在物种水平上没有显著的相关关系外, N 、P 含量以及N:P 在所有植物不同器官间都显著正相关(表3)。此外, 叶片和细根的组织密度在种群水平上显著负相关(图4B), 而在物种水平上没有显著的相关关系(图4D); 比叶面积和比根长在两个水平上都没有显著的相关关系(图4A 、C)。
3 讨论
基于内蒙古温带草地19个地点、42种优势草本植物的野外观测, 我们发现各器官的N 和P 含量正相关, N 、P 含量以及N:P 在不同器官间也一致正相关, 这与Kerkhoff 等(2006)的结果一致。而叶片和细根的组织形态(比叶面积/比根长、组织密度)与化学组成(N 、P 含量和N:P)之间并没有一致的关系。
本研究中, 叶片的平均N 、P 含量分别为26.4和1.7 mg·g –1, 与He 等(2006b, 2008)对中国草地的研究结果(分别为27.6、1.9 mg·g –1)接近。而Thompson 等(1997)研究得出英国草地的N 、P 含量分别为27.8和2.7 mg·g –1, 可见中国草地叶片的P 含量普遍较低(He et al ., 2006b), 导致研究区内N:P (16.5)高于全
球植被叶片的平均水平(13.8) (Reich & Oleksyn, 2004)。与地上器官相比, 关于植物根系功能性状(尤其是化学组成)的研究较少, 从而限制了我们对植物整体适应环境策略的内在机制的认识。我们得出细根的N 、P 含量分别为10.9和0.7 mg·g –1, 与全球植被细根的平均N 、P 水平(分别为11.7和1.1 mg·g –1) (Jackson et al ., 1997)相比, N 含量相近而P 含量低。同时, 细根、粗根、茎的N 、P 含量低于叶片和生殖结构, 细根N 、P 含量还不到叶片和生殖器官(分别为23.5和2.38 mg·g –1)的1/2。对于不同的功能群, 除茎的N 、P 含量在禾草和非禾草之间没有显著的差异, 其他各器官(叶片、生殖结构、细根和粗根)的N 、P 含量都表现为: 非禾草>禾草; 与非豆科相比, 豆科植物相应器官具有更高的N 含量, 而P 含量没有显著的差异。植物功能性状的变异反映了组织结构和功能的关键平衡(Tilman et al ., 1997), 从而在组织水平上, 禾草比非禾草、非豆科植物比豆科植物具有更高的养分利用效率(Tjoelker et al ., 2005)。
在不同器官内, N 、P 含量之间均正相关。这一方面因为有机体合成蛋白质用于生长, 需要消耗大量的三磷酸腺苷(ATP) (Chapin, 1980), 另一方面也反映了环境供应的养分元素的共变性(Güsewell & Koerselman, 2002; Sterner & Elser, 2002)。此外, 细
12 植物生态学报Chinese Journal of Plant Ecology 2010, 34 (1): 7–16
https://www.doczj.com/doc/4a11865576.html,
图2 内蒙古温带草地所有植物比叶面积(SLA)和叶片的N 含量(A , E )、P 含量(B , F )、N:P (C , G )、组织密度(D , H )在两个水平上的相关关系(II 类线性回归)。A –D , 种群水平; E –H , 物种水平, 用物种平均值。
Fig. 2 Relationships between specific leaf area (SLA) and leaf N concentrations (A , E ), P concentrations (B , F ), N: P ratios (C , G ), tissue density (D , H ) at two levels across all species in Inner Mongolia grassland (type II linear regression). A –D , population level; E –H , interspecific level, using species means.
根的N:P 受N 限制, 这与叶片的N:P 受P 限制的格局相反。一般而言, 植物的N:P 受P 限制, 主要是因为环境供应的可供植物直接吸收和利用的活性P 比N 更少(Güsewell, 2004)。与叶片相比, 根系大多与真菌或其他有机体共生(Gordon & Jackson, 2000), 这些微生物为了自身的繁殖和生长, 通常会保留大部分从分解基质中获得的N, 而N 从有机物转化为生物可利用状态的过程落后于植物凋落物的分解过程(曾德慧和陈广生, 2005)。因此, 根系往往容易受
到N 不足的限制。在不同器官间, N 、P 含量以及N:P 也一致正相关。在植物生长发育过程中, 尽管结构型(粗根和茎)和新陈代谢型(叶片、生殖结构和细根) 器官具有不同的功能, 养分含量在不同功能群植物主要器官间却表现出一致的关系。
叶片和细根的组织形态(比叶面积/比根长、组织密度)与化学组成(N 、P 含量和N:P)之间并没有一致的关系。Reich 等(1997)和He 等(2006a)发现植物随比叶面积增大, 叶片N 含量增多。而本研究中比叶
周鹏等: 温带草地主要优势植物不同器官间功能性状的关联 13
doi: 10.3773/j.issn.1005-264x.2010.01.003
图3 内蒙古温带草地所有植物比根长(SRL)和细根的N 含量(A , E )、P 含量(B , F )、N:P (C , G )、组织密度(D , H )在两个水平上的相关关系(II 类线性回归)。A –H 同图2。
Fig. 3 Relationships between specific root length (SRL) and fine root N concentrations (A , E ), P concentrations (B , F ), N: P ratios (C , G ), tissue density (D , H ) at two levels across all species in Inner Mongolia grassland (type II linear regression). A –H see Fig. 2.
面积与叶片N 含量没有一致的关系, 这可能是由于适应极端环境(如干旱)的物种往往表现出储存更多N 、P 等养分的能力(Chapin et al ., 1990), 从而与典型草原和草甸草原相比, 荒漠草原植物虽然比叶面积更小, 但叶片的N 、P 含量却更高。同时, 我们发现细根N 含量和比根长并不相关。Tjoelker 等(2005)对草本植物细根N 含量与比根长的研究也得出相同的结论。而以往的研究大多认为, 比根长与根系养分吸收以及根的寿命有关(Eissenstat et al ., 2000), 如Pregitzer 等(1998)和Reich 等(1998)的研究表明木本
植物比根长越大, 细根N 含量越多。另一方面, 叶片和细根的组织形态之间也没有一致的相关关系。比叶面积与比根长不相关, 这与Craine 等(2001)和Tjoelker 等(2005)的结果一致。叶片和细根的组织密度也没有一致性。Craine 和Lee (2003)发现新西兰30个地点、24种草地植物叶片组织密度的变化只解释了细根组织密度变化的9%, 而Craine 等(2005)的研究表明, 全球4个草地区域植物叶片和细根的组织密度没有显著的相关关系。植物根系具有很多分支结构, 不同级别的根在结构和功能上差异较大(Guo
14 植物生态学报Chinese Journal of Plant Ecology 2010, 34 (1): 7–16
https://www.doczj.com/doc/4a11865576.html,
表3 内蒙古温带草地N 、P 含量和N:P 在不同器官间的相关系数
Table 3 Pearson correlation coefficients for N and P concentrations and N:P ratios among different organs in Inner Monglolia grass-land
种群水平 Population level 物种水平 Interspecific level
叶片
Leaf
茎 Stem
生殖器官 Re
细根 Fine root
叶片 Leaf
茎 Stem
生殖器官 Re
细根 Fine root
N
茎 Stem 0.75 0.82 生殖器官 Re 0.80 0.62
0.81 0.62
细根 Fine root 0.65 0.41 0.53 0.72 0.49 0.55 粗根 Coarse root
0.58 0.54 0.38 0.70
0.58 0.51 0.35 0.72
P 茎 Stem 0.67 0.73
生殖器官 Re 0.80 0.57 0.66 0.40 细根 Fine root 0.58 0.32 0.47 0.50 0.24 0.36 粗根 Coarse root 0.58 0.45 0.55 0.75 0.51 0.31 0.36 0.69 N: P
茎 Stem 0.69 0.74
生殖器官 Re 0.75 0.73 0.82 0.70 细根 Fine root 0.42 0.55 0.53 0.48 0.54 0.60 粗根 Coarse root
0.46 0.71 0.54
0.64
0.54 0.70 0.52
0.74
黑体为显著相关(p < 0.05)。
Bold coefficients denote statistical significance. Re, reproductive structure.
图4 内蒙古温带草地所有植物比叶面积和比根长(SRL) (A , C )、叶片和细根的组织密度(B , D )在两个水平上的相关关系(II 类线性回归)。A , B , 种群水平; C , D , 物种水平。
Fig. 4 Relationships between specific leaf area and specific root length (SRL) (A , C ), tissue density (B , D ) of leaves and fine roots, at two levels across all species in Inner Mongolia grassland (type II linear regression). A , B , Population level; C , D , Interspecific level, using species means.
周鹏等: 温带草地主要优势植物不同器官间功能性状的关联 15
doi: 10.3773/j.issn.1005-264x.2010.01.003
et al ., 2004), 从而可能导致根系与叶片的结构特征
不平行。尽管草本植物没有木本植物的根系分支多、结构复杂, 但并不排除上述原因可能引起草本植物根系与叶片性状产生差异。
植物养分含量及在各个器官间的分配既受生长地点养分有效性的制约, 同时也受植物自身生长型、生理特征和生活史的影响, 是环境和物种系统发育共同作用的结果(Aerts & Chapin, 2000)。生态和进化的动态过程决定了物质和能量在植物体内的流通和转化形式(Kerkhoff et al ., 2006)。我们的结果表明尽管不同的功能群(如禾草/非禾草, 豆科/非豆科)在器官元素含量上存在差异, 但功能性状的关联, 特别是N 和P 呈现出一致的格局。特定的环境选择了植物特定的生长策略。植物为了适应环境中养分的限制状况而调整不同器官的养分含量及其化学计量比值, 如Reich 和Oleksyn (2004)发现在较广的纬度梯度上, 叶片的N:P 可以反映土壤养分有效性。这种适应在不同功能群属性的植物中表现出一致性; 另一方面, 植物养分含量的变异在很大程度上可以通过系统发育来解释(Broadley et al ., 2004; He et al ., 2008), 也是物种水平上的重要性状。植物不同器官间养分分配形式及相应功能性状的关联特征将植物及其生存环境从生态和进化的角度联系起来(Kay et al ., 2005), 为进一步研究植物功能多样性、养分收支以及建立生态机理模型提供依据。
4 结论
本研究得出在种群和物种水平上, N 、P 含量以及N:P 在各器官内、之间一致正相关。叶片和细根的组织形态(比叶面积/比根长、组织密度)与化学组成(N 、P 含量和N:P)之间并没有一致的关系; 叶片和细根的组织形态之间也没有表现出一致性。究竟这些规律和结论在其他生态系统类型及更大的尺度上是否成立, 植物器官的养分含量及其化学计量比值、功能性状间的关系与土壤的性质联系如何, 以及物种对全球变化如何响应都将是今后研究的重要方面。
致谢 国家自然科学基金(90411004和30670322)资助项目。 参考文献
Ackerly DD, Donoghue MJ (1998). Leaf size, sapling allome-try, and Corner’s rules: phylogeny and correlated evolu-
tion in maples (Acer ). American Naturalist, 152, 767–791. Aerts R, Chapin FS III (2000). The mineral nutrition of wild
plants revisited: a reevaluation of processes and patterns. Advances in Ecological Research, 30, 1–67.
Broadley MR, Bowen HC, Cotterill HL, Hammond JP,
Meacham MC, Mead A, White PJ (2004). Phylogenetic variation in the shoot mineral concentration of angio-sperms. Journal of Experimental Botany, 55, 321–336. Chapin FS III (1980). The mineral nutrition of wild plants.
Annual Review of Ecology and Systematics, 11, 233–260. Chapin FS III, Schulze ED, Mooney HA (1990). The ecology
and economics of storage in plants. Annual Review of Ecology and Systematics, 21, 423–447.
Craine JM, Froehle J, Tilman DG, Wedin DA, Chapin FS III
(2001). The relationships among root and leaf traits of 76 grassland species and relative abundance along fertility and disturbance gradients. Oikos, 93, 274–285.
Craine JM, Lee WG (2003). Covariation in leaf and root traits
for native and non-native grasses along an altitudinal gra-dient in New Zealand. Oecologia, 134, 471–478.
Craine JM, Lee WG, Bond WJ, Williams RJ, Johnson LC
(2005). Environmental constraints on a global relationship among leaf and root traits of grasses. Ecology, 86, 12–19. Eissenstat DM, Wells CE, Yanai RD, Whitbeck JL (2000).
Building roots in a changing environment: implications for root longevity. New Phytologist, 147, 33–42.
Eviner VT, Chapin FS III (2003). Functional matrix: a concep-tual framework for predicting multiple plant effects on ecosystem processes. Annual Review of Ecology , Evolu-tion and Systematics, 34, 455–485.
Gordon WS, Jackson RB (2000). Nutrient concentrations in
fine roots. Ecology, 81, 275–280.
Guo DL, Mitchell RJ, Hendricks JJ (2004). Fine root branch
orders respond differentially to carbon source sink ma-nipulations in a longleaf pine forest. Oecologia, 140, 450–457.
Güsewell S, Koerselman W (2002). Variation in nitrogen and
phosphorus concentrations of wetland plants. Perspectives in Plant Ecology, Evolution and Systematics, 5, 37–61. Güsewell S (2004). N:P ratios in terrestrial plants: variation and
functional significance. New Phytologist, 164, 243–266. Han WY, Fang JY, Guo DL, Zhang Y (2005). Leaf nitrogen
and phosphorus stoichiometry across 753 terrestrial plant species in China. New Phytologist, 168, 377–385.
He JS, Wang ZH, Wang XP, Schmid B, Zuo WY, Zhou M,
Zheng CY, Wang MF, Fang JY (2006a). A test of the generality of leaf trait relationships on the Tibetan Plateau. New Phytologist, 170, 835–848.
He JS, Fang JY, Wang ZH, Guo DL, Flynn DFB, Geng Z
(2006b). Stoichiometry and large-scale patterns of leaf carbon and nitrogen in the grasslands of China. Oecologia, 149, 115–122.
He JS, Wang L, Flynn DFB, Wang XP, Ma WH, Fang JY
16 植物生态学报Chinese Journal of Plant Ecology 2010, 34 (1): 7–16
https://www.doczj.com/doc/4a11865576.html,
(2008). Leaf nitrogen : phosphorus stoichiometry across
Chinese grassland Biomes. Oecologia, 155, 301–310.
Jackson RB, Mooney HA, Schulze ED (1997). A global budget
for fine root biomass, surface area and nutrient contents. Proceedings of the National Academy of Science of the United States of America, 94, 7362–7366.
Kay AD, Ashton IW, Gorokhova E, Kerkhoff AJ, Liess A,
Litchman E (2005). Toward a stoichiometric framework for evolutionary biology. Oikos, 109, 6–17.
Kerkhoff AJ, Fagan WF, Elser JJ, Enquist BJ (2006). Phyloge-netic and growth form variation in the scaling of nitrogen and phosphorus in the seed plants. American Naturalist, 168, E103–E122.
Kuo S (1996). Phosphorus. In: Bigham JM ed. Methods of Soil
Analysis. Part 3. Chemical Methods . Soil Science Society of America, American Society of Agronomy, Madison, Wis. 869–919.
Norby RJ, Jackson RB (2000). Root dynamics and global
change: seeking an ecosystem perspective. New Phytolo-gist, 147, 3–12.
Pregitzer KS, Laskowski MJ, Burton AJ, Lessard VC, Zak DR
(1998). Variation in sugar maple root respiration with root diameter and soil depth. Tree Physiology, 18, 665–670. Pregitzer KS, DeForest JL, Burton AJ, Allen MF, Ruess RW,
Hendrick RL (2002). Fine root architecture of nine North American trees. Ecological Monographs, 72, 293–309. Reich PB, Walters MB, Ellsworth DS (1997). From tropics to
tundra: global convergence in plant functioning. Proceed-ings of the National Academy of Sciences of the United States of America, 94, 13730–13734.
Reich PB, Walters MB, Tjoelker MG, Vanderklein D,
Buschena C (1998). Photosynthesis and respiration rates depend on leaf and root morphology and nitrogen concen-tration in nine boreal tree species differing in relative growth rate. Functional Ecology, 12, 395–405.
Reich PB, Tilman D, Craine J, Ellsworth D, Tjoelker MG,
Knops J, Wedin D, Naeem S, Bahauddin D, Goth J, Beng-ston W, Lee TD (2001). Do species and functional groups differ in acquisition and use of C, N and water under varying atmospheric CO 2 and N availability regimes? A field test with 16 grassland species. New Phytologist, 150, 435–448.
Reich PB, Oleksyn J (2004). Global patterns of plant leaf N and
P in relation to temperature and latitude. Proceedings of the National Academy of Sciences of the United States of
America, 101, 11001–11006.
Ryan MG (1991). Effects of climate change on plant respira-tion. Ecological Applications, 1, 157–167.
Sterner RW, Elser JJ (2002). Ecological Stoichiometry : the
Biology of Elements from Molecules to the Biosphere . Princeton University Press, Princeton, NJ.
Tilman D, Knops J, Wedin D (1997). The influence of func-tional diversity and composition on ecosystem processes. Science, 277, 1300–1302.
Tjoelker MG, Craine JM, Wedin D, Reich PB, Tilman D
(2005). Linking leaf and root trait syndromes among 39 grassland and savannah species. New Phytologist, 167, 493–508.
Thompson K, Parkinson JA, Band SR, Spencer RE (1997). A
comparative study of leaf nutrient concentrations in a re-gional herbaceous flora. New Phytologist, 136, 679–689. Wahl S, Ryser P (2000). Root tissue structure is linked to eco-logical strategies of grasses. New Phytologist, 148, 459–471.
Westoby M, Falster DS, Moles AT, Vesk PA, Wright IJ (2002).
Plant ecological strategies: some leading dimensions of variation between species. Annual Review of Ecology and Systematics, 33, 125–159.
Wright IJ, Reich PB, Westoby M, Ackerly DD, Baruch Z,
Bongers F, Cavender-Bares J, Chapin T, Cornelissen JHC, Diemer M, Flexas J, Garnier E, Groom PK, Gulias J, Hi-kosaka K, Lamont BB, Lee T, Lee WJ, Lusk C, Midgley JJ, Navas ML, Niinemets U, Oleksyn J, Osada N, Poorter H, Poot P, Prior L, Pyankov VI, Roumet C, Thomas SC, Tjoelker MG, Veneklaas EJ, Villar R (2004). The world-wide leaf economics spectrum. Nature, 428, 821–827. Wright IJ, Westoby M (2004). The economics of leaves: plants
build leaves as investments that vary in cost, revenue and lifespan. Australasian Science, 25, 34–37. Zhang L (张林), Luo TX (罗天祥) (2004). Advances in eco-logical studies on leaf lifespan and associated leaf traits. Journal of Plant Ecology (Chinese version ) (植物生态学报), 28, 844–852. (in Chinese with English abstract) Zeng DH (曾德慧), Chen GS (陈广生) (2005). Ecological
stoichiometry: a science to explore the complexity of living systems. Journal of Plant Ecology (Chinese version ) (植物生态学报), 29, 1007–1019. (in Chinese with English abstract)
责任编委: 马克平 责任编辑: 姜联合
植物生态学报 2010, 34 (1): 17–22 doi: 10.3773/j.issn.1005-264x.2010.01.004 Chinese Journal of Plant Ecology https://www.doczj.com/doc/4a11865576.html,
青藏高原草地植物群落冠层叶片氮磷化学计量学分析
杨阔1,2黄建辉1*董丹1,2马文红3,4贺金生4
1中国科学院植物研究所植被与环境变化国家重点实验室, 北京 100093; 2中国科学院研究生院, 北京 100049; 3内蒙古大学生命科学学院, 呼和浩特010021; 4北京大学城市与环境学院生态学系, 北京 100871
摘要叶片氮(N)和磷(P)的化学计量学研究涉及到植物生态学的众多领域与多个尺度, 然而各个尺度上的化学计量学研究并未同步展开。通过对青藏高原47个草地样地连续3年的调查, 分析了当地群落水平上的植物叶片N、P含量及其化学计量学特征, 并结合温度和降水气候数据研究了N、P含量及N:P比值与这两个气候因子的相关关系。研究结果显示: 青藏高原草地群落水平的叶片N含量变化范围为14.8–36.7 mg·g–1, 平均为23.2 mg·g–1; P含量变化范围为0.8–2.8 mg·g–1, 平均为1.7 mg·g–1; N:P比值变化范围为6.8–25.6, 平均为13.5。群落叶片N含量与P含量呈显著正相关关系, 叶片的N:P比值与P含量呈显著负相关关系, N:P比值的变化主要由P含量变化决定。另外发现: 群落水平叶片N、P含量及N:P比值存在着显著的年际变化, 叶片的N、P含量及N:P比值与年平均气温之间存在着极显著的相关关系。通过该研究结果推测: P含量较高的变异系数及其与环境因子表现出的显著相关性, 在一定程度上体现了植物群落对当地气候条件的一种适应。
关键词草原群落, 叶片N、P含量, N:P比, 青藏高原
Canopy leaf N and P stoichiometry in grassland communities of Qinghai-Tibetan Plateau, China YANG Kuo1,2, HUANG Jian-Hui1*, DONG Dan1,2, MA Wen-Hong3,4, and HE Jin-Sheng4
1State Key Laboratory of Vegetation and Environmental Change, Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China; 2Graduate Univer-sity of Chinese Academy of Sciences, Beijing 100049, China; 3College of Life Sciences, Inner Mongolia University, Hohhot 010021, China; and 4Department of Ecology, College of Urban and Environmental Sciences, Peking University, Beijing 100871, China
Abstract
Aims Leaf N and P stoichiometry has been widely studied at the species level in both aquatic and terrestrial ecosystems, however, it lacks research at the community level. Since the ecological stoichiometric characteristics could play important roles in connecting different levels of ecological studies and former studies mainly focused on the individual level, in this study, we try to figure out the pattern of foliar N and P at the community level of grassland ecosystems in Qinghai-Tibetan Plateau. Additionally, we also try to find out the relationships between community level leaf N, P and site climate factors.
Methods Leaf samples were collected from 47 research sites in Qinghai-Tibetan Plateau at the end of the grow-ing season yearly from 2006 to 2008. We measured the leaf N concentrations by using an elemental analyzer and the leaf P concentration based on a molybdate/stannous chloride method. Climate data of annual mean tempera-ture and annual mean precipitation (65 national standard stations) between 2006 and 2008 were used to interpolate into gridded data with a resolution of 1 km × 1 km through the tchebycheffian spline function.
Important findings Leaf N, P concentrations and N:P ratios at the community level over the southern part of Qinghai-Tibetan Plateau were 23.2 mg·g–1, 1.7 mg·g–1 and 13.5, respectively. Significant inter-annual differences were presented in leaf N, P concentrations and N:P ratios. Mean annual temperature was strongly correlated with leaf N, P and N:P ratios. Besides, the correlations between climate factors and leaf N, P, N:P ratios were generally consistent with the previous results found at the global scale. Our results suggest that the high variation in leaf P concentration and its strong correlation with environmental factors reveal that, to some extent, stoichiometric traits at the community level are adaptive to local environmental conditions.
Key words grassland community, leaf N and P concentrations, N:P ratios, Qinghai-Tibetan Plateau
——————————————————
收稿日期Received: 2009-11-17 接受日期Accepted: 2009-12-11
* 通讯作者Author for correspondence (E-mail: jhhuang@https://www.doczj.com/doc/4a11865576.html,)
18 植物生态学报Chinese Journal of Plant Ecology 2010, 34 (1): 17–22
https://www.doczj.com/doc/4a11865576.html,
氮(N)和磷(P)是植物的基本营养元素, 它们的循环限制着生态系统中的大多数过程(Chapin, 1980; Aerts & Chapin, 2000)。N 和P 也是各种蛋白质和遗传物质的重要组成元素。由于自然界中N 和P 元素供应往往受限, 成为生态系统生产力的主要限制因素(Vitousek & Howarth, 1991; Elser et al., 2007)。因此, 研究N 、P 在植物群落中的含量和分布格局十分必要。由于N 、P 元素在陆地生态系统中有着紧密的交互作用(Vitousek, 1982; Chapin et al.,
2002), 群体水平的N 、P 含量及其分布特征可能 对我们了解整个生态系统对N 、P 的需求更加重 要。
生态化学计量学的发展为解决上述问题提供了有力的工具, 通过化学计量学方法研究N 、P 的区域分布规律成为近年来的研究热点(Zhang et al., 2003)。植物叶片中的N 、P 含量与N:P 比值的化学计量学特征, 以及这些因子如何限制植物的生长已经被广泛研究(Thompson et al., 1997; Braakhekke & Hooftman, 1999; Reich et al., 1999; Elser et al., 2000; Wright et al., 2004; He et al., 2006)。另外, 一些研究着眼于揭示种质、气候因子对叶片N 、P 含量及N:P 比值的影响(K?rner, 1989; Reich & Oleksyn, 2004; Reich et al., 2006; He et al., 2008)。我国对中国植物叶片N 、P 含量及N:P 比值的研究则显示了一些不同于全球格局的分布特征(Han et al., 2005)。然而, 这些陆地生态系统的生态计量学研究, 主要是针对植物个体水平的调查以及物种水平的研究, 基于群落水平的研究报道非常少(McGroddy et al., 2004; Kerkhoff & Enquist, 2006)。而植物在群落水平上是否存在和植物个体类似的生态计量关系, 目前尚不清楚(G üsewell & Koerselman, 2002)。这一问题关系到能否将不同植物个体的特征和生理生态功能与群落以及生态系统的结构、动态相连接, 统一各个尺度的相关问题, 综合理解群落乃至生态系统水平的化学计量学特征以及相关功能。
本文通过对青藏高原47个样地连续3年的调查, 分析了群落水平植物叶片N 、P 含量及N:P 比值的统计特征、分布格局, 并研究了其与气候因子的关系, 揭示了青藏高原草原群落水平上叶片N 、P 的化学计量学特征, 以期为我国植物元素计量学研究以及对其区域性群落尺度上特征的了解提供基础数据。
1 材料和方法
本研究在青藏高原选择了受人类活动影响较少的47个草原群落样地(图1), 2006年至2008年连续3年进行了样品采集。由于交通原因, 2007年和2008年只对其中43个样地进行了重复调查采样。样地主要植被类型为高寒草原与高寒草甸, 其中, 高寒草原建群植物以针茅属(Stipa )植物为主, 高寒草甸建群植物以嵩草属(Kobresia )、薹草属(Carex )植物为
主。样品采集工作在每年的7–8月当地植被生长季
末期进行。为了进行群落水平叶片N 、P 含量的计量学测定, 我们在样地中选取了3个1 m × 1 m 的小样方, 齐地面刈割地上绿色植物, 将小样方中样品各自混匀后, 分别挑选植物叶片约10 g, 并在野外进行简单的烘干处理, 之后, 在实验室65 ℃温度条件下烘干至恒重。
使用粉碎机(NM200, Retsch, Haan, Germany)将叶片粉碎后, 利用C/N 元素分析仪(2400 II CHNS/O Element Analyzer, Perkin-Elmer, Boston, MA, USA)进行N 含量的测定。P 含量的测量采用磷钼蓝比色法。采用H 2O 2-H 2SO 4开氏(Kjedahl)消煮法对叶片粉碎样品进行消解, 利用钼蓝比色法测量并计算P 含量。
根据研究区内65个气象站逐月的平均气温和降水量数据, 通过薄盘光滑样条函数插值方法(Kimeldorf & Wahba, 1971)计算得到每个样地每年的平均气温、降水量, 以及每年生长季(5–8月)的平均气温和降水量。
由于测得的N 、P 数据均呈对数正态分布, 进行统计分析时对数据进行了以10为底的对数转换(Reich & Oleksyn, 2004; Wright et al., 2004)。利用t 检验与单因素方差分析F 检验对比了不同年份间的N 、P 含量, 并对N 、P 与气候因子之间进行Pearson 相关分析。以上统计分析均通过软件SPSS statistics 17.0 (2008, v. 17.0; SPSS Inc., Chicago, USA)完成。
2 结果和分析
2.1 群落水平叶片N 、P 含量及N:P 比值的统计特征
青藏高原草地群落水平的叶片N 、P 含量及N:P 的算术平均值分别为23.5 mg·g –1、1.9 mg·g –1和13.9 (表1); 几何平均值分别为23.2 mg·g –1、1.7 mg·g –1和
杨阔等: 青藏高原草地植物群落冠层叶片氮磷化学计量学分析 19
doi: 10.3773/j.issn.1005-264x.2010.01.004
图1 研究区植被类型及样地分布。
Fig. 1 Vegetation types of the study area and distribution of field sites.
表1 青藏高原草地群落水平叶片氮(N)、磷(P)含量及氮磷比值(N:P)化学计量学特征(2006–2008年)
Table 1 Statistical characteristics of leaf N, P concentrations and N:P ratios at the community level in the grassland of Qing-hai-Tibetan Plateau (2006–2008)
N (mg·g –1) P (mg·g –1) N:P
年 Year n AM/GM SD CV AM/GM SD CV AM/GM SD CV 2006 47 21.5/21.2 3.08 0.14 1.5/1.5 0.50 0.33 15.2/14.6 4.11 0.27 2007 43 26.2/25.7 4.85 0.18 1.5/1.7 0.37 0.21 15.4/15.2 2.46 0.16 2008 43
23.2/22.9
3.24
0.14
2.1/2.1 0.33
0.16
11.0/10.9 1.58
0.14
总计 Total
133 23.5/23.2 3.24 0.18 1.9/1.7
0.48 0.28 13.9/13.5
3.55 0.26
AM , 算术平均数; CV , 变异系数; GM , 几何平均数; SD, 标准偏差。
AM , arithmetic mean; CV , coefficient of variation; GM , geometric mean; SD , standard deviation.
13.5。总体上讲, 叶片P 含量与N:P 的变异系数较近似, 分别为0.28与0.26, 均高于N 含量的变异系数0.18。群落叶片N 、P 含量在同一年内以及总体上均呈显著的正相关关系(图2, R 2 = 0.13–0.45; p < 0.01)。另外, 叶片的N:P 比率与P 含量呈显著的负相关关系, 且相关性较强(图3, R 2 = 0.61; p < 0.01), 而其与N 含量虽有显著相关, 但决定系数较低(R 2 = 0.04; p < 0.05), 说明N:P 比值的变化主要由P 含量变化决定。
2.2 群落水平叶片N 、P 含量及N:P 比值的年际变化
单因素方差分析的结果表明, 2006–2008年的叶片N 、P 含量及N:P 比值均存在显著差异, 由于以往的研究通常只关注植物叶片化学计量学特征的空间变异性, 因此这一现象非常值得我们关注。通过t 检验我们发现, 2006年与2008年的叶片N 含量基本一致, 没有显著差异, 而2007年的叶片N 含量显
著高于其他两年。2007年与2008年的叶片P 含量显著高于2006年, 2008年叶片P 含量与2007年相比
20 植物生态学报Chinese Journal of Plant Ecology 2010, 34 (1): 17–22
https://www.doczj.com/doc/4a11865576.html,
图2 叶片氮含量与磷含量相关关系图。
Fig. 2 Scatter plots showing the relationship between leaf N
and leaf P concentrations.
图3 叶片磷含量与N:P 比值相关关系图。 Fig. 3 Scatter plots showing the relationship between leaf N:P ratios and leaf P concentrations. 表2 叶片氮(N)、磷(P)含量和氮磷比值(N:P)之间及其与气候因子的相关性(Pearson 检验) Table 2 Correlations between leaf N, P concentrations, N:P ratios and climate factors (Pearson test)
P N:P MAT AP GST GSP N 0.446** 0.210* –0.265** 0.205* –0.249** 0.243** P –0.782** –0.420** 0.374** –0.449** 0.300**
N:P
0.274** –0.266** 0.317** –0.159
AP , 年降水量; GSP , 生长季降水量; GST , 生长季平均气温; MAT , 年平均气温。*, p < 0.05; **, p < 0.01。
AP , annual precipitation; GSP , growing season precipitation; GST , growing season temperature; MAT , mean annual temperature. *, p < 0.05; **, p < 0.01.
图4 叶片N:P 比值与年平均气温相关关系图。
Fig. 4 Scatter plots showing the relationship between leaf N:P ratios and mean annual temperature.
平均值略有升高但不显著。N:P 比值同样是2006年与其他两年有显著差异, 2007年与2008年间则差异不显著, 这也体现了N:P 比值主要由P 含量变化决定。
2.3 气候因子对叶片N 、P 含量及N:P 的影响
群落水平上叶片N 、P 含量及N:P 与气候因子显示了较显著的相关性(表2)。其中, 叶片N 含量与年平均气温及生长季平均气温呈显著的负相关关系(R 2 = 0.07, 0.06; p < 0.01), 与年降水量及生长季降水量呈显著的正相关关系(R 2 = 0.04, p < 0.05; R 2 = 0.06, p < 0.01)。叶片P 含量与年平均气温及生长季平均气温呈显著的负相关关系(R 2 = 0.18, 0.20; p < 0.01), 与年降水量及生长季降水量呈显著的正相关关系(R 2 = 0.14, 0.09; p < 0.01)。叶片N:P 比值则与年平均气温及生长季平均气温呈显著的正相关关系
(图4, R 2 = 0.07, 0.10; p < 0.01), 与年降水量呈显著的负相关关系(R 2 = 0.07; p < 0.01), 而与生长季降水量不相关。
3 讨论
3.1 青藏高原草地群落水平多年N 和P 的化学计量学特征
作为在群落水平上进行的多年N 和P 含量研究,
杨阔等: 青藏高原草地植物群落冠层叶片氮磷化学计量学分析 21
doi: 10.3773/j.issn.1005-264x.2010.01.004
我们对比算术平均值, 发现青藏高原N 含量(23.5 mg·g –1)均高于Reich 和Oleksyn (2004)在全球尺度上的研究(20.1 mg·g –1), 而P 含量(1.9 mg·g –1)和N:P 比值(13.9)则与全球水平结果较为一致(分别为1.8 mg·g –1和13.8)。
本研究P 含量与He 等(2008)在西藏地区个体水平的研究相一致, 而N 含量低于He 等(2008)研究中的28.6 mg·g –1, 这也导致了本研究的N:P 比值(13.9)要低于He 等(2008)的研究(15.7)。由于本研究数据是基于群落水平的, 所以, 也许能更真实地反映一个地区植被光合器官的元素化学计量学特征。
通常我们认为, 大尺度上叶片的N 和P 的化学计量学特征是植物对环境条件长期适应的结果。而值得注意的是, 本研究发现群落水平上的N 含量和P 含量及N:P 比值在不同年份间有着显著差异, 这说明了植物群落的N 、P 含量可以被迅速调节以适应环境因子的变动, 而并没有形成较为稳定的元素化学计量关系。这一点或许也可以解释为什么本研究得出的一些结果与He 等(2008)在本地区的研究结果不符。我们推测这种年际变化来自于气候条件、土壤养分的影响和群落物种组成的变化, 并在接下来分析了气候因子与群落化学计量学特征的相关关系。同时这也提出了一系列的问题, 即: 仅做一次测量是否可以明确真实地反映一个地区植被的元素计量学特征?而当这些因素在逐年变化的时候, 如何准确地描述一个地区或一种植被群落水平的N 和P 的化学计量学特征?这些都是需要我们做进一步研究与分析的。
3.2 青藏高原草地N 、P 含量及N:P 比值与气候因子的相关性
虽然有研究表明, 叶片的N 、P 含量和N:P 比值反映了植物对当地土壤条件的适应(Thompson et al., 1997; Güsewell, 2004), 但也有研究发现, 温度也是植物叶片N 、P 含量的影响因子之一。例如, 由海拔引起的温度变化可以解释植物叶片N 含量的种内差异(K?rner, 1989)。有假说认为: 土壤P 在土壤发育时间较长的、温暖地区被高度淋溶的土壤中是植物生长更主要的限制因子; 而在土壤发育时间较短的、中高纬度的土壤中, N 则是主要的限制因子(the soil substrate age N/P hypothesis) (Reich & Oleksyn, 2004)。本研究结果部分支持该假说, 即叶片N:P 比值与温度呈显著的正相关关系, 在较温暖地区生长
的植物相对更加地受到P 的限制。而研究结果中的叶片N:P 比值与降水量呈显著负相关关系(研究区域内降水量与气温未表现出相关性), 似乎与该假说不符。我们认为这一结果与本研究集中于高海拔较寒冷区域, 土壤发育、淋溶与降水量关系不明确有关。
Reich 和Oleksyn (2004)研究了大尺度上叶片的N 、P 含量和N:P 比值与气候的相关性, 将植物在较低温度下的高N 、P 含量解释为植物为抵消低温对代谢反应抑制的一种适应机制。我们在研究中发现, 年平均气温及生长季平均气温均与青藏高原草地区群落水平上叶片N 含量呈显著的负相关关系, 与叶片P 含量有着相对较强的显著负相关关系, 而N:P 比值则随着温度的升高而增加, 这一结果与Reich 和Oleksyn (2004)在全球尺度上的研究结果完全一致。说明在群落尺度上, 植物叶片的化学计量学特征有着与个体水平相类似的与环境因子的相关性。
另外, 对比各气候因子与N 含量和P 含量的相关性, 我们发现, P 含量与环境因子的相关关系显著性水平更高、决定系数更大。这可能是由于P 含量的变异系数高于N 含量的变异系数造成的。而根据生长速率假说, 自养生物加大生物量的积累速率需要提高其P 含量、降低其N:P 比值来加速蛋白质的合成, 加强同化作用(Elser et al., 2003; Niklas et al., 2005)。我们推测, P 含量较高的变异系数及其与环境因子表现出的这种相关性, 在一定程度上验证了生长速率假说, 体现了植物群落对当地气候条件的一种适应。
4 结语
本研究首次进行了植物叶片N 、P 含量化学计量学的多年群落水平的调查分析, 结果显示: 青藏高原草地群落水平叶片的N 、P 含量及N:P 比值分别为23.2 mg·g –1、1.7 mg·g –1和13.5; 叶片N 、P 含量及N:P 比值均存在着显著的年际差异; 温度因子与N 、P 含量及N:P 比值的相关关系与全球尺度的研究一致。而植物叶片N 、P 含量的年际变动是否存在着某种规律, 或是某种一致性、周期性?这是需要我们进一步研究和思考的问题。
致谢 国家自然科学基金(30870381和30670322)资
助项目。项目野外考察中得到了内蒙古大学梁存柱老师、北京大学周文嘉、石岳等同学的支持, 在此
第32卷第15期2012年8月 生态学报ACTA ECOLOGICA SINICA Vol.32,No.15Aug.,2012 基金项目:国家重点基础研究发展计划项目(2009CB421104);中国科学院战略性先导科技专项(XDA05060100) 收稿日期:2011-04-20;修订日期:2012-04-24 *通讯作者Corresponding author.E-mail :ghliu@rcees.ac.cn DOI :10.5846/stxb201104200523丁小慧,宫立,王东波伍星,刘国华.放牧对呼伦贝尔草地植物和土壤生态化学计量学特征的影响.生态学报, 2012,32(15):4722-4730.Ding X H ,Gong L ,Wang D B ,Wu X ,Liu G H.Grazing effects on eco-stoichiometry of plant and soil in Hulunbeir ,Inner Mogolia.Acta Ecologica Sinica ,2012,32(15):4722-4730. 放牧对呼伦贝尔草地植物和土壤生态 化学计量学特征的影响 丁小慧1,2,宫立1,2,王东波3,伍星1,刘国华 1,*(1.中国科学生态环境研究中心城市与区域生态国家重点实验室,北京100085;中国科学院研究生院,北京 100049;3.内蒙古自治区呼伦贝尔市环境科学研究所,呼伦贝尔 021008)摘要:放牧通过畜体采食、践踏和排泄物归还影响草地群落组成、植物形态和土壤养分,植物通过改变养分利用策略适应环境变化。通过分析呼伦贝尔草原放牧和围封样地中的群落植物和土壤的碳氮磷养分及化学计量比,探讨放牧对生态系统化学计量 学特征和养分循环速率的影响机制。结果如下:(1)群落尺度上,放牧和围封草地植物叶片C 、 N 和P 的含量没有显著差异;但是在种群尺度上,放牧草地植物叶片N 含量显著高于围封草地;(2)放牧草地土壤全C 、全N 、有机C 、速效P 含量,低于围封草地,硝态N 含量高于围封草地;土壤全P 和铵态N 指标没有显著差异;(3)放牧草地植物C ?N 比显著低于围封草地,植物残体分解速率较快,提高了生态系统养分循环速率。 关键词:草地生态系统;放牧;围封;化学计量学;呼伦贝尔 Grazing effects on eco-stoichiometry of plant and soil in Hulunbeir ,Inner Mogolia DING Xiaohui 1,2,GONG Li 1,2,WANG Dongbo 3,WU Xing 1,LIU Guohua 1, *1State Key Laboratory of Urban and Regional Ecology ,Research Center for Eco-Environmental Sciences ,Chinese Academey of Sciences ,Beijing 100085,China 2Graduate University of Chinese Academy of Sciences ,Beijing 100049,China 3Institute of Environmental Science of Inner Mongolia ,Hulunbeir 021008,China Abstract :The grassland in Inner Mongolia ,one of the largest remaining grassland ecosystems in the world ,is extensive and covers an area of up to 68%of the entire region.Recently ,with the increasing needs of livestock products ,the ecological function of the grassland is severely degraded by overgrazing.The effect of grazing has become an urgent and crucial issue in the study of grassland ecology.The early researches mainly focused on the effects of grazing on the productivity ,community structure and biodiversity of the grasslands through the browsing ,trampling around and excretion of livestock.Focusing on C4soil or plants ,only a few studies have considered the effects of grazing on the stoichiometry traits of soil and plants.The mechanism by which grazing affects the stoichiometry traits of soil and plants is far from clear.For this issue , we carried out a study considering the role of grazing on the stoichiometry traits of soil and plant at both population and community scales.We determined the C ,N ,P content and the C /N ratios of plant and soil to explore how grazing affects the stoichiometry traits and nutrient cycling rates in an ecosystem.We found that :(1)On community scale ,C ,N ,P content of the leaves had no-significant difference between the grazed and the exclosure grassland ;however ,on species scale ,the N content in
植物生态学报 2010, 34 (1): 2–6 doi: 10.3773/j.issn.1005-264x.2010.01.002 Chinese Journal of Plant Ecology http:// www. https://www.doczj.com/doc/4a11865576.html, —————————————————— 收稿日期Received: 2009-11-24 接受日期Accepted: 2009-12-15 生态化学计量学: 探索从个体到生态系统的统一化理论 贺金生1,2 韩兴国3 1 北京大学城市与环境学院生态学系, 北京 100871; 2中国科学院西北高原生物研究所, 西宁 810008; 3 中国科学院植物研究所, 北京 100093 Ecological stoichiometry: Searching for unifying principles from individuals to ecosystems HE Jin-Sheng 1,2 and HAN Xing-Guo 3 1 Department of Ecology, College of Urban and Environmental Sciences, Peking University, Beijing 100871, China; 2Northwest Institute of Plateau Biology, Chinese Academy of Sciences, Xining 810008, China; and 3Institute of Botany, Chinese Academy of Sciences, Beijing 100093, China 从1909年丹麦哥本哈根大学Johannes Warming (1841–1924)出版第一本生态学教科书《植物生态学》到现在, 生态学经历了100年的发展。作为研究生物与生物、生物与环境相互关系的学科, 生态学具有高度的综合性和交叉性, 研究的问题也具有复杂性和多尺度的特点。正因为这样的学科特点, 传统上生态学家多强调研究对象的特殊性、研究问题的局域性, 缺乏统一的理论框架。进入21世纪以后, 生态学在统一化理论方面出现了一些尝试, 代表性的有生态学的代谢理论(metabolic theory of ecology) (Brown et al ., 2004)和生态化学计量学(ecological stoichiometry) (Sterner & Elser, 2002)。 生态化学计量学综合生物学、化学和物理学的基本原理, 利用生态过程中多重化学元素的平衡关系, 为研究C 、N 、P 等元素在生态系统过程中的耦合关系提供了一种综合方法。1958年, 哈佛大学的A. Redfield 首次证明: (1)海洋浮游生物的C 、N 、P 有特定的组成(摩尔比106:16:1, 该比率后被称为Redfield 比率); (2)这一比率受海洋环境和生物相互作用的调节(Redfield, 1958)。这一开创性的研究成为以后生态化学计量学的奠基之作。从Elser 和Hassett (1994)以及Elser 等(2000)的重要文章, 到以后Sterner 和Elser (2002)的专著《Ecological Stoichi- ometry 》的出版, 标志着生态化学计量学理论的系统化和逐步成熟。 2004年著名生态学杂志《Ecology 》 (85卷1 177页)和2005年《Oikos 》(109卷3页)分别出版了“Ecological stoichiometry”专辑, 进一步推动了这一领域的发展。本刊也于2005年发表了曾德慧和 陈广生(2005)的综述性文章, 系统介绍了生态化学计量学的理论与进展。 1 生态化学计量学为从个体到生态系统统一化理论的构建提供了新思路 生态化学计量学主要研究生态过程中化学元素的比例关系, 因此跨越了个体、种群、群落、生 态系统、景观和区域各个层次。生态化学计量学目前主要集中在C 、N 、P 元素的计量关系。这是因为, C 、N 、P 是重要的生命元素, 它们是地球上所有生命化学组成的基础。一般来说, 组成地球上有机体蛋白质的16%是N, 核酸组成的9.5%是P, 这两个比例在不同来源的生物中相对稳定。而有机体干物质的50%左右是C, 这一比例在生物的不同类群中随细胞的结构组成发生变化。生物在长期进化过程中, 形成了一定的内稳态机制(homeostatic mecha-nism), 即生物在变化的环境(包括食物)中具有保持其自身化学组成相对恒定的能力, 它是生态化学计量学存在的前提(Sterner & Elser, 2002)。现在还不清楚生物组成的其他元素是否存在相对稳定的计量关系, 但从生态化学计量学角度, 对其他化学元素的探索是必要的, 特别是S 的作用。 无论是植物个体水平, 还是生态系统水平, C 、N 、P 都是相互作用的。研究其中一个元素在生态学过程中的作用, 必须同时考虑其他元素的影响。生态化学计量学为研究C 、N 、P 等主要元素的生物地球化学循环和生态学过程提供了一种新思路 (Sterner & Elser, 2002; Güsewell, 2004)。在植物的个
自然资源学报JOURNAL OF NATURAL RESOURCES 第30卷第10期 2015年10月V ol.30No.10Oct.,2015延河流域不同立地条件下植物-枯落物-土壤 生态化学计量学特征 向云1,4,程曼1,安韶山1,2*,曾全超2,3 (1.西北农林科技大学资源环境学院,陕西杨凌712100;2.中国科学院、教育部水土保持与生态环境研究 中心黄土高原土壤侵蚀与旱地农业国家重点实验室,陕西杨凌712100;3.中国科学院大学, 北京100049;4.山西省环境科学研究院,太原030000) 摘要:通过对黄土高原延河流域不同立地条件的植物、枯落物和土壤进行调查、取样和分析,选取不同立地条件下的共有植物达乌里胡枝子,研究了不同立地条件下共有植物-枯落物-土 壤的碳、氮、磷化学计量特征,以期探索不同立地条件中的元素迁移和转化。结果表明:1)土壤 碳氮比的平均值为10.88,土壤碳磷比和氮磷比的平均值分别为23.14和2.13。2)土壤碳氮比表 现为阳坡>阴坡、沟坡>峁坡,氮磷比无显著差异;共有植物碳氮比表现为阳坡>阴坡,峁坡>沟 坡,碳磷比和氮磷比为阴坡>阳坡,峁坡>沟坡;枯落物碳氮比表现为阳坡>阴坡,峁坡>沟坡;氮 磷比和碳磷比在不同立地条件中均表现为阴坡>阳坡,沟坡>峁坡。3)通过对不同立地条件下 植物-枯落物-土壤化学计量特征的探讨,得知沟坡条件下土壤有机碳累积速率相对高于峁坡,阳坡土壤有机碳固定大于阴坡,各立地条件中土壤全磷含量相对一致;在植物生长后期以及枯 落物分解过程中,阳坡和峁坡表现为氮素迁移转化相对强烈,阴坡和沟坡则有利于磷的迁移和 转化。 关键词:土壤;植物;枯落物;生态化学计量学;立地条件 中图分类号:S154 文献标志码:A 文章编号:1000-3037(2015)10-1642-11DOI :10.11849/zrzyxb.2015.10.004 植物生长发育所需的养分主要来源于土壤,土壤对植物的生长具有非常重要的影响[1]。同时,植物在生长过程中通过光合作用固定碳,以枯落物分解和根系分泌的方式将养分归还于土壤中。枯落物及其分解过程是联系植被和土壤的纽带,植物-枯落物-土壤构成一个微观的生态系统,其中土壤养分供应量、植物养分需求量以及枯落物分解过程中养分的返还量的不断调节[2],使植物-枯落物-土壤系统的养分含量具有明显的时空变化,但同时也增加了植物-枯落物-土壤中碳、氮、磷之间相关关系的复杂性。 生态化学计量学(Ecological Stoichiometry )结合了生态学和化学计量学的基本原理,是研究生物系统能量平衡和多重化学元素平衡的科学,它是分析多重化学元素的质量平衡对生态过程相互影响的一种理论[3-4],为探究不同生态系统中元素迁移和元素限制提供了有效手段。近年来,越来越多的研究工作者利用化学计量手段来研究土壤植物的元素 收稿日期:2014-09-10;修订日期:2015-04-23。 基金项目:国家自然科学基金面上项目(41171226);新世纪优秀人才支持计划(NCET-12-0479);国家自然科学基金重点基金(41030532)。 第一作者简介:向云(1987-),男,湖南澧县人,博士研究生,主要从事植被恢复与土壤的相互作用机制研究。E-mail:xy020824@https://www.doczj.com/doc/4a11865576.html, *通信作者简介:安韶山(1972-),男,研究员,主要从事土壤生态学、水土保持效益评价、流域生态与管理研究。E-mail:shan@https://www.doczj.com/doc/4a11865576.html,
高三化学复习专题一物质的量 【考纲要求】 了解定量研究的方法是化学发展为一门科学的重要标志,掌握化学常用计量方法。 1.了解物质的量的含义,并能进行简单的化学计算。 2.了解物质的量的单位——摩尔、摩尔质量、气体摩尔体积、物质的量浓度、阿伏加德罗常数的含义。 3.根据物质的量与微粒数目、气体体积(标准状况下之间的相互关系进行有关计算。 4.了解溶液的组成。理解溶液中溶质的质量分数的概念,并能进行有关计算。【过程方法】要注意气体摩尔体积的适用条件、要注意物质的状态、要注意物质的组成形式、要注意晶体的结构、要注意微粒的种类、要注意氧化还原反应中电子转移数目与元素价态的变化、要注意可逆过程或化学平衡等等。【重点知识整合】【考点1】关于以物质的量为中心的各化学量的相互关系
特别提醒: 1.以物质的量为中心的计算需注意的问题(1 “一个中心”:以物质的量为中心。 (2 “两个前提”:在应用V m =22.4 L·mol -1时,一定要有“标准状况”和“气体及其状态”两个前提条件(混合气体也适用 。(3 “三个关系”: ①直接构成物质的粒子与间接构成物质的粒子(原子、电子等间的关系; ②摩尔质量与相对分子质量间的关系;
③“强、弱、非”电解质与溶质粒子(分子或离子数之间的关系。 (4“四个无关”:物质的量、质量、粒子数的多少均与温度、压强的高低无关;物质的量浓度的大小与所取该溶液的体积多少无关(但溶质粒子数的多少与溶液体积有关。 1、(上海卷工业上将氨气和空气的混合气体通过铂-铑合金网发生氨氧化反应,若有标准状况下VL 氨气完全反应,并转移n 个电子,则阿伏加德罗常数N A 可表示为【】 A . 5V 11.2n B .11.2n 5V C .5n 22.4V D .5V 22.4n 班级:___________ 姓名:___________ 学号:___________ 2、(广东卷设N A 为阿伏加德罗常数的数值,下列说法正确的是【】 A .常温下,4g CH 4含N A 个C —H 共价键 B .常温常压下,22.4L 的NO 2和CO 2混合气体含有2N A 个O 原子 C .1 mol Fe 与足量稀HNO 3反应,转移个2N A 个电子 D .1L 0.1 mol/L NaHCO 3溶液中含有0.1N A 个HCO 3—
化学常用剂量 1.高锰酸钾法测定水体COD (化学需氧量) 的实验步骤如下: 步骤1 准确移取100 mL 水样,置于250 mL 锥形瓶中。加入10 mL 1∶3 的硫酸,再加入15.00 mL 0.020 0 mol ·L -1 KMnO 4 溶液(此时溶液仍呈紫红色)。 步骤2 用小火煮沸10 min (水中还原性物质被MnO -4氧化,本身还原为Mn 2+ ),取下锥形瓶趁热加10.00 mL 0.050 0 mol ·L -1 Na 2C 2O 4溶液,充分振荡(此 时溶液为无色)。 步骤3 趁热用0.020 0 mol ·L -1 KMnO 4溶液滴定至呈微红色,消耗KMnO 4 溶液4.500 mL 。 通过计算确定该水样的化学需氧量(写出计算过程)。 [已知: COD 是指在一定条件下,以氧化1L 水样中还原性物质所消耗的氧化剂的量,通常换算为需要的O 2的质量(mg),COD 的单位mg ·L -1。] 【解析】n (Na 2C 2O 4)=0.050 0 mol ·L -1×10.00 mL ×10-3 L ·mL -1=5.000×10-4 mol ,两次共消耗n (KMnO 4)=0.020 0 mol ·L -1×(15.00+4.500)mL ×10 -3 L ·mL -1= 3.900×10-4mol ,氧化有机物消耗n (KMnO 4)=3.900×10-4mol - 2 5 n (Na 2C 2O 4)=3.900×10-4 mol -25×5.000×10-4mol =1.900×10-4 mol ,n (O 2)= 54×1.900×10-4mol = 2.375×10-4mol ,m (O 2)=2.375×10-4mol ×32 g ·mol -1=7.600×10-3g = 7.600 mg ,COD =76.0 mg ·L -1。 2.为确定由CoC 2O 4·2H 2O 获得Co 3O 4的最佳煅烧温度,准确称取4.575 g 的CoC 2O 4·2H 2O 样品,在空气中加热,固体样品的剩余质量随温度的变化如图所示(已
化学计量内容丰富,围绕化学计量的核心,即保证化学测量结果的准确、可靠和可比,归纳起来至少应包括以下几个方面: 一、SI基本单位摩尔的复现和基本物理量的测量 摩尔是SI单位制中表示物质的量的基本单位,也是唯一一个直接关于化学计量的SI单位,是化学计量的基础。物质的量(即化学成分量)测量是化学测量的最主要内容。将化学成分量溯源至SI单位,是化学计量追求的终极目标。摩尔的复现和基本物理常数的测量是国际化学计量界共同努力的方向。 二、化学计量基(标)准的建立、复现、保存和使用 化学测量从内容上说,可分为化学成分量、物理化学量、化学工程量和生物量测量几大类。目前,在我国已经建立的化学计量基准只有6项,远不能满足化学测量溯源的需求。由于化学测量的复杂性,要研究和建立更多的化学计量基准,其难度是巨大的。即使是已经建立的计量基准,随着科学技术的发展,也需要不断地更新,才能持续满足社会发展的需要。此外,种类数以百计的化学测量标准,作为全国、部门或区域的量值传递的测量标准,在量值溯源链中发挥了非常重要的作用。它们的建立、复现、保存、更新和使用在一定程度上,保证了化学测量结果的准确可靠。 三、标准物质及其相关技术的研究和应用 标准物质是测量标准的一个种类。其与传统定义的物理测量标准相比,具有其特殊性,因此国际上有专门针对标准物质的定义。化学测量绝大多数是相对测量,而且影响测量不确定度的因素复杂。可以说,没有标准物质,化学测量无法实现,更谈不上准确可靠和可比。标准物质可分为纯物质和基体标准物质两大类。其中纯物质及其相应的纯溶液,主要用于校准分析仪器,或用作测量标准给待测量赋值;另一类是基体标准物质,主要用于考察确认测量方法、测量人员以及测量过程的质量控制,有时也用于校准仪器(X-射线荧光光谱法、近红外光谱法)。由于化学测量涉及的物质种类繁多,成分复杂,不同基体、不同物质、甚至是同一种物质和成分的不同含量水平,对测量结果的影响也不尽相同,某一化学特定量的标准物质基本上仅适用于基体和含量与其相同或相似的特定量的测量。目前,虽然已经有上千种标准物质在各个领域使用,但是仍然有许多重要的领域需要更多适用的标准物质。尤其是制造工业、贸易、健康和环境等领域,标准物质的需求量巨大。需要不断地研究标准物质的有关理论、测量技术、不确定度表述方法等,研制不同种类的标准物质满足量值传递和溯源的要求。这些工作需要化学计量工作者和相关测量领域分析专家长期共同的努力。 四、化学测量方法的研究、确认和应用
(1)生态化学计量学涵义及理论基础 进入21世纪以来,生态学已经发展为研究从分子尺度到全球尺度的多学科相互交叉的综合性科学,然而如何使相关学科不同层次(分子、细胞、有机体、种群、群落、生态系统和全球尺度)的研究理论能够有机统一起来,一直是该领域探讨的热门话题。为解决这个问题,生态化学计量学(Ecological stoichiometry)在最近20年应蕴而生(Elser,J.,2000;Sterner,R.2002;曾德慧,2005)。 所谓生态化学计量学结合了生物学、化学和物理学等基本原理,包括了生态学和计量学的原理,考虑了热力学第一定律、生物进化的自然选择原理以及分子生物学中心法则的理论,研究在生态过程和生态作用中能量与多重化学元素(主要是C、N和P素)平衡以及这种平衡对生态系统的影响的科学(Zhang,L.2003;程滨,2010;王绍强,2008)。简言之,生态化学计量学就是研究生态系统中能量与化学元素间平衡的学科[191],主要强调的是活有机体的主要组成元素(特别是C、N和P)的关系。 用组成有机体的元素的独特化学计量值来反映生物进化的结果,主要包括影响生物生长速率、健康状况、新陈代谢、组织结构和生态演替等某些方面(Michaels,A.,2003)[199]。在生态学中聚焦元素间化学计量比,主要意义在于:首先,元素容易在不同生物组织水平间移动,因此能够计算元素的组成和估计进入或流出各种生物实体化学元素的通量;其次,元素是不变的,可根据质量守恒定律去研究元素在各生物学组织水平间的流量;最后,生命有机体都是由化学反应所产生、维持和增殖的化学实体,且化学反应都是以复杂和耦合的反应网形式进行的(Sterner,R.2002;曾德慧,2005)。 生态化学计量学发展了两个重要的具体理论,即动态平衡原理和生长速率理论。 动态平衡原理是生态化学计量学理论成立的理论基础(曾德慧,2005)。指有机体各种元素组成与其周围环境(包括利用资源)养分元素供应保持相对稳定的一种状态(Sterner,R.2002)。它被认为是生态化学计量学理论存在的理论基础和生命的本质特征。 “生长速率理论”是有机体生态化学计量控制的基本途径,这个理论提供了生态化学计量控制生命进化论、细胞生物学特性、种群动态以及生态系统功能机制的基本框架(Elser,J.,2000;曾德慧,2005;Reiners,W.1986;Vanni,M.2002)。最初源于对最适条件下不同物种生物量C∶N∶P和最大生长速率的研究(Elser,J.1996;Main,T.1997),而且适用于同物种生物的生物量C∶N和N∶P变化的研究(Hessen,D.,2004;Hessen,D.;1990)。生长速率理论把生物组织的不同层次(分子、个体、种群和生态系统)有机地联系起来了。并认为,单个有机体P的生物地球化学过程与生长及繁殖过程紧密相关(它是种群动态的一个主要决定者)(Elser,J.;1996)。具体来说,有机体C∶N∶P的变化主要由生物体P含量的变化所决定。生物有机体必须调整它们的C∶N∶P化学计量比来适应生长速率的改变,生长快速的物种通常具有较低的C∶P和N∶P(Elser,J.,2003;Makino,W.,2003),这种直接或间接地造成生活史特性产生变化的演化过程会对有机体的元素组成产生影响(Elser,J.,2000;曾德慧,2005)。 (2)陆地生态化学计量学研究的基本内容 生态化学计量学目前多数研究集中水生生态系统,其理论的提出、验证和补充大都是基于对海洋生物的研究获得的,陆地生态系统方面的研究起步较晚,但近10 年来也得到了较大的发展。其中对天然草原和森林亚系统研究较多。总体分为几个方面: 首先,植物个体生态化学计量学研究 通过研究植物体自身生态化学计量特征与生境生态化学计量特征之间的关系与差异;找
生态学:研究生物的生理特性和生活习性及生存、生长、生活、生育与生衍状态的科学。 生态系统:在一定地理空间范围内的生物群落与非生物环境构成的具有特定组成、结构和功能属性的生态学系统;可以抽象的概括 为是一个生物组分及其与环境之间相互作用、相互依存、动态 变化的物理、化学和生物学复合系统。 生态系统生态学:是研究生态系统的科学,是认识生态系统的科学知 识体系。 生物学物种或生物种:在自然界中同一个物种的个体之间应该繁殖出 具有生育能力的后代。 自然选择的准则:在特定环境中能够留下更多的后代,即保留适合度 高的个体或类型。 物理防御:种植物刺、种植物壳、植物的种子(脱落) 化学防御:植物和动物生成次生化合物保护剂(有毒化合物,消化不良化合物。 形态和行为防御:体色或形态图案成为隐态,警戒态,模仿味道不佳猎 物体姿的贝氏拟态、警戒色+模拟体姿的繆氏拟态。生存环境:影响活体生物功能的非生物环境,包括温度、相对湿度、氧气、pH、盐度、污染物浓度等、条件可以改变但不能被消耗。生境:生物有机体的生活场所。 生态位:生物有机体对生境条件以及生境资源需求的综合,有机体的生态位是生物的生活模式。
基础生态位:在竞争者或扑食者不存在情况下,一个物种可以生存繁殖的潜能。 实际生态位:在竞争者或扑食者存在情况下可以生存繁殖的条件和资源范围。 低温的影响与适应策略:冷害与冻害(细胞的渗透压和水平衡、细胞 膜);避冻策略和耐冻策略。 高温的影响与适应策略: 高温危害:酶失活和变性、高温失水(蒸腾维温)、火灾。 适应策略:蒸腾维温--动植物的水冷系统,特化物种、耐高温生物,嗜热真菌,极端嗜热原核生物。 必需资源:资源间不可替代。 互补性资源:摄取两种资源湿比摄取一种资源的需求和消耗的更少。拮抗性资源:摄取两种资源明显比摄取一种资源的需求和消耗的更多。抑制性资源:必须资源在适宜范围年供给量增加促进生长,在过量供给时则变得得有害,抑制生物生长率。 氮限制机制:供给光合作用酶的N素不足,叶绿体内的ATP、NADPH 和羧化作用受限制。 磷限制机制:供给光合成RUBP的磷酸盐或者磷糖酸不足,限制光合速率。 生理生态学调控机制:①植物气孔对水碳耦合的调控机理②根系对 水分、养分吸收和碳归还的调控机理③微生物功 能群对碳氮转化和排放的调控机理④土壤-植物
常见化学计算方法 主要有:差量法、十字交叉法、平均法、守恒法、极值法、关系式法、方程式叠加法、等量代换法、摩尔电子质量法、讨论法、图象法(略)、对称法(略)。 一、差量法 在一定量溶剂的饱和溶液中,由于温度改变(升高或降低),使溶质的溶解度发生变化,从而造成溶质(或饱和溶液)质量的差量;每个物质均有固定的化学组成,任意两个物质的物理量之间均存在差量;同样,在一个封闭体系中进行的化学反应,尽管反应前后质量守恒,但物质的量、固液气各态物质质量、气体体积等会发生变化,形成差量。差量法就是根据这些差量值,列出比例式来求解的一种化学计算方法。该方法运用的数学知识为等比定律及其衍生式: a b c d a c b d == --或c a d b --。差量法是简化化学计算的一种主要手段,在中学阶段运用相当普遍。常见的类型有:溶解度差、组成差、质量差、体积差、物质的量差等。在运用时要注意物质的状态相相同,差量物质的物 理量单位要一致。 1.将碳酸钠和碳酸氢钠的混合物21.0g ,加热至质量不再变化时,称得固体质量为1 2.5g 。求混合物中碳酸钠的质量分数。 2.实验室用冷却结晶法提纯KNO 3,先在100℃时将KNO 3配成饱和溶液,再冷却到30℃,析出KNO 3。现欲制备500g 较纯的KNO 3,问在100℃时应将多少克KNO 3溶解于多少克水中。(KNO 3的溶解度100℃时为246g ,30℃时为46g ) 3.某金属元素R 的氧化物相对分子质量为m ,相同价态氯化物的相对分子质量为n ,则金属元素R 的化合价为多少? 4.将镁、铝、铁分别投入质量相等、足量的稀硫酸中,反应结束后所得各溶液的质量相等,则投入的镁、铝、铁三种金属的质量大小关系为( ) (A )Al >Mg >Fe (B )Fe >Mg >Al (C )Mg >Al >Fe (D )Mg=Fe=Al 5.取Na 2CO 3和NaHCO 3混和物9.5g ,先加水配成稀溶液,然后向该溶液中加9.6g 碱石灰(成分是CaO 和NaOH ),充分反应后,使Ca 2+、HCO 3-、CO 32-都转化为CaCO 3沉淀。再将反应容器内水分蒸干,可得20g 白色固体。试求: (1)原混和物中Na 2CO 3和NaHCO 3的质量; (2)碱石灰中CaO 和NaOH 的质量。 6.将12.8g 由CuSO 4和Fe 组成的固体,加入足量的水中,充分反应后,滤出不溶物,干燥后称量得5.2g 。试求原混和物中CuSO 4和Fe 的质量。 二、十字交叉法 凡能列出一个二元一次方程组来求解的命题,即二组分的平均值,均可用十字交叉法,此法把乘除运算转化为加减运算,给计算带来很大的方便。 十字交叉法的表达式推导如下:设A 、B 表示十字交叉的两个分量,AB —— 表示两个分量合成的平均量,x A 、x B 分别表示A 和B 占平均量的百分数,且x A +x B =1,则有:
化学计量在中国的进展 1生态化学计量学概念 生态化学计量学(ecological stoichiometry)结合了生物学、化学和物理学等基本原理,包括了生态学和化学计量学的基本原理,考虑了热力学第一定律、生物进化的自然选择原理和分子生物学中心法则的理论,是研究生物系统能量平衡和多重化学元素(主要是碳、氮、磷)平衡的科学,以及元素平衡对生态交互作用影响的一种理论,这一研究领域使得生物学科不同层次(分子、细胞、有机体、种群、生态系统和全球尺度)的研究理论能够有机地统一起来。 2生态化学计量学小史 1862年李比希提出的最小因子定律(Liebig’Slaw of the minimum),这个理论认为低于某种生物需要的最少量的任何特定因子,是决定该种生物生存和分布的根本因素。这个定律阐述的精华就是生物体中元素的组成平衡对于生物体生长是非常重要的。李比希认为化学在动物和植物生理学研究中具有不可替代的地位,许多生命有机体对于贫瘠环境的适应的研究以此为基础产开,大量证据表明,限制性元素的含量不同会影响有机体元素的组成。 1925年,Lotka首先将物理一化学系统热力学定律与生物世界相联系起来,著成了《物理生物学的基础)(Elements of Physical Biology)一书,提出了一个重要的模型:捕食者~猎物相互作用模型,这个模型定量阐述了生物之间的相互作用关系。许多生态学基础理论由于他的思想的影响得到了广泛的完善和发展。1958年,哈佛大学的Redfiel首次提出了Redfield比率:海洋浮游生物的C、N、P有特定的组成,摩尔比为106:16:1,后人认为这个比率不是不变的,而是受海洋环境和生物相互作用的调节。这个假设的提出极大发展了海洋生物地球化学研究。 1986年Reiners集合前人的研究结果,提出了化学计量学理论在生态学中的应用,并且结合化学计量学理论提出了生态学研究的理论模型。自Reiners提出生态化学计量学,拉开了化学计量学在生态学中应用的序幕,科学家们在2O多年的时间内取得了瞩目的成绩。研究结果显示,不仅群落结构与动态、物种共生、营养级动态、生物的养分限制受生态化学计量学的影响,生态系统养分循环与供求平衡和全球生物地球化学循环等关系也受生态化学计量学的制约。因此,生态化学计量学成了探索从个体到生态系统的统一化的一个重要理论成为连接分子、细胞、种群、群落和生态系统等不同尺度生物学研究的新工具,为研究营养级动态、生物多样性和生物地球化学循环提供了崭新的视点E 。 生态化学计量学近年来在国内发展较快。最近的研究不仅包括了不同生态系统类型之间不同演替阶段植物之间生态化学计量特征的差异,还包括了植物叶片生态化学计量学特征的季节变化,以及植物叶片和细根不同器官之间计量特征的关联_3]。 3 生态化学计量学在我国的研究进展 3.1不同生态系统类型之间 自从生态化学计量学被作为生态系统研究的一个重要补充理论,已经在需多个方面得到了应用,比如种群动态、森林演替和碳循环。尽管我国在这方面的研究起步较国外玩,但是也在东部南北样带、草地生态系统及全国水平的陆地生态系统做了大尺度的研究。2004年,McGroddy等总结了世界范围内森林生态系统的叶片和凋落物的生态化学计量学特征,发现了不同生物群(温带阔叶林、温带针叶林和热带森林)具有不同的生态化学计量学特征,但有关同一区域不同森林类型间的生态化学计量学研究还未曾报道,于是吴统贵]以珠江三角洲3种典型森林类型(常绿阔叶林、针阔混交林和针叶林)为研究对象,分析了各类型优势乔木叶片C、N、P化学计量特征。闫恩荣]以浙江天童常绿阔叶林、常绿针叶林和落叶阔叶林为对象,通过对叶片和凋落物c:N:P比率与N、P重吸收的研究,揭示3种植被类型N、P养分限制和N、P重吸收的内在联系。 3.2不同演替阶段之间
-46-科学技术创新2019.11 湿地生态系统C、N、P生态化学计量学特征的研究进展 范全城柴娜李萍王志强 (青岛大学环境科学与工程学院,山东青岛266071) 摘要:湿地(wetland)是处于水生生态系统和陆生生态系统之间的生态交错区,兼具水陆生态系统的特征,蕴含了丰富的自然资源,是地球上生产力最高的过渡生态系统之一,其与森林生态系统,海洋生态系统被称为地球三大生态系统。 关键词:湿地;生态系统;化学计量学 中图分类号:X171文献标识码:A文章编号:2096-4390(2019)11-0046-02 湿地(wetland)是处于水生生态系统和陆生生态系统之间的生态交错区,兼具水陆生态系统的特征,蕴含了丰富的自然资源,是地球上生产力最高的过渡生态系统之一,其与森林生态系统,海洋生态系统被称为地球三大生态系统。由于湿地生态系统的复杂性与多样性,对于湿地的定义还没有一个完全科学统一的定义,而纵观国内外对湿地的定义也多达60种。而有关湿地的最早的定义可以追溯至20世纪50年代,美国渔业局首次对湿地进行了定义,主要包含了水文和植物两大板块。目前,被大多数国家所接受的是《国际生物学计划》和《湿地公约》中所提及的定义,其中前者定义湿地为陆地与水域之间的过渡区域或生态交错带,对水域的界定是在低水位时水深不得大于2m;后者将其定义为低潮时水深在6m以下的水域或海洋水域,还包含湿地内的岛屿及临近湿地的近海岸地区,如河流、湖泊、沼泽、滩涂、水库、浅海区等。我国湿地管理部门在《湿地公约》对湿地定义的基础上,规定湿地是指天然或人工的、长久性或暂时性沼泽地、泥炭地或者水域地带。带有静止或流动淡水、半咸水、咸水水体等,包括低潮时水深不高于6米的海域。 1生态化学计量学概述 近年来,生态化学计量学发展迅速,在水生生态系统和陆生生态系统地研究取得了重大的突破,研究领域广泛涉及到植物组织、动物、微生物、土壤和枯落物元素的生态化学计量学,涵盖了物种水平上物种之间的生物关系,群落水平上群落结构变化与养分的动态平衡,全球水平上生态过程与生物地球化学循环过程。与国外的研究相比,国内的生态化学计量学起步比较晚,研究的基础较为薄弱,但是近30年来发展迅速。其中比较具有代表性和影响力的研究有:张仲胜等较为全面的概述了生态化学计量学的起源、发展历程和发展前景,统计分析了国内119块湿地土壤的生态化学计量特征及分布格局,并与国外、世界湿地土壤的生态化学计量比进行了对比分析,探索了影响湿地土壤的碳氮磷生态化学计量特征的因子。贺金生等第一次提岀生态化学计量学为将个体、群落、生态系统等不同尺度的生物系统统一联系起来提供了新的构思。 2土壤生态化学计量学研究现状 土壤作为植物生长的主要环境之一,为植物的生长提供所必需的养分;其储集的养分含量直接影响到植物个体的生长趋势以及植物群落的生产力。湿地土壤中的有机碳、氮、磷作为生态系统中主要的生源要素,不仅是生态系统物质循环的一个核心环节,还是影响湿地生态系统生产力的重要因子。Cleveland 等研究发现土壤中也存在类似于“Redfled比值”的碳氮磷比值,如全球土壤及土壤微生物的C:N:P比值相对稳定在186:13:1和60:7:1。Tian等发现在区域尺度上,土壤的C:N也保持在一个相对稳定的区域波动。目前,有关森林、草原、荒漠等地的土壤生态化学计量学已开展了较为全面的研究,张仲胜等问对中 国湿地土壤的生态化学计量特征统计发现,C:N、C:P及N:P,均远远高于全球土壤的平均水平。曹磊等对黄河三角洲潮汐湿 地C、N、P的化学计量学的特征研究发现,土壤中C、N、P的空间分布具有高度的异质性。但是基于湿地处在生物地球化学过程的热点区域,决定了湿地土壤环境的复杂性,有关湿地土壤碳、氮、磷等主要营养元素的化学循环机制及生态化学计量学特征的影响因素还有待进一步研究。 3植物的生态化学计量学研究现状 生态化学计量学为深入研究植物的碳、氮、磷与土壤中碳、 氮、磷的相关性以及限制性元素对植物生长的影响提供了新的 思路。目前研究学者大多从不同的系统、功能群和物种的角度 研究植物的生态化学计量学特征及影响因子,也有从区域尺度、群落水平、植物器官元素等方面开展了广泛的研究。Han等研究得出,我国陆生植物叶的平均N含量为18.6,与全球植物N 含量大体相近,而P的平均含量为1.21,明显低于全球的植物叶P含量的平均值,并推测P可能是我国植物生长的主要限制性营养元素。而国内研究主要集中在森林、草原、荒漠等陆生生态系统及植物器官的C、N、P生态化学计量特征。王维奇等对闽江河口湿地的C、N、P含量及季节动态开展了研究,发现活体植物、枯落物和土壤的生态计量比均表现出C:P>C:N>N:P的规律,而且植物活体部分与枯落物的C、N含量均高于土壤。 4枯落物的生态化学计量学研究现状 枯落物作为湿地生态系统关键的养分储存库,其养分的变 化特征会影响到整个生态系统的生产力及养分循环。枯落物的 分解过程是不仅涵盖了植物光合作用合成有机体的过程,还包 括衰亡组织有机体的分解过程。国外针对不同湿地生态系统的 枯落物分解过程及其影响因素、测定方法和模型建立等方面开 展了大量的研究工作,并取得了许多重要的研究成果。在枯落物的分解过程方面,研究学者从时间和空间的研究角度探索了 其物理、化学及生物过程;在模型建立方面,01so提出的单指数时间衰减模型被广泛用以预测枯落物的分解过程。在影响因素 方面,Roache等研究了盐度对湿地枯落物分解过程的影响,分析了盐度与枯落物分解速率的相关性。相比之下,国内有关湿地枯落物地研究还处在起步阶段,但是近年来发展迅速,先后有学者从枯落物的分解过程、分解速率和影响因子进行了大量的研究工作,研究区域包含了闽江河口湿地,三江平原和鄱阳湖湿地等湿地生态系统。 参考文献 [1]熊汉锋,王运华.湿地碳氮磷的生物地球化学循环研究进展[J].土壤通报,2005,36⑵:240-243. [2]罗美娟,黄炜娟,谭芳林等.闽江河口湿地主要植物(转下页)
专题二化学常用计量 1.(2012·高考课标全国卷)用N A表示阿伏加德罗常数的值.下列叙述中不正确的是() A.分子总数为N A的NO2和CO2混合气体中含有的氧原子数为2N A B.28 g乙烯和环丁烷(C4H8)的混合气体中含有的碳原子数为2N A C.常温常压下,92 g的NO2和N2O4混合气体含有的原子数为6N A D.常温常压下,22.4 L氯气与足量镁粉充分反应,转移的电子数为2N A 2.(2012·高考山东卷)下列与含氯化合物有关的说法正确的是() A.HClO是弱酸,所以NaClO是弱电解质 B.向沸水中逐滴加入少量饱和FeCl3溶液,可制得Fe(OH)3胶体 C.HCl溶液和NaCl溶液均通过离子导电,所以HCl和NaCl均是离子化合物 D.电解NaCl溶液得到22.4 L H2(标准状况),理论上需要转移N A个电子(N A表示阿伏加德罗常数) 3.(2012·高考广东卷)设n A为阿伏加德罗常数的数值,下列说法正确的是() A.常温下,4 g CH4含有n A个C—H共价键 B.1 mol Fe与足量的稀HNO3反应,转移2n A个电子 C.1 L 0.1 mol·L-1NaHCO3溶液中含有0.1n A个HCO-3 D.常温常压下,22.4 L的NO2和CO2混合气体含有2n A个O原子 4.(2012·高考江苏卷)设N A表示阿伏加德罗常数的值.下列说法正确的是() A.标准状况下,0.1 mol Cl2溶于水,转移的电子数目为0.1N A B.常温常压下,18 g H2O中含有的原子总数为3N A C.标准状况下,11.2 L CH3CH2OH中含有的分子数目为0.5N A D.常温常压下,2.24 L CO和CO2混合气体中含有的碳原子数目为0.1N A 5.(2012·高考福建卷)下列说法正确的是() A.0.5 mol O3与11.2 L O2所含的分子数一定相等 B.25 ℃与60 ℃时,水的pH相等 C.中和等体积、等物质的量浓度的盐酸和醋酸所消耗的n(NaOH)相等 D.2SO2(g)+O2(g)2SO3(g)和4SO2(g)+2O2(g)4SO3(g)的ΔH相等 6.(2012·高考四川卷)设N A为阿伏加德罗常数的值,下列叙述正确的是() A.标准状况下,33.6 L氟化氢中含有氟原子的数目为1.5N A B.常温常压下,7.0 g乙烯与丙烯的混合物中含有氢原子的数目为N A C.50 mL 18.4 mol/L浓硫酸与足量铜微热反应,生成SO2分子的数目为0.46N A D.某密闭容器盛有0.1 mol N2和0.3 mol H2,在一定条件下充分反应,转移电子的数目为0.6N A 专题二化学常用计量 1.【解析】选D.A项:NO2和CO2中都含相同数目的氧原子,故N A个NO2和CO2混合气体中含有的氧原子数为2N A. B项:乙烯和环丁烷的最简式为CH2,所以28 g乙烯和环丁烷的混合气体中含有碳原子的物质 的量为28 g 14 g/mol=2 mol,碳原子数目为2N A. C项:等质量的NO2与N2O4所含原子数目相等, 则92 g混合气体所含原子数目为: 92 g 46 g/mol×3=6 mol,数目为6N A. D项:当应用V m=22.4 L/mol时,应指明气体所在的状况为标准状况,而题目所给状况为常温常压,故不能应用V m=22.4 L/mol. 2.【解析】选B.NaClO属于盐为强电解质,A项错误;加热可促进Fe3+的水解,所以向沸水中滴入饱和氯化铁溶液可制得氢氧化铁胶体,B项正确;HCl属于共价化合物,C项错误;电解氯化钠溶液时,H+在阴极上的电极反应为2H++2e-====H2↑,生成1 mol氢气转移2N A个电子,D项错误. 3.【解析】选A.A项,1 mol CH4中含4 mol C—H键,4 g CH4的物质的量为0.25 mol,含C—H 键的物质的量为0.25 mol×4=1 mol,共n A个,故A正确.B项,HNO3过量时,铁被氧化为+3价,1 mol Fe转移3n A个电子,B项错误;C项,HCO-3在水溶液中既发生电离又发生水解,以水解为主,故c(HCO-3)<0.1 mol/L,HCO-3的个数小于0.1n A,C项错误;D项,气体所处的状态不是标准状况,故D错误. 4.【解析】选B.A项,由于Cl2溶于水存在平衡体系Cl2+H2O HCl+HClO,0.1 mol Cl2不可能全部与H2O反应,因此转移的电子数目无法计算.B项,18 g水为1 mol,含有3N A个原子,B项正确.C项,标准状况下CH3CH2OH为液态,D项气体不是在标准状况下,故C、D两选项均错误. 5.【解析】选C.A项中没有标明标准状况,11.2 L O2不一定是0.5 mol O2,故错误;不同温度下,水的电离程度不同,水中c(H+)不同,pH不同,B项错;等体积等浓度的盐酸和醋酸所含溶质的物质的量相等,所消耗的n(NaOH)相等,C项正确;D项ΔH与化学计量数有关,D项错误. 6.【解析】选B.标准状况下HF为液态,33.6 L不是1.5 mol,故A项错误.乙烯、丙烯的混合物分子通式为(CH2)n, 7.0 g烯烃混合物中CH2原子团为0.5 mol,故氢原子数目为N A,B项正确.C 项中只有浓H2SO4才与Cu加热反应,当反应一段时间浓H2SO4变稀后与Cu的反应停止,SO2的分子数目一定小于0.46N A,故C项错误.0.1 mol N2与0.3 mol H2在容器中存在化学平衡,故转移电子数目一定小于0.6N A,D项错误.