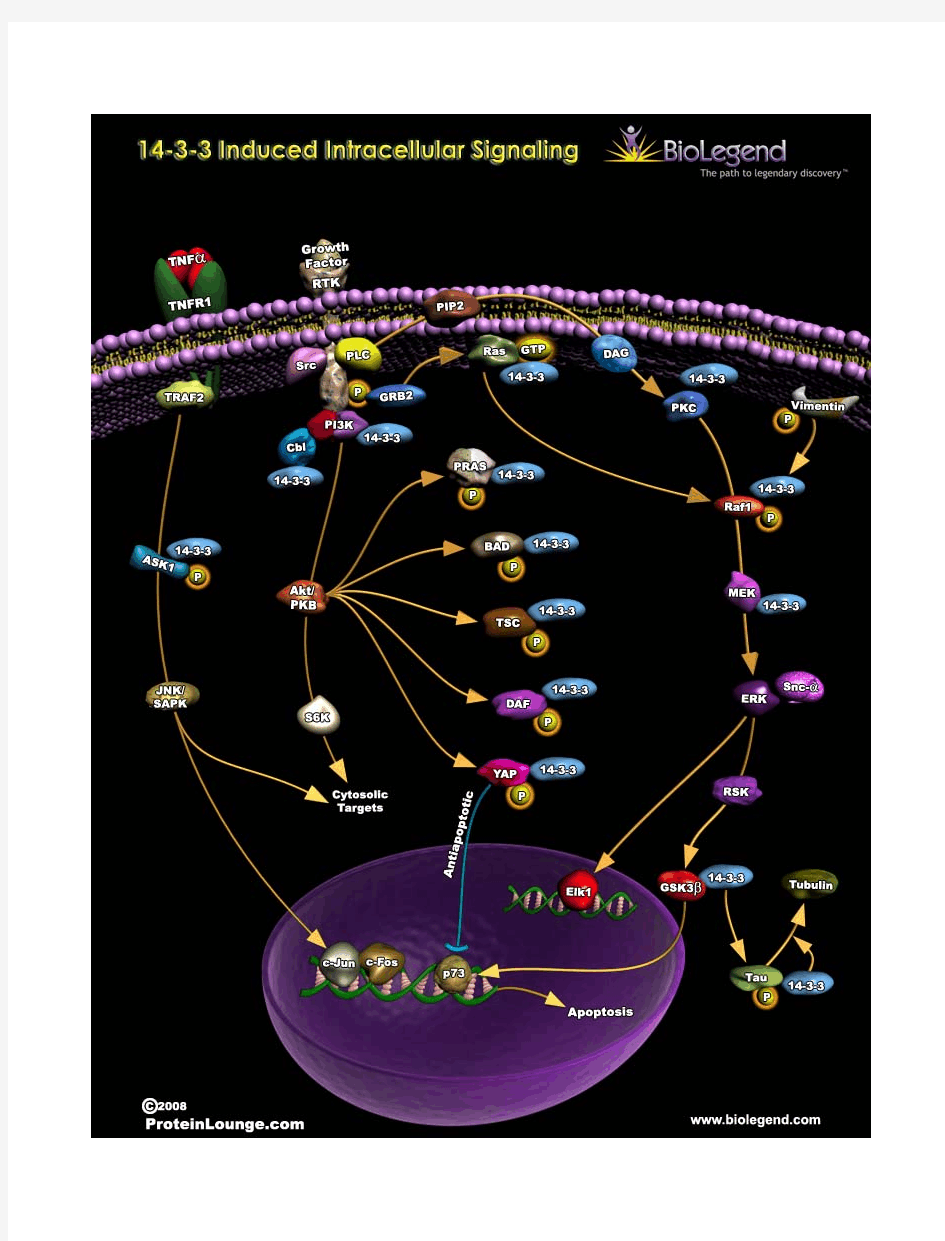
综述与进展 p38M APK信号转导通路与细胞凋亡研究进展 王誉霖1,张励才2 作者单位:1.安徽省宣城市人民医院麻醉科242000;2江苏徐州医学院作者简介: 王誉霖(1978,女,吉林市人,住院医师,硕士。研究方向:疼痛信号转导及调控。 主题词p38丝裂原活化蛋白激酶类;细胞凋亡;综述 中图分类号R345文献标识码A文章编号1674 8166(201012 1665 03 丝裂原活化蛋白激酶(mitog en2activated pr otein kinase,MA PK级联是细胞内广泛存在的丝/苏氨酸蛋白激酶超家族,是将细胞质的信号传递至细胞核并引起细胞核发生变化的重要物质。目前在人类已鉴定了4条MAPK途径:细胞外信号调节蛋白 激酶(ex tra cellular sig nal regulated protein kinase,ERK途径,C Jun 基末端激酶(c Jun N term inal kinase,JN K/应激活化蛋白(stress activated protein kinase,SAPK途 径,ERK5/大丝裂素活化蛋白激酶1(big MAP MAP kinase,BM K1途径和p38M APK(p38mitogen activated protein kinases,p38MA PK 传导途径[1]。p38 信号途径是 MAPK家族中的重要组成部分,多种炎症因子和生长因子及应激反应可使p38MAPK的酪氨酸和苏氨酸双磷酸化,从而激活p38M APK,使它在炎症、细胞应激、凋亡、细胞周期和生长等多种生理和病理过程中起重要作用。因此,p38MAPK 通路参与了多种刺激引起的信号级联反应,表明它在引起多种细胞反应中起重要作用,并且,p38在细胞凋亡中也有着重要的调节效应。1 p38M APK信号转导通路 丝裂原活化蛋白激酶(m ito gen activated pr otein kinase,MA PK级联是细胞内重 要的信号转导系统之一。在哺乳动物细胞M APK通路主要有:细胞外信号调节激酶(extracellular signal r eg ulated kinase,ERK ffi路、p38MA PK 通路、c jun 氨基末端激酶(c jun N term inal kinase,JNK通路和ERK5 通路[1]。其中,p38MAPK 是M APK 家族中的重要成员。
1 PPAR信号通路:过氧化物酶体增殖物激活受体( PPARs) 是与维甲酸、类固醇 和甲状腺激素受体相关的配体激活转录因子超家族核激素受体成员。它们作为脂 肪传感器调节脂肪代谢酶的转录。PPARs由PPARα、PPARβ和PPARγ 3种亚型组成。PPARα主要在脂肪酸代谢水平高的组织,如:肝、棕色脂肪、心、肾和骨骼肌表达。他通过调控靶基因的表达而调节机体许多生理功能包括能量代谢、生 长发育等。另外,他还通过调节脂质代谢的生物感受器而调节细胞生长、分化与 凋亡。PPARa同时也是一种磷酸化蛋白,他受多种磷酸化酶的调节包括丝裂原激活蛋白激酶( ERK-和p38.M APK) ,蛋白激酶A和C( PKA,PKC) ,AM PK和糖原合成酶一3( G SK3) 等调控。调控PPARa生长信号的酶报道有M APK、PKA和G SK3。PPARβ广泛表达于各种组织,而PPAR γ主要局限表达在血和棕色脂肪,其他组织如骨骼肌和心肌有少量表达。PPAR-γ在诸如炎症、动脉粥样硬化、胰岛素抵抗和糖代谢调节,以及肿瘤和肥胖等方面均有着举足轻重的作用, 而其众多生物学效应则是通过启动或参与的复杂信号通路予以实现。鉴于目前人 们对PPAR—γ信号通路尚不甚清,PPARs通常是通过与9-cis维甲酸受体( RXR)结合实现其转录活性的。 2 MAPK信号通路:mapk简介:丝裂原激活蛋白激酶(mitogen—activated protein kinase,MAPK)是广泛存在于动植物细胞中的一类丝氨酸/苏氨酸蛋白激酶。作用主要是将细胞外刺激信号转导至细胞及其核内,并引起细胞的生物化学反应(增殖、分化、凋亡、应激等)。 MAPKs家族的亚族 :ERKs(extracellular signal regulated kinase):包括ERK1、ERK2。生长因子、细胞因子或激素激活此通路,介导细胞增殖、分化。 JNKs(c-Jun N-terminal kinase)包括JNK1、JNK2、JNK3。此亚族成员能使 Jun转录因子N末端的两个氨基酸磷酸化而失活,因此称为Jun N末端激酶(JNKs)。物理、化学的因素引起的细胞外环境变化以及致炎细胞因子调节此通路。P38 MAPKs:丝氨酸/络氨酸激酶,包括p38 α、p38β、p38γ、p38δ。p38 MAP K参与多种细胞内信息传递过程 ,能对多种细胞外刺激发生反应,可磷酸化其它细胞质蛋白,并能从胞浆移位至细胞核而调节转录因子的活性来改变基因的表达水平 ,从而介导细胞生长、发育、分化及死亡的全过程。 ERK5:是一种非典型的MAPK通路,也叫大MAPK通路,只有一个成员。它可被各种刺激因素激活。不仅可以通过磷酸化作用使底物活化,并且通过C端的物理性结合作用激活底物。 3 ERBB信号途径:ErbB 蛋白属于跨膜酪氨酸激酶的 EGF 受体家族成员。ErbB 的命名来源于在禽红白血病 B( v-Erb-B) 发现的 EGF 受体的突变体,因而 EGF 受体 亦称为“ ErbB1”。人源 ErbB2 称为HER2, 特指人的 EGF 受体。ErbB 家族的
干货细胞信号通路图解之MAPK通路【值得珍藏】 科研小助手原创,转载请注明来源。公众号内回复“Cell Signaling Pathway”获取全套信号通路图本文由百度贴吧nosce吧吧主黄杰投稿一、MAPK信号通路: (1)有丝分裂原激活的蛋白激酶(MAPK)是一族在真核生物中非常保守的丝/苏氨酸蛋白激酶,在许多细胞活动中起作用,如生长增殖,细胞分化,细胞运动或死亡。MAPK级联信号传导由3 个不同层次的分子所组成。MAPK被MAPK的激 酶( MAPKK)磷酸化后激活,MAPKK被MAPKK的激酶(MAPKKK )磷酸化而激活。而MAPKKK通过与小GTPase 和/或其他蛋白酶相互作用而被激活,从而将MAPK和细胞 表面的受体以及胞外的信号联系在一起。 (2)许多参与生长和分化的受体都能够激活MAPK/ERK信号通路,比如说受体酪氨酸激酶(RTK),整合素,和离子通道。响应特定信号所涉及到的具体分子会相差很大,但通路的结构是一致的,那就是接头分子(adaptor,如Shc, GRB2, Crk等)将鸟苷酸交换因子(SOS, C3G 等)和受体连接在一起,然后把信号向小GTP 结合蛋白(Ras, Rap1)传递,后者又激活核心的级联反应,这是由一个MAPKKK( Raf) ,一个MAPKK( MEK1/2)和MAPK( Erk)所构成的。活化的ERK 二聚体能调节胞浆中的目标分子,也可以转移到细胞核中,然
后对一系列转录因子进行磷酸化以调节基因表达。SciRes(3)很多外部的刺激都能够激活G蛋白偶联受体(GPCR)。在受体活化以后,G 蛋白将GDP 转换成GTP ,然后结合了GTP的α和β/γ亚基从受体脱离开,启动信号向胞内的传导。与不同亚型的异质三聚体G 蛋白结合的受体可以采取不同 的手段激活小G 蛋白/MAPK级联反应,至少有三个不同家族的酪氨酸激酶参与其中。Src家族激酶响应活化的PI3Kγ,而后者被β/γ亚基激活。它们还能够响应受体的内化,受体酪氨酸激酶的交叉活化,以及有Pyk2 和/或FAK参与的整 合素途径信号。GPCRs同样可以通过PLCβ去激活PKC 和CaMKII ,对下游的MAPK通路可以有激活或抑制的影响。SciRes(4)压力激活的蛋白激酶(Stress-activated protein kinase, SAPK)或称Jun氨基端激酶(Jun amino-terminal kinase, JNK) 是MAPK的家族成员,能被一系列的环境压力,炎症细胞因子,生长因子和GPCR激动剂所激活。压力信号通过Rho家族的小GTP 酶(small GTPase)向这条级联通路传导,这些小GTP酶包括(Rac, Rho, cdc42) 。和其他的MAPK情况一样,靠近膜的激酶是一个MAPKKK,一般 是MEKK1-4 ,或者是一个混合激酶去磷酸化并激活 MKK4(SEK)或MKK7,它们是SAPK/JNK的激酶。另外,MKK4/7也可以被生发中心激酶(germinal center kinase, GCK)以一种GTPase 依赖的方式激活。活化后的
1JAK-STAT信号通路 1)JAK与STAT蛋白 JAK-STAT信号通路是近年来发现的一条由细胞因子刺激的信号转导通路,参与细胞的增殖、分化、凋亡以及免疫调节等许多重要的生物学过程。与其它信号通路相比,这条信号通路的传递过程相对简单,它主要由三个成分组成,即酪氨酸激酶相关受体、酪氨酸激酶JAK和转录因子STAT。(1)酪氨酸激酶相关受体(tyrosinekinaseassociatedreceptor) 许多细胞因子和生长因子通过JAK-STAT信号通路来传导信号,这包括白介素2?7(IL-2?7)、GM-CSF(粒细胞/巨噬细胞集落刺激因子)、GH(生 长激素)、EGF(表皮生长因子)、PDGF(血小板衍生因子)以及IFN(干扰素)等等。这些细胞因子和生长因子在细胞膜上有相应的受体。这些受体的共同特点是受体本身不具有激酶活性,但胞内段具有酪氨酸激酶JAK 的结合位点。受体与配体结合后,通过与之相结合的JAK的活化,来磷酸化各种靶蛋白的酪氨酸残基以实现信号从胞外到胞内的转递。 (2)酪氨酸激酶JAK(Januskinase) 很多酪氨酸激酶都是细胞膜受体,它们统称为酪氨酸激酶受体(receptor tyrosinekinase,RTK),而JAK却是一类非跨膜型的酪氨酸激酶。JAK是英文Januskinase的缩写,Janus在罗马神话中是掌管开始和终结的两面神。之所以称为两面神激酶,是因为JAK既能磷酸化与其相结合的细胞因子受体,又能磷酸、JAK1个成员:4蛋白家族共包括JAK结构域的信号分子。SH2化多个含特定
JAK2、JAK3以及Tyk2,它们在结构上有7个JAK同源结构域(JAKhomologydomain,JH),其中JH1结构域为激酶区、JH2结构域是“假”激酶区、JH6和JH7是受体结合区域。 (3)转录因子STAT(signaltransducerandactivatoroftranscription)STAT被称为“信号转导子和转录激活子”。顾名思义,STAT在信号转导和转录激活上发挥了关键性的作用。目前已发现STAT家族的六个成员,即STAT1-STAT6。STAT蛋白在结构上可分为以下几个功能区段:N-端保守序列、DNA结合区、SH3结构域、SH2结构域及C-端的转录激活区。其中,序列上最保守和功能上最重要的区段是SH2结构域,它具有与酪氨酸激酶Src的SH2结构域完全相同的核心序列“GTFLLRFSS”。 2)JAK-STAT信号通路 与其它信号通路相比,JAK-STAT信号通路的传递过程相对简单。信号传 递过程如下:细胞因子与相应的受体结合后引起受体分子的二聚化,这使得与受体偶联的JAK激酶相互接近并通过交互的酪氨酸磷酸化作用而活化。JAK激活后催化受体上的酪氨酸残基发生磷酸化修饰,继而这些磷酸化的酪氨酸位点与周围的氨基酸序列形成“停泊位点”(dockingsite),同时含有SH2结构域的STAT蛋白被招募到这个“停泊位点”。最后,激酶JAK 催化结合在受体上的STAT蛋白发生磷酸化修饰,活化的STAT蛋白以二 聚体的形式进入细胞核内与靶基因结合,调控基因的转录。值得一提的是,一种JAK激酶可以参与多种细胞因子的信号转导过程,一种细胞因子的信号通路也可以激活多个JAK激酶,但细胞因子对激活的STAT分子却具有一定的选择性。例如IL-4激活STAT6,而IL-12 。STAT4却特异性激活
目录 actin肌丝 (5) Wnt/LRP6 信号 (7) WNT信号转导 (7) West Nile 西尼罗河病毒 (8) Vitamin C 维生素C在大脑中的作用 (10) 视觉信号转导 (11) VEGF,低氧 (13) TSP-1诱导细胞凋亡 (15) Trka信号转导 (16) dbpb调节mRNA (17) CARM1甲基化 (19) CREB转录因子 (20) TPO信号通路 (21) Toll-Like 受体 (22) TNFR2 信号通路 (24) TNFR1信号通路 (25) IGF-1受体 (26) TNF/Stress相关信号 (27) 共刺激信号 (29) Th1/Th2 细胞分化 (30) TGF beta 信号转导 (32) 端粒、端粒酶与衰老 (33) TACI和BCMA调节B细胞免疫 (35) T辅助细胞的表面受体 (36) T细胞受体信号通路 (37) T细胞受体和CD3复合物 (38) Cardiolipin的合成 (40) Synaptic突触连接中的蛋白 (42) HSP在应激中的调节的作用 (43) Stat3 信号通路 (45) SREBP控制脂质合成 (46) 酪氨酸激酶的调节 (48) Sonic Hedgehog (SHH)受体ptc1调节细胞周期 (51) Sonic Hedgehog (Shh) 信号 (53) SODD/TNFR1信号 (56) AKT/mTOR在骨骼肌肥大中的作用 (58) G蛋白信号转导 (59) IL1受体信号转导 (60) acetyl从线粒体到胞浆过程 (62) 趋化因子chemokine在T细胞极化中的选择性表达 (63) SARS冠状病毒蛋白酶 (65) SARS冠状病毒蛋白酶 (67) Parkin在泛素-蛋白酶体中的作用 (69)
CELL DEATH AND CELL-CYCLE CHECKPOINT DURING DNA DAMAGE 细胞死亡及周期阻滞基本信号通路 有哪些因素可引起DNA损伤?DNA损伤的结局如何? (课件) (一)DNA损伤的原因 环境因素,化学因素,生物因素例如: UV ,离子辐射,基因毒性化学试剂引起ssDNA/dsDNA 损伤,DNA两条链交联或链内交联。正常细胞线粒体的一些代谢物(ROS)活泼氧类过多引起损伤。 (二) DNA损伤结局: 急性效应:干扰核酸代谢,触发细胞周期阻滞或死亡 长期效应:不可逆转突变导致肿瘤 细胞周期阻滞,衰老,肿瘤/癌症,有丝分裂危象 (一)DNA损伤的原因 1.DNA分子的自发性损伤 (1)DNA复制中的错误。 (2)DNA的自发性化学变化 a.碱基的异构互变性损伤 b.碱基的脱氨基作用 c.脱嘌呤与脱嘧啶 d.碱基修饰与链断裂 2.物理因素引起的DNA损伤 (1)紫外线引起的DNA损伤 (2)电离辐射引起的DNA损伤 a.碱基变化 b.脱氧核糖变化 c.DNA链断裂 d.交联 3.化学因素引起的DNA损伤 (1)烷化剂对DNA的损伤 a.碱基烷基化 b.碱基脱落 c.断链 d.交联 (2)碱基类似物、修饰剂对DNA的损伤 DNA损伤的后果 1.点突变(point mutation)指DNA上单一碱基的变异。嘌呤替代嘌呤(A与G之间的相互替代)、嘧啶替代嘧啶(C与T之间的替代)称为转换(transition);嘌呤变嘧啶或嘧啶变嘌呤则称为颠换(transvertion)。 2.缺失(deletion)指DNA链上一个或一段核苷酸的消失。 3.插入(insertion)指一个或一段核苷酸插入到DNA链中。在为蛋白质编码的序列中如缺失及插入的核苷酸数不是3的整倍数,则发生读框移动(reading frame shift),使其后所译读的 氨基酸序列全部混乱,称为移码突变(frame shift mutaion)。 4.倒位或转位(transposition)指DNA链重组使其中一段核苷酸链方向倒置、或从一处迁移到另一处。 5.双链断裂已如前述,对单倍体细胞一个双链断裂就是致死性事件。 (2)THE CONSEQUENCES OF DNA INJURY
山东农业大学学报(自然科学版),2015,46(4):514-518VOL.46N0.42015 Journal of Shandong Agricultural University(Natural Science Edition)doi:10.3969/j.issn.1000-2324.2015.04.007 细胞凋亡的信号通路 谢昆,李兴权 红河学院生命科学与技术学院,云南蒙自661199 摘要:细胞凋亡是细胞程序性死亡的一种方式,与自噬和坏死有明显的区别。细胞凋亡的信号途径比较复杂,在凋亡诱导因子的刺激下经历不同的信号途径。本文就细胞凋亡的三条信号通路——线粒体途径、内质网途径和死亡受体途径做一综述,以便为人们进一步了解细胞凋亡发生的机制,从而对癌症及其他一些相关疾病的治疗奠定基础。关键词:细胞凋亡;信号通路;线粒体途径;内质网途径;死亡受体途径 中图法分类号:R329.2+8文献标识码:A文章编号:1000-2324(2015)04-0514-05 The Signal Pathway of Apoptosis XIE Kun,LI Xing-quan Department of Life Science and Technology/Honghe University,Mengzi661199,China Abstract:Apoptosis is a process of programmed cell death which distinguishes from autophagy and necrosis.The signal pathways of apoptosis are complex and different under apoptosis induced factor stimulating.Three kinds of signal pathways of apoptosis including Mitochondrial pathway,Endoplasmic Reticulum pathway and Death Receptor pathway were summarized in this review in order to make people further comprehend the mechanism of apoptosis,so that it should make a basis for us all to treat cancer and other related diseases. Keywords:Apoptosis;signal pathway;Mitochondrial pathway;Endoplasmic Reticulum pathway;Death Receptor pathway 细胞凋亡是细胞程序性死亡(Program cell death,PCD)中特有的一种细胞死亡方式,是细胞在一系列内源性基因调控下发生的自然或生理性死亡过程。Kerr等1972年最早提出了凋亡(apoptosis)和坏死(necrosis)的概念[1],随后Paweletz等对其进行了详细的描述[2,3]。在形态学上,凋亡表现为核浓缩、细胞质密度增高、染色质凝聚、核膜破裂、核内DNA断裂、细胞集聚成团、形成凋亡小体(Apoptosome)等特征,这些凋亡小体最终被巨噬细胞清除,但不会引起周围细胞的炎症反应,另外,凋亡发生在单个细胞之间[4,5]。坏死,通常是由相邻的多个细胞之间发生细胞肿胀,细胞核溶解,细胞膜破裂,细胞质流入到细胞间质中,并伴发一系列的炎症反应,从而与凋亡表现为本质性区别[6,7]。 目前认为,凋亡发生的途径分为三种。第一种是线粒体途径,也称为内源性途径,该途径包括两类,第一类需要通过激活Caspase通路促进凋亡,在一序列凋亡诱导因素刺激下,线粒体中的Cyt C(细胞色素C)释放至细胞质中,从而与Apaf-1(Apoptosis protease activating factor1,凋亡蛋白酶活化因子1)结合形成多聚体,形成的多聚体再进一步与凋亡起始分子Caspase-9结合形成凋亡小体,凋亡小体激活Caspase-9,从而激活下游的凋亡执行分子Caspase-3,Caspase-6和Caspase-7等诱导细胞凋亡的级联反应;第二类是不依赖于Caspase途径的,通过线粒体释放AIF(Apoptosis induce factor,凋亡诱导因子)直接诱导凋亡的发生。但是在细胞内,直接检测AIF比较困难,而且AIF的变化不一定能代表凋亡发生的程度,因为引起凋亡发生的途径不一。第二种是死亡受体途径(也称为外源性途径),经由死亡受体(如TNF,Fas等)与FADD的结合而激活Caspase-8和caspase-10,进一步激活凋亡执行者caspase-3,6,7,从而促进凋亡的发生;第三条途径是内质网途径,内质网应激(蛋白质错误折叠或未折叠、内质网胁迫)会导致细胞内钙超载或钙离子稳态失衡一方面激活caspase-12,caspase-12进一步激活caspase-9而促进凋亡的发生,另一方面诱导Bcl-2(B细胞淋巴瘤蛋白)家族中促凋亡蛋白Bax和Bak的激活诱导凋亡[8]。 1凋亡的线粒体途径 在哺乳动物中,由于凋亡的激活需要线粒体中细胞色素C(CytC)的释放,因此CytC由线粒体膜间隙释放到细胞质中的多少可以作为判断凋亡发生强弱的指标之一。有研究认为,CytC的释放是通过Bcl-2家族调控线粒体膜透化(Mitochondrial outer membrane permeabilization,MOMP),科学 收稿日期:2013-03-07修回日期:2014-09-11 基金项目:云南省科技厅应用基础研究面上项目(2010ZC151) 作者简介:谢昆(1975-),男,云南富民人,博士研究生,研究方向为动物生物化学与分子生物学.E-mail:xk_biology2@https://www.doczj.com/doc/1a14467528.html, 数字优先出版:2015-06-03https://www.doczj.com/doc/1a14467528.html,
mTOR信号通路图 mTOR可对细胞外包括生长因子、胰岛素、营养素、氨基酸、葡萄糖等多种刺激产生应答。它主要通过PI3K/Akt/mTOR途径来实现对细胞生长、细胞周期等多种生理功能的调控作用。正常情况下,结节性脑硬化复合物-1(TSC-1)和TSC-2形成二聚体复合物,是小GTP 酶Rheb(Ras-homolog enriched in brain)的抑制剂,而Rheb是mTOR活化所必需的刺激蛋白,因此TSC-1/TSC-2在正常情况下抑制mTOR的功能。当Akt活化后,它可磷酸化TSC-2的Ser939和Thr1462,抑制了TSC-1/TSC-2复合物的形成,从而解除了对Rheb 的抑制作用,使得mTOR被激活。活化的mTOR通过磷酸化蛋白翻译过程中的某些因子来参与多项细胞功能,其中最主要的是4EBP1和P70S6K。
在整个PI3K/Akt/mTOR信号通路中,有一条十分重要的负反馈调节剂就是10号染色体上缺失与张力蛋白同源的磷酸酶基因(phosphatase and tensin homology deleted on chromosome 10, PTEN)。PTEN是一个肿瘤抑制基因,位于人染色体10q23。它有一个蛋白酪氨酸磷酸酶结构域,在这条通路中可以将PI-3,4-P2与PI-3,4,5-P3去磷酸化,从而负调节PI3K下游AKt/mTOR信号通路的活性。 本信号转导涉及的信号分子主要包括 IRS-1,PI3K,PIP2,PIP3,PDK1,PTEN,Akt,TSC1,TSC2,Rheb,mTOR,Raptor,DEPTOR,GβL,p70S6K,ATG13,4E-BP1,HIF-1,PGC-1α,PPARγ,Sin1,PRR5,Rictor,PKCα,SGK1,PRAS40,FKBP12,Wnt,LRP,Frizzled,Gαq/o,Dvl,Erk,RSK,GSK-3,REDD1,REDD2,AMPK,LKB1,RagA/B,RagC/D等。
KEGG上的信号通路图怎么看? 提示:请点击标题下方蓝色“实验万事屋”,添加关注后,发“嗯”可以查看我们之前的文章。未经允许,其他公众号不得转载哦! 想要把自己研究的分子扯上明星分子或者明星通路?那是不难,难的是具体到底要怎么去扯,芯片结果啊,生信结果啊,都会给你提示,但真的要具体扯上去,还得看懂那些七七八八的信号通路图。 KEGG Pathway上有着大量的信号通路图,画得一个复杂啊!巨坑爹有没有?曾经有师弟说我之前曾经把Wnt通路描述错了,他师兄告诉他,应该是GSK-3β磷酸化抑制β-Catenin降解,并促进它入核的。在这里,我们只能默默地祝福这位师兄了…… 那我们就用Wnt通路来做例子吧。先上KEGG下载一个Wnt的信号通路图,如下: 绝壁是很高大上的不是么?这要咋看呢?其实这张图上把三个Wnt通路都画上去了,也就是Wnt/β-Catenin(经典Wnt通路),Wnt/PCP(平面的细胞极性途径)和Wnt/Ca2+(Wnt/钙离子)三条信号通路组成,我们就删减一下,就光看经典的Wnt通路,就变成了下面这个模样:
感觉还是很高大上有木有?那就再删减一下,把它变成经典Wnt信号通路的骨架会是什么样呢?就是这样: 简洁明快了吧,但要怎么来看懂这样的图呢?我们来看一下KEGG Pathway的具体图例:
把这些图例用来解释经典Wnt信号通路骨架图,就变成了: 看懂了么?那给你从左到右解释一下: 1)Wnt激活膜上受体,将信号传递到第二信使Dvl,活化的Dvl抑制由Axin、APC 和GSK-3β组成的复合物的活性,使β-catenin不能被GSK-3β磷酸化。 2)磷酸化的β-catenin才可通过泛素化(ubiquitination)而被胞浆内的蛋白酶体所降解,由于非磷酸化的β-catenin不能被蛋白酶体降解,从而导致β-catenin在胞浆内积聚,并移向核内。
细胞凋亡信号转导途径及其调控的研究 进展 学科:基础兽医学 专业:药理毒理学 姓名:ma cai hui 学号:13203023
细胞凋亡信号转导途径及其调控的研究进展 摘要目的:为了研究抗肿瘤药物促使细胞凋亡的作用机理,探讨细胞凋亡的信号转导途径以及相关基因对其的调控。方法:查阅近年的国内外相关文献,归纳整理细胞凋亡的信号转导途径和相关的调控基因。结果:介绍了细胞凋亡存在三条主要通路:线粒体通路、内质网通路和死亡受体通路,各通路间互相联系,共同调节细胞凋亡。以及调控凋亡的主要基因,Bcl-2、p53、c-myc、P16、Rb。结论:研究抗肿瘤药物的作用机理应从以上三条凋亡途径和相关调控基因出发。 关键词细胞凋亡;信号转导途径;基因调控;caspase Progress study on signal transmission pathways and regulation of cell apoptosis Wang Saiqi School of Pharmaceutical Sciences, Zhengzhou University, Zhengzhou, 450001 Key words : cell apoptosis; signal transmission pathways; gene regulation; caspase Abstract Aim : To check the mechanism of apoptosis induced by anticarcinogen and research the cell apoptosis signal transmission pathways and related genes on its regulation. Methods: Signal transmission pathways and related genes were concluded by referring to related papers at home and abroad in recent years. Results: Three main signal transmission pathways, death receptor-mediated pathways, mitochondrial pathway, endoplasmic reticulum pathway and several main regulator genes,Bcl-2,p53, c-myc,P16,Rb were introduced. Conclusions: Research on the mechanism of anticarcinogen should start from the said signal transmission pathways and genes. 1 细胞凋亡概述 细胞凋亡,又名细胞程序性死亡,是诱导性的细胞自杀过程,它使生物体可以有序地清除受损伤或无用的细胞。自从1927年John Kerr第一次提出凋亡这一概念后,人们发现它在多细胞生物的基本生命活动中起着十分重要的作用。它对于
细胞凋亡的过程大致可分为以下几个阶段:接受凋亡信号→凋亡调控分子间的相互作用→蛋白水解酶的活化(Caspase)→进入连续反应过程细胞凋亡的启动是细胞在感受到相应的信号刺激后胞内一系列控制开关的开启或关闭,不同的外界因素启动凋亡的方式不同,所引起的信号转导也不相同,客观上说对细胞凋亡过程中信号传递系统的认识还是不全面的,比较清楚的通路主要有:1)细胞凋亡的膜受体通路:各种外界因素是细胞凋亡的启动剂,它们可以通过不同的信号传递系统传递凋亡信号,引起细胞凋亡,我们以Fas -FasL为例:Fas是一种跨膜蛋白,属于肿瘤坏死因子受体超家族成员,它与FasL结合可以启动凋亡信号的转导引起细胞凋亡。它的活化包括一系列步骤:首先配体诱导受体三聚体化,然后在细胞膜上形成凋亡诱导复合物,这个复合物中包括带有死亡结构域的Fas相关蛋白FADD。Fas又称CD95,是由325个氨基酸组成的受体分子,Fas一旦和配体FasL结合,可通过Fas分子启动致死性信号转导,最终引起细胞一系列特征性变化,使细胞死亡。Fas作为一种普遍表达的受体分子,可出现于多种细胞表面,但FasL的表达却有其特点,通常只出现于活化的T细胞和NK细胞,因而已被活化的杀伤性免疫细胞,往往能够最有效地以凋亡途径置靶细胞于死地。Fas分子胞内段带有特殊的死亡结构域(DD,death domain)。三聚化的Fas和FasL结合后,使三个Fas分子的死亡结构域相聚成簇,吸引了胞浆中另一种带有相同死亡结构域的蛋白FADD。FADD是死亡信号转录中的一个连接蛋白,它由两部分组成:C端(DD结构域)和N端(DED)部分。DD结构域负责和Fas分子胞内段上的DD结构域结合,该蛋白再以DED连接另一个带有DED的后续成分,由此引起N段DED随即与无活性的半胱氨酸蛋白酶8(caspase8)酶原发生同嗜性交联,聚合多个caspase8的分子,caspase8分子遂由单链酶原转成有活性的双链蛋白,进而引起随后的级联反应,即Caspases,后者作为酶原而被激活,引起下面的级联反应。细胞发生凋亡。因而TNF诱导的细胞凋亡途径与此类似2)细胞色素C释放和Caspases激活的生物化学途径线粒体是细胞生命活动控制中心,它不仅是细胞呼吸链和氧化磷酸化的中心,而且是细胞凋亡调控中心。实验表明了细胞色素C从线粒体释放是细胞凋亡的关键步骤。释放到细胞浆的细胞色素C在dATP存在的条件下能与凋亡相关因子1(Apaf-1)结合,使其形成多聚体,并促使caspase-9与其结合形成凋亡小体,caspase-9被激活,被激活的caspase-9能激活其它的caspase如caspase-3等,从而诱导细胞凋亡。此外,线粒体还释放凋亡诱导因子,如AIF,参与激活caspase。可见,细胞凋亡小体的相关组份存在于正常细胞的不同部位。促凋亡因子能诱导细胞色素C 释放和凋亡小体的形成。很显然,细胞色素C从线粒体释放的调节是细胞凋亡分子机理研究的关键问题。多数凋亡刺激因子通过线粒体激活细胞凋亡途经。有人认为受体介导的凋亡途经也有细胞色素C从线粒体的释放。如对Fas应答的细胞中,一类细胞(type1)中含有足够的胱解酶8 (caspase8)可被死亡受体活化从而导致细胞凋亡。在这类细胞中高表达Bcl-2并不能抑制Fas诱导的细胞凋亡。在另一类细胞(type2)如肝细胞中,Fas受体介导的胱解酶8活化不能达到很高的水平。因此这类细胞中的凋亡信号需要借助凋亡的线粒体途经来放大,而Bid -- 一种仅含有BH3结构域的Bcl-2家族蛋白是将凋亡信号从胱解酶8向线粒体传递的信使。尽管凋亡过程的详细机制尚不完全清楚,但是已经确定Caspase即半胱天冬蛋白酶在凋亡过程中是起着必不可少的作用,细胞凋亡的过程实际上是Caspase不可逆有限水解底物的级联放大反应过程,到目前为止,至少已有14种Caspase被发现,Caspase分子间的同源性很高,结构相似,都是半胱氨酸家族蛋白酶,根据功能可把Caspase基本分为二类:一类参与细胞的加工,如Pro-IL-1β和Pro-IL-1δ,形成有活性的IL-1β和IL-1δ;第二类参与细胞凋亡,包括caspase2,3,6,7,8,9.10。Caspase家族一般具有以下特征:1)C端同源区存在半胱氨酸激活位点,此激活位点结构域为QACR/QG。2)通常以酶原的形式存在,相对分子质量29000-49000(29-49KD),在受到激活后其内部保守的天冬氨酸残基经水解形成大(P20)小(P10)两个亚单位,并进而形成两两组成的有活性的四聚体,其中,每个P20/P10异二聚体可来源于同一前体分子也可来源于两个不同的前体分子。3)末端具有一个小的或大的原结构域。参与诱导凋亡的Caspase分成两大类:启动酶(inititaor)和效应酶(effector)它们分别在死亡信号转导的上游和下游发挥作用。
一 MAPK信号转导途径 MAPK途径的基本组分 MAPK级联反应包含三个顺序激活的成分:MAPK激酶的激酶(MAPKKK或MEKK),MAPK 激酶(MAPKK,MKK 或MEK) 和MAPK [1]。目前在人类主要有三组MAPK通路:ERK1/2(细胞外信号调节激酶)MAPK家族,P38MAPK家族,JNK/SAPK(c-Jun 氨基端激酶/应激活化蛋白激酶)MAPK 家族[2]。 1.1 ERK1/2家族ERK1/2信号通路包括五个亚组,ERK1/2,ERK3/4和ERK5[3]。ERK1 /2 与细胞增殖最为密切,其上游激酶为MAPK 激酶(MEK1/2), MEK1与细胞分化有关,而MEK2 与细胞增殖有关[4]。 1.2 JNK/SAPK MAPK家族外界刺激可通过Ras依赖或非Ras依赖的两条途径激活JNK[6]。已有研究证实,双特异性激酶JNK Kinase(JNKK)是JNK/S A P K的上游激活物,其中M K K7/JNKK2可特异性地激活JNK[5],MKK4则可同时激活JNK1和p38。 1.3 P38MAPK家族p38是由360个氨基酸组成的38kD的蛋白,与JNK 同属应激激活的蛋白激酶。研究表明,在许多细胞反应中发现P38 活化,并且与细胞种类及外界刺激有关。p38MAPK 通路可 被应激刺激(Uv、H 2O 2 、热休克和缺氧 等)、炎性因子(TNF-α、IL-1 和FGF 等) 及LPS 和革兰氏阳性细菌细胞壁成分而激活[7,8]。SKF86002 是第一个报道的P38M A P K抑制剂,以后又出现了SB203580 和其他的2 ,4 ,5 -三芳基咪唑, 它们能够特异性地抑制P38 MAPKα和P38 MAPKβ,而不影响JNK和ERK 的活性[9]。二 并行的MAPKs 信号通路在 细胞信号转导中的协调作用 研究表明,哺乳类细胞可通过多种机制维持其每一条MAPKs信号通路信号转导的特异性。Schaeffer和Whitmarsh 等人报告在哺乳类细胞中也存在着类似于真菌的支架蛋白[10]。此外,在哺乳类细胞中并行的MAPKs信号通路对细胞信号转导具有协调作用。有研究证实,在成纤维细胞中,激活SAPK的刺激可以诱导MKP-1基因的表达,但激活ERK的刺激并无此作用,提示这二条通路之间具有相互调控,这一调节机制的存在可使细胞特异地对激活SAPK通路的刺激发生反应[11]。此外还有学者报告,J N K及ERK均可磷酸化转录因子Elk-1,促进TCF的形成、增加SRE的转录活性,提示SRE是这两条通路的汇合点,表明细胞可对不同的细胞外刺激信号进行整合,最终产生协调的生物学反应[12]。 三 MAPK 通路与细胞凋亡 3.1 显性失活(dominant-negative)Ras、Raf-1突变体可以抑制细胞增殖,而持续激活的Raf-1可介导细胞增殖;同样,显性失活MEK突变体或持续激活的MEK 分别抑制或促进NIH3T3细胞的增殖;突变的ERK或其反义cDNA可抑制细胞增殖[13]。 3.2J u r k a t细胞经γ射线处理后JNK可被激活,出现细胞凋亡,而当细胞被转染了显性失活JNK突变体后,γ射线诱导的凋亡可以被阻断[15]。以上研究表明,JNK的激活可诱导细胞发生凋亡。JNK 激活方式的不同也可产生不同的生物学效应,γ射线照射Jurkat T细胞后,JNK 被持续激活,细胞发生凋亡;而CD28单 DOI:10.3969/j.issn.1001-8972.2010.14.092 MAPK信号通路与细胞凋亡的关系王利东 韩冲 吴冬金 中国医科大学
1PPAR信号通路:过氧化物酶体增殖物激活受体(PPARs)是与维甲酸、类固醇和甲状腺激素受 体相关的配体激活转录因子超家族核激素受体成员。它们作 为脂肪传感器调节脂肪代谢酶的转录。PPARs由PPARα、PPARβ和PPARγ3种亚型组成。PPARα主要在脂肪酸代谢水平高的组织,如:肝、棕色脂肪、心、肾和骨骼肌表达。他通过 调控靶基因的表达而调节机体许多生理功能包括能量 代谢、生长发育等。另外,他还通过调节脂质代谢的生物感受器而调节细胞生长、分化与凋亡。PPARa同时也是一种磷酸化蛋白,他受多种磷酸化酶的调节包括丝裂原激活蛋白激酶(ERK-和 p38.MAPK),蛋白激酶A和C(PKA,PKC),AMPK和糖原合成酶一3(GSK3)等调控。调控PPARa 生长信号的酶报道有MAPK、PKA和GSK3。PPARβ广泛表达于各种组织,而PPARγ主 要局限表达在血和棕色脂肪,其他组织如骨骼肌和心肌有少量表达。PPAR-γ在诸如炎症、动 脉粥样硬化、胰岛素抵抗和糖代谢调节,以及肿瘤和肥胖等方面 均有着举足轻重的作用,而其众多生物学效应则是通过启动或参与的复杂信号 通路予以实现。鉴于目前人们对PPAR—γ信号通路尚不甚清,PPARs通常是通过与9-cis维 甲酸受体(RXR)结合实现其转录活性的。 2MAPK信号通路:mapk简介:丝裂原激活蛋白激酶(mitogen—activatedproteinkinase,MAPK)是广泛存在于动植物细胞中的一类丝氨酸/苏氨酸蛋 白激酶。作用主要是将细胞外刺激信号转导至细胞及其核内,并引起细胞的生物化学反 应(增殖、分化、凋亡、应激等)。 MAPKs家族的亚族:ERKs(extracellularsignalregulatedkinase) :包括 ERK1、ERK2。生长因子、细胞因子或激素激活此通路,介导细胞增殖、分化。 JNKs(c-JunN-terminalkinase)包括JNK1、JNK2、JNK3。此亚族成员能使Jun转录因子N末 端的两个氨基酸磷酸化而失活,因此称为JunN末端激酶(JNKs)。物理、化学的因素引起的 细胞外环境变化以及致炎细胞因子调节此通 路。 P38MAPKs:丝氨酸/络氨酸激酶,包括p38α、p38β、p38γ、p38δ。p38MAPK参与多种细胞内信息传递过程,能对多种细胞外刺激发生反应,可磷酸化 其它细胞质蛋白,并能从胞浆移位至细胞核而调节转录因子的活性来改变基因的表达水平,从而 介导细胞生长、发育、分化及死亡的全过程。 ERK5:是一种非典型的MAPK通路,也叫大MAPK通路,只有一个成员。它可 被各种刺激因素激活。不仅可以通过磷酸化作用使底物活化,并且通过C端的物理性结合作用 激活底物。 3ERBB信号途径:ErbB蛋白属于跨膜酪氨酸激酶的EGF受体家族成员。ErbB的命名来源于在禽 红白血病B(v-Erb-B)发现的EGF受体的突变体,因而EGF
Toll样受体信号通路图 TLR家族成员(TLR3除外)诱导的炎症反应都经过一条经典的信号通路(图1),该通路起始于TLRs的一段胞内保守序列—Toll/IL-1受体同源区(Toll/IL-1receptorhomologousregion,TIR).TIR可激活胞内的信号介质—白介素1受体相关蛋白激酶(IL-1Rassociatedkinase,IRAK)IRAK-1和IRAK-4、肿瘤坏死因子受体相关因子6(TNFR-associatedfactor6,TRAF-6)、促分裂原活化蛋白激酶(mitogenactivatedproteinkinase,MAPK)和IκB激酶(IκBkinase,IκK),进而激活核因子κB(nuclearfactorκB,NF-κB),诱导炎症因子的表达。 Toll-liker Receptor Signaling 本信号转导涉及的信号分子主要包括: CD14,MD-2,TRAM,TRIF,TIRAP,MyD88,TLR1,TLR2,TLR3,TLR4,TLR5,TLR6,TLR7,TLR8,TLR9,IRAK-1,IRAK-2,IRAK-4,IRAK-M,TRAF6,TRIAD3A,ST2L,SOCS1,RIG-I,FADD,TOLLIP,RIP1,A20,UEV1A,Ubc13,ECSIT,MEKK-1,TAK1,
TBK1,MKK3/6,p38,TAB1/2,MKK4/7,JNK,IKKα,IKKβ,IKKγ,IKKε,NEMO,IκBα,NF-κB,p65/RelA,Casp-8,IRF-3,IRF-7,MA VS等
最新细胞各种信号通路《Cell》 SnapShots are handy reference guides, carefully designed to highlight the key information on a particular topic on one page. SnapShots present up-to-date tables of nomenclature and glossaries, full signaling pathways, and schematic diagrams of cellular processes.Snapshots in red are FREE[/B]. Actin Regulators I[/url] Actin Regulators II[/url]
Antibiotic Inhibition of Protein Synthesis I[/url] Antibiotic Inhibition of Protein Synthesis II[/url] ENHANCED[/url]
Auxin Signaling and Transport Bacterial Appendages I Bacterial Appendages II B7/CD28 Costimulation
BCL-2 Proteins Ca2+-Calcineurin-NF A T Signaling
Ca2+-Dependent Transcription in Neurons Cell-Cycle Regulators I