DNA分子标记研究进展
- 格式:doc
- 大小:28.00 KB
- 文档页数:10
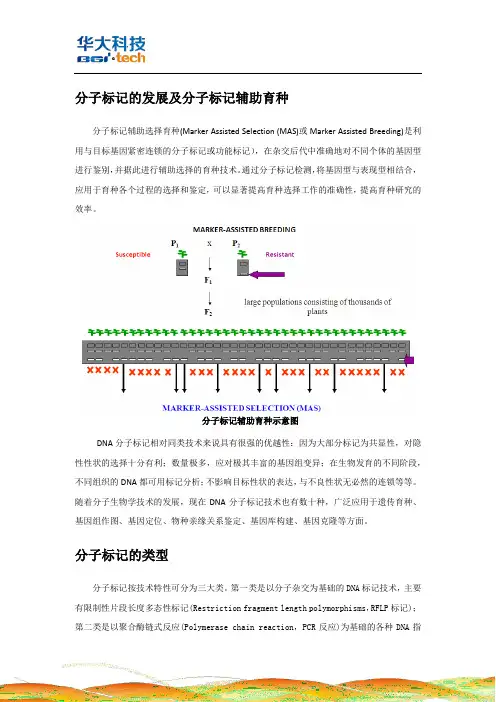
分子标记的发展及分子标记辅助育种分子标记辅助选择育种(Marker Assisted Selection (MAS)或Marker Assisted Breeding)是利用与目标基因紧密连锁的分子标记或功能标记),在杂交后代中准确地对不同个体的基因型进行鉴别,并据此进行辅助选择的育种技术。
通过分子标记检测,将基因型与表现型相结合,应用于育种各个过程的选择和鉴定,可以显著提高育种选择工作的准确性,提高育种研究的效率。
分子标记辅助育种示意图DNA分子标记相对同类技术来说具有很强的优越性:因为大部分标记为共显性,对隐性性状的选择十分有利;数量极多,应对极其丰富的基因组变异;在生物发育的不同阶段,不同组织的DNA都可用标记分析;不影响目标性状的表达,与不良性状无必然的连锁等等。
随着分子生物学技术的发展,现在DNA分子标记技术也有数十种,广泛应用于遗传育种、基因组作图、基因定位、物种亲缘关系鉴定、基因库构建、基因克隆等方面。
分子标记的类型分子标记按技术特性可分为三大类。
第一类是以分子杂交为基础的DNA标记技术,主要有限制性片段长度多态性标记(Restriction fragment length polymorphisms,RFLP标记);第二类是以聚合酶链式反应(Polymerase chain reaction,PCR反应)为基础的各种DNA指纹技术;第三类是一些新型的分子标记,如单核苷酸多态性(Single nucleotide polymorphism,SNP),由基因组核苷酸水平上的变异引起的DNA序列多态性,包括单碱基的转换、颠换以及单碱基的插入/缺失等。
分子标记是以DNA多态性为基础,因而具有以下优点:①表现稳定,多态性直接以DNA 形式表现,无组织器官、发育时期特异性,不受环境条件、基因互作影响;②数量多,理论上遍及整个基因组;③多态性高,自然界存在许多等位变异,无需专门人为创造特殊遗传材料,这为大量重要性状基因紧密连锁的标记筛选创造了条件;④对目标性状表达无不良影响,与不良性状无必然连锁;⑤部分标记遗传方式为共显性,可鉴别纯合体与杂合体;⑥成本不高,一般实验室均可进行。

dna分子标记技术及其在蔬菜遗传育种研究中的应用
DNA分子标记技术是一种通过分析DNA序列上的特定标记位点来研究物种的遗传变异和亲缘关系的技术。
在蔬菜遗传育种研究中,DNA分子标记技术被广泛应用于以下方面:
1. 遗传多样性研究:DNA分子标记技术可以通过分析不同蔬菜品种或不同个体之间的DNA序列差异来评估物种的遗传多样性。
通过比较不同品种或个体之间的DNA分子标记,可以确定它们之间的亲缘关系和遗传距离。
2. 基因定位和图谱构建:DNA分子标记技术可以用来帮助研究人员定位蔬菜的重要遗传特征或性状的基因。
通过分析与目标性状相关联的DNA分子标记的位置,可以确定这些标记位点与目标基因的连锁关系,并构建相应的遗传图谱。
3. 品种鉴定和纯度鉴定:DNA分子标记技术可以用来对蔬菜品种进行鉴定和纯度测试。
通过与已知标准品种的DNA序列进行比对,可以确定蔬菜品种的基因组组成,并判断其纯度和真实性。
4. 分子辅助选择育种:DNA分子标记技术可以与传统育种方法相结合,进行分子辅助选择育种。
通过对目标性状相关的DNA分子标记进行筛选、分析和评价,可以在早期育种阶段就有效地选择与目标性状相关的优良个体,提高育种效率。
总之,DNA分子标记技术在蔬菜遗传育种研究中发挥重要作
用,可以帮助研究人员分析遗传多样性、定位遗传特征、鉴定品种和辅助选择育种,为蔬菜遗传改良提供科学依据。


江苏农业学报(JiangsuJ.ofAgr.Sci.)ꎬ2024ꎬ40(1):183 ̄192http://jsnyxb.jaas.ac.cn孙雨桐ꎬ刘德帅ꎬ冯㊀美ꎬ等.果树分子标记辅助育种研究进展[J].江苏农业学报ꎬ2024ꎬ40(1):183 ̄192.doi:10.3969/j.issn.1000 ̄4440.2024.01.020果树分子标记辅助育种研究进展孙雨桐ꎬ㊀刘德帅ꎬ㊀冯㊀美ꎬ㊀齐㊀迅ꎬ㊀姚文孔(宁夏大学农学院/宁夏优势特色作物现代分子育种重点实验室/林木资源高效生产全国重点实验室ꎬ宁夏银川750021)收稿日期:2023 ̄02 ̄25基金项目:宁夏回族自治区农业育种项目(NXNYYZ202101)ꎻ宁夏回族自治区重点研发项目(2018BEB04004)作者简介:孙雨桐(1998-)ꎬ女ꎬ黑龙江五常人ꎬ硕士研究生ꎬ研究方向为果树学ꎮ(E ̄mail)syt15146063010@163.com通讯作者:姚文孔ꎬ(E ̄mail)yaowenkong@163.com㊀㊀摘要:㊀随着分子生物学的不断发展ꎬ分子标记在果树育种中发挥的作用也愈发重要ꎮ本文主要对不同果树育种的分子标记类型及分子标记在果树种质资源鉴定㊁抗性育种㊁无核育种㊁早熟育种㊁品质改良育种㊁分子遗传图谱构建与数量性状座位(QTL)基因定位等方面的应用进行了综述ꎬ为果树分子标记辅助育种提供参考ꎮ关键词:㊀果树ꎻ分子标记ꎻ遗传图谱ꎻQTL定位中图分类号:㊀S603.6㊀㊀㊀文献标识码:㊀A㊀㊀㊀文章编号:㊀1000 ̄4440(2024)01 ̄0183 ̄10Advancesofmolecularmarkersassistedselectionappliedinfruittreebreed ̄ingSUNYu ̄tongꎬ㊀LIUDe ̄shuaiꎬ㊀FENGMeiꎬ㊀QIXunꎬ㊀YAOWen ̄kong(SchoolofAgricultureꎬNingxiaUniversity/NingxiaKeyLaboratoryofModernMolecularBreedingofDominantandCharacteristicCrops/StateKeyLabora ̄toryofEfficientProductionofForestResourcesꎬYinchuan750021ꎬChina)㊀㊀Abstract:㊀Withthedevelopmentofmolecularbiologyꎬmolecularmarkershaveplayedmoreimportantrolesinthebreedingoffruittrees.Inthispaperꎬwemainlyexpoundedthetypesofmolecularmarkersindifferentfruittreesbreedingꎬandsummarizedtheapplicationofmolecularmarkersintheidentificationofgermplasmresourcesꎬresistancebreedingꎬseedlessbreedingꎬearlymaturitybreedingꎬqualityimprovementbreedingꎬmoleculargeneticmapconstructionandquanti ̄tativetraitlocus(QTL)mappinginfruittrees.Ourstudycanprovidereferenceformolecularmarkerassistantbreedinginfruittrees.Keywords:㊀fruittreesꎻmolecularmarkersꎻgeneticmappingꎻQTLlocalization㊀㊀果树育种的常规手段主要有杂交育种㊁诱变育种㊁倍性育种等ꎬ但由于果树杂种比较多㊁品种来源复杂㊁多为多年生木本植物㊁生长周期长等因素ꎬ导致传统育种周期长㊁效率低㊁不确定因素多ꎮ相比于常规育种ꎬ分子标记辅助选择(Molecularmarkeras ̄sistedselectionꎬMAS)可以提高育种效率ꎬ缩短育种周期ꎬ并且在遗传多样性㊁品种鉴定等方面也有较好的效果[1]ꎮ分子标记技术种类繁多ꎬ如随机扩增多态性DNA(RandomamplifiedpolymorphicDNAꎬRAPD)㊁扩增片段长度多态性(AmplifiedfragmentlengthpolymorphismꎬAFLP)㊁简单序列重复(SimplesequencerepeatꎬSSR)㊁单核苷酸多态性(Singlenu ̄cleotidepolymorphismꎬSNP)等ꎬ这些分子标记被广泛应用于果树的遗传育种㊁亲缘关系判断㊁遗传图谱构建以及数量性状座位(QTL)定位等研究中ꎮ这将加速果树品种的改良进程ꎬ提高育种效率ꎮ3811㊀果树上应用分子标记的主要类型1.1㊀RAPD分子标记为适应不良环境以及抵御病虫危害ꎬ培育具有一定抗性的果树品种至关重要ꎮTartarini[2]从5种不同的RAPD标记中筛选出与MdVf基因紧密相关的OPAM192200和OPAL07580ꎬ标记了苹果(Malusdo ̄mestica)抗赤霉病Vf基因ꎮ杨亚州等[3]以燕山葡萄和河岸葡萄的F1代为材料ꎬ通过7个RAPD标记对其抗旱性进行研究ꎬ将RAPD标记5226 ̄1100转化成专一性的SCAR(Sequencecharacterizedamplifiedregionsꎬ特定序列扩增)标记DR ̄760ꎮ其次RAPD分子标记多用于果树遗传多样性及品种鉴定ꎬ余智城等[4]对16份柑橘包括11份琯溪蜜柚进行遗传多样性分析ꎬ通过15条RAPD引物将16份材料分为2大类(表1)ꎮ表1㊀随机扩增多态性DNA(RAPD)分子标记在果树上的应用Table1㊀ApplicationofrandomamplifiedpolymorphicDNA(RAPD)molecularmarkersonfruittrees物种(属)功能主要结果参考文献苹果(MalusMill.)筛选抗病基因从5种不同的RAPD标记中筛选出与Vf基因紧密相关OPAM192200和OPAL07580标记苹果抗赤霉病Vf基因[2]葡萄(VitisL.)筛选抗旱基因以燕山葡萄和河岸葡萄的F1代为材料ꎬ通过7个RAPD标记对其抗旱性进行研究ꎬ获得了抗旱基因的RAPD标记[3]柑橘(CitrusL.)遗传多样性分析以16份柑橘包括11份琯溪蜜柚为材料用15条RAPD引物对其遗传多样性进行分析ꎬ将16份材料分为2大类[4]李(PrunusL.)绘制遗传图谱利用RFLP(限制性内切酶片段长度多态性)和RAPD标记绘制桃的遗传连锁图谱[5]品种鉴定使用360个RAPD引物进行大片段分析ꎬ以鉴定桃与桃㊁桃与杏仁杂交中特定位点相关的标记[6]遗传多样性分析用RAPD分子标记技术对7个樱桃品种进行多态性分析ꎬ将7个樱桃品种分为4类[7]杧果(MangiferaL.)遗传多样性分析从20个RAPD引物中筛选15个引物ꎬ分析了34份传统杧果种质的遗传变异和亲缘关系[8]1.2㊀AFLP分子标记分析果树遗传多样性ꎬ有助于果树的分类ꎮ董美超等[9]针对90份鳄梨品种材料从24对AFLP引物中筛选出8对进行遗传多样性分析ꎬ根据遗传相似系数可划分为4个类群ꎮLai等[10]采用AFLP和甲基化敏感扩增多态性(Methylationsensitiveampli ̄fiedpolymorphismꎬMSAP)的分子标记技术探究车道晚脐橙与芽变南瓜状脐橙之间的基因和基因组甲基化差异ꎬ结果表明芽变南瓜状脐橙的出现是由于基因突变ꎮ可见AFLP结合MSAP分子标记技术可以研究橙的早熟及果实形状的基因突变ꎬ这为此方面其他果树的研究提供了理论及技术的支持(表2)ꎮ表2㊀扩增片段长度多态性(AFLP)分子标记在果树上的应用Table2㊀Applicationofamplifiedfragmentlengthpolymorphism(AFLP)molecularmarkersonfruittrees物种(属)㊀㊀㊀功能㊀主要结果参考文献鳄梨(PerseaMill.)遗传多样性分析为了研究90份鳄梨品种材料ꎬ从24对AFLP引物中筛选出8对进行遗传多样性分析ꎬ根据遗传相似系数可划分为4个类群[9]苹果(MalusMill.)遗传多样性分析用AFLP对泰山红星㊁沂蒙短枝红星㊁红星和金冠4个苹果品种进行了遗传分析ꎬ结果表明ꎬ泰山红星苹果多态性显著高于其他品种[11]柑橘(CitrusL.)筛选果实形状基因采用AFLP和MSAP分析车道晚脐橙及其芽变南瓜状脐橙的基因和基因组甲基化差异ꎬ结果表明基因突变发生在车道晚脐橙和其芽变南瓜状脐橙之间[10]遗传多样性分析用AFLP技术对3个亲本和16个辐射诱变和芽变的柑橘品种进行了遗传差异分析[12]筛选早熟基因以脐橙试验材料ꎬ用AFLP和MSAP对其在基因组㊁甲基化修饰水平与脐橙早熟性状之间的关系进行探究ꎬ结果表明ꎬ果实熟期与去甲基化相关[13]猕猴桃(ActinidiaLindl.)遗传多样性分析使用AFLP技术对33份猕猴桃的遗传多样性进行分析ꎬ可以将33份种质分为3个类群[14]MSAP:甲基化敏感扩增多态性ꎮ1.3㊀SSR分子标记SSR标记的优点是大量标记及其共显性遗传ꎬ也正因如此SSR分子标记被广泛应用在果树育种中ꎮ王立新等[15]从144对SSR引物中筛选出3对引物对40481江苏农业学报㊀2024年第40卷第1期份苹果进行检测ꎬ结果表明ꎬ这3对引物可以鉴定区分40份苹果ꎮKimura等[16]用9个SSR标记鉴别60个亚洲梨品种ꎬ研究结果表明其中7个SSR标记能将58个品种区分开ꎮ刘国彬等[17]以19种欧李种质及其近缘种杏李㊁李资源为试验材料ꎬ利用SSR标记进行品种鉴定并构建分子身份证ꎮ魏姗姗等[18]以95份桃品种为试材ꎬ利用18对SSR引物对桃进行遗传多样性分析ꎬ发现了桃品种的8个连锁群ꎮ胡光明等[19]以红阳猕猴桃为材料ꎬ从435对SSR引物中筛选出67对引物ꎬ用于猕猴桃属种质资源的亲缘关系及遗传多样性分析ꎮMahjbi等[20]以20个突尼斯柑橘品种为材料ꎬ通过7个SSR位点建立了它们的亲缘关系来探究其遗传多样性ꎮ此外SSR分子标记也被用于果树品质育种㊁抗性育种以及遗传图谱构建(表3)ꎮ表3㊀简单序列重复(SSR)分子标记在果树上的应用Table3㊀Applicationofsimplesequencerepeat(SSR)molecularmarkersonfruittrees物种(属)㊀㊀㊀功能㊀主要结果参考文献苹果(MalusMill.)品种鉴定用144对SSR引物对40份苹果进行检测ꎬ最少用3对引物进行组合即可鉴定40份苹果[15]筛选品质位点以短枝富士和粉红女士216株F1代群体为试验材料ꎬ使用SSR标记获得了2个与果实酸度连锁的分子标记[21]筛选抗病基因从144对SSR引物中选出17对在抗病和感病之间表现出多态性的引物标记苹果抗褐斑病ꎬ结果表明标记基因位于苹果第8连锁群LG8上[22]梨(PyrusL.)品种鉴定以亚洲梨为材料ꎬ用9个SSR标记鉴别60个品种的亚洲梨能将58个品种区分开[16]李(PrunusL.)品种鉴定以19种欧李种质及其近缘种杏李㊁李资源为试验材料ꎬ利用SSR标记进行品种鉴定并构建分子身份证[17]遗传多样性分析以95份不同的桃品种为研究材料ꎬ用SSR标记对桃遗传多样性进行分析ꎬ可将桃分为扁球形和球形2个类群[18]柑橘(CitrusL.)遗传多样性分析以20个突尼斯柑橘品种为材料ꎬ通过7个SSR位点建立了它们的亲缘关系[20]葡萄(VitisL.)筛选无核基因20个个体被选为无籽葡萄育种研究的遗传资源ꎬVMC7f2标记被鉴定为与无核性状最相关的标记[23]绘制遗传图谱在96个个体的全同胞群体的2个亲本上共测试了346对引物ꎬ成功扩增了310个标记ꎬ基于2个群体中245个SSR标记的分离ꎬ构建了4个图谱[24]筛选抗性基因使用葡萄参考连锁图的SSR标记ꎬ在连锁群15(LG15)上确定抗性位点的位置[25]猕猴桃(ActinidiaLindl.)性别鉴定利用毛花猕猴桃的基因组来开发与性别鉴定相关SSR标记ꎬ从中筛选出了1对可以鉴定毛花猕猴桃雌雄株的引物[26]1.4㊀SRAP分子标记相关序列扩增多态性(Sequencerelatedampli ̄fiedpolymorphismꎬSRAP)标记与RAPD标记的原理和步骤极为相似ꎬ但SRAP相对于RAPDꎬ稳定性高ꎬ重复性好ꎬ多态性较高ꎮ董星光[27]以黄冠和鸭梨为试材ꎬ用SRAP和AFLP标记对其抗病基因进行分析ꎬ发现与抗黑星病相关的1条SRAP标记和1条AFLP标记ꎬ可见SRAP与AFLP可联合用于梨的抗病品种的选育ꎮ冯涛等[28]以小白桃㊁津柳早红为试材ꎬ用12条SRAP引物对其早熟基因进行分析ꎬ结果表明SRAP标记可以用于鉴定桃成熟期芽变(表4)ꎮ表4㊀相关序列扩增多态性(SRAP)分子标记在果树上的应用Table4㊀Applicationofsequencerelatedamplifiedpolymorphism(SRAP)molecularmarkersonfruittrees物种(属)㊀功能㊀主要结果参考文献梨(PyrusL.)筛选抗病基因以黄冠梨和鸭梨为试材ꎬ用SRAP标记对其抗病基因进行分析ꎬ从120对引物中筛选出1条与抗黑星病相关的SRAP标记[27]李(PrunusL.)筛选早熟基因以小白桃㊁津柳早红为试材用12条SRAP引物对其早熟基因进行分析ꎬ结果表明SRAP标记可以鉴定桃成熟期芽变[28]苹果(MalusMill.)筛选耐盐基因以西府海棠和S19杂交组的F1单株为试材ꎬ用SRAP标记对其耐盐基因进行分析ꎬ获得了4条与耐盐基因相关的标记[29]柑橘(CitrusL.)遗传多样性分析以51份酸橙(Citrusaurantium)为研究材料ꎬ用21个SRAP引物来研究其与亲本的遗传关系和多样性[30]葡萄(VitisL.)遗传多样性分析以39个玫瑰香系葡萄品种为试材ꎬ利用SRAP标记技术对其遗传多样性进行分析并构建其DNA指纹图谱[31]椰子(CocosL.)遗传多样性分析使用SRAP标记分析从湄公河三角洲收集的19个椰子品种的遗传多样性ꎬ并绘制其遗传关系[32]581孙雨桐等:果树分子标记辅助育种研究进展1.5㊀SCAR分子标记测序的扩增区段(Sequencecharacterizedampli ̄fiedregionꎬSCAR)标记最初是从RAPD分子标记技术衍生而来的ꎬ并且重复性和特异性比RAPD更好ꎮ为减少农药对环境和人体的危害ꎬ选育抗病品种具有重要意义ꎬSCAR分子标记在果树抗病育种方面发挥着重要的作用ꎮ祁楠等[33]以秦冠和富士F1代群体为试材ꎬ将苹果抗斑点落叶病相关基因的1个RAPD标记转换为SCAR标记(表5)ꎮDeng等[34]以柑橘为材料克隆并测序了20个与抗柑橘三叶草病毒基因连锁的RAPD片段中的7个ꎬ并将它们转化为SCAR标记ꎮ赵伟[35]以用8种葡萄作为亲本的杂交后代为材料ꎬ用SCAR标记SCO11 ̄914对其白粉病抗性进行检测ꎮ表5㊀测序的扩增区段(SCAR)分子标记在果树上的应用Table5㊀Applicationofsequencecharacterizedamplifiedregion(SCAR)molecularmarkersonfruittrees物种(属)㊀功能㊀主要结果参考文献苹果(MalusMill.)筛选抗病基因以秦冠和富士F1代群体为试材ꎬ将苹果抗斑点落叶病基因的1个RAPD标记转换为SCAR标记[33]柑橘(CitrusL.)筛选抗病基因克隆并测序了20个与抗柑橘三叶草病毒基因连锁的RAPD片段中的7个ꎬ并将它们转化SCAR标记[34]梨(PyrusL.)筛选矮化基因以矮化梨与茌梨的杂交后代共111个单株为试材ꎬ获得了1个控制梨树矮化性状基因的RAPD标记S1172 ̄940ꎬ并转换成了SCAR标记[36]葡萄(VitisL.)筛选无核基因利用SCF27 ̄2000和SCC8 ̄1018对32个杂种株系进行分子标记检测ꎬ其中有17个株系都出现了无核的特异性条带[37]筛选抗病基因以8种葡萄作为亲本的杂交后代材料ꎬ用SCAR标记SCO11 ̄914对其白粉病抗性进行检测[35]1.6㊀SNP分子标记SNP检测方法主要有酶切扩增多态性序列(CAPS)㊁单链构象多态性(SSCP)㊁直接测序和基因芯片等ꎮBaldi等[38]通过2个苹果品种Fiesta和Discovery之间杂交的分离群体开发CAPS和SSCP标记ꎬ标记其抗性基因的定位ꎬ实现了克隆序列的遗传作图ꎮSNP多用于遗传图谱的构建ꎮAntan ̄aviciute等[39]使用来自28种海棠基因型的SNP数据为海棠开发了全基因组基因分型阵列ꎮ该阵列为任何给定的海棠后代提供了高通量基因分型和连锁图谱开发的前景ꎮ将2272个SNP标记的数据整合到M432子代的图谱中ꎬ并展示了最完整和最饱和的普米拉分枝杆菌17个连锁群的图谱ꎮ唐海霞等[40]以冬枣和金丝4号的F1代103株群体为材料ꎬ用简化基因组测序技术(GBS)开发的SNP构建了1张包含12条连锁群的遗传图谱ꎮ相关研究(表6)为果树数量性状定位㊁功能基因挖掘以及图位克隆等研究提供了有效的理论依据ꎮ表6㊀单核苷酸多态性(SNP)分子标记在果树上的应用Table6㊀Applicationofsinglenucleotidepolymorphism(SNP)molecularmarkersonfruittrees物种(属)㊀㊀㊀功能㊀主要结果参考文献苹果(MalusMill.)绘制遗传图谱将2272个SNP标记的数据整合到M432子代的图谱中ꎬ并展示了最完整和最饱和的普米拉分枝杆菌17个连锁群的图谱[39]枣(ZiziphusMill.)绘制遗传图谱以冬枣和金丝4号杂交的F1代103株群体为材料ꎬ用SNP分子标记构建了1张包含12条连锁群的遗传图谱[40]柑橘(CitrusL.)品种鉴定利用SNP分子标记区分Haryejosaeng与其他7个温州蜜柑品种的DNA序列差异[41]葡萄(VitisL.)筛选抗虫基因SNP标记可用于葡萄对爪哇根结线虫抗性基因(MJR1)的标记辅助选择[42]龙眼(DimocarpusLour.)绘制遗传图谱以200株凤梨朵和大乌圆的杂交后代F1作为作图群体ꎬ利用RAD ̄seq技术开发SNP分子标记构建遗传图谱[43]2㊀果树遗传图谱构建概况分子标记已被广泛用于构建遗传图谱ꎬ遗传图谱是基于遗传距离的图谱ꎬ它反映的是不同基因座之间的遗传距离和连锁程度ꎮ目前ꎬ大量分子标记如SNP㊁SCAR㊁SSR㊁AFLP㊁SRAP等被用于果树遗传作图ꎮWu等[44]以八月红和砀山酥梨杂交的102个梨F1代单株为作图群体ꎬ用SNP与SSR整合构建681江苏农业学报㊀2024年第40卷第1期了梨的高密度连锁图谱ꎬ该图谱是用快速和稳健的限制性相关DNA测序技术(RADseq)绘制的ꎮ连锁图谱由SNP标记和SSR标记组成ꎬ共3241个标记ꎬ跨度为2243.4cMꎬ平均标记距离为0 70cMꎮ刘更森[45]以富士和金冠杂交的F1代122个单株为作图群体ꎬ选用已开发的SNPꎬ结合SSR标记构建了苹果遗传图谱ꎮ以杧果金黄和贵妃杂交的98株F1代植物为作图群体ꎬ用SRAP㊁AFLP和ISSR分子标记进行作图ꎬ该图谱由33个连锁群组成ꎬ总遗传距离为1561.1cM[46]ꎮ已构建遗传图谱的果树品种见表7ꎮ表7㊀果树遗传图谱构建概况Table7㊀Overviewoffruittreegeneticmappingconstruction物种㊀㊀㊀(属的拉丁名)㊀㊀㊀作图亲本㊀㊀㊀㊀㊀㊀㊀㊀标记类型㊀㊀㊀㊀㊀㊀㊀㊀群体大小(株)连锁群数量(个)参考文献苹果(MalusMill.)富士ˑ坂田津轻SNP15017[47]蜜脆ˑ秦冠SNP35034[48]富士ˑ金冠SNP㊁SSR12217[45]梨(PyrusL.)崇化大梨ˑ新世纪梨SSR㊁SRAP㊁ISSR21014[49]红茄梨ˑ晚秀梨SNP㊁SSR16117[50]八月红ˑ砀山酥梨AFLP㊁SRAP㊁SSR9717[51]李(PrunusL.)黄水蜜ˑ中油桃14号SNP868[52]红垂枝ˑ白花山碧SSR㊁AFLP㊁SRAP5211[53]柑橘(CitrusL.)梨橙2号ˑ晚蜜2SSR㊁COS8010[54]Murcott橘橙ˑPêra甜橙AFLP879[55]Cravo橘ˑPêra甜橙RAPD9412[56]葡萄(VitisL.)赤霞珠ˑ左优红SNP㊁SSR18119[57]枣(ZiziphusMill.)JMS2ˑ邢16SNP16712[58]冬枣ˑ金丝4号SNP㊁SSR11112[59]荔枝(LitchiSonn.)马贵荔ˑ焦核三月红RAPD㊁SRAP㊁AFLP7620[60]芒果(MangiferaL.)金黄ˑ贵妃SRAP㊁AFLP㊁ISSR9833[46]龙眼(DimocarpusLour.)凤梨朵ˑ大乌圆RAPD㊁ISSR㊁SRAP㊁AFLP9421[61]SNP:单核苷酸多态性ꎻSSR:简单序列重复ꎻSRAP:相关序列扩增多态性ꎻISSR:简单重复系列区间ꎻAFLP:扩增片段长度多态性ꎻCOS:保守直系同源序列ꎻRAPD:随机扩增多态性DNAꎮ3㊀QTL基因定位在果树育种中的应用通常没有一种基因能唯一决定某些性状ꎬ一般一组基因作为一个整体控制着某一特性ꎮ与特定数量性状相关的基因所在的基因组区域称为QTLꎬ即数量性状位点ꎮ自从分子标记出现以来ꎬ研究人员和育种人员一直致力于识别与这些QTL相关的功能标记ꎬ在果树的重要性状QTL定位和分子标记辅助育种等方面均取得较好的成果ꎬ如果树生长发育㊁果实的品质㊁抗逆性的强弱等ꎮ前人已对苹果㊁梨㊁柑橘等多种果树的生长发育特性(开花㊁生根能力㊁矮化等)㊁果实品质(质量㊁大小㊁颜色㊁硬度㊁风味等)㊁抗生物胁迫㊁抗非生物胁迫(耐盐碱㊁抗干旱㊁抗寒等)等重要农艺性状进行分析ꎬ有助于培育特定目标性状的果树品种ꎮ3.1㊀与苹果相关的QTL苹果QTL的鉴定集中于其主要农艺性状[如矮化㊁果实品质(包括果实质量㊁果实大小㊁果实颜色以及果肉的糖酸含量)㊁苹果的抗逆性(抗寒㊁抗旱㊁耐盐碱等)]ꎮFoster等[62]对41份M93Robusta5(非矮化)与Braeburn接穗嫁接的砧木群体的QTL进行分析ꎬ结果表明ꎬ连锁群LG5上的一个主要QTL对接穗的矮化有显著影响ꎮZheng等[63]用来自海棠㊁红富士㊁金冠㊁乔纳森家系9422株苹果F1代杂交种为试材ꎬ检测得到9个与苹果果皮颜色遗传变异有关的微效QTL(表8)ꎮ孙瑞[64]以红玉㊁金冠以及红781孙雨桐等:果树分子标记辅助育种研究进展玉和金冠的F1代杂交群体297株苹果为试材ꎬ对其品质性状进行QTL定位ꎬ共得到12个QTL位点ꎮ3.2㊀与梨相关的QTL迄今ꎬQTL定位在梨的研究上取得了一定的成果ꎮ赵亚楠[65]利用苹果梨和八月红150株F1代材料对14个果实品质性状进行基因定位分析ꎬ共获得28个QTL位点ꎬ其中与单果质量相关的QTL位点有5个㊁与果心大小相关的QTL位点有2个㊁与果肉硬度相关的QTL位点有3个㊁与果实横径相关的QTL位点有6个㊁与果梗长度相关的QTL位点有1个㊁与可溶性固形物含量相关的QTL位点有6个㊁与可滴定酸含量相关的QTL位点有2个ꎬ共扫描到2266个基因ꎮSun等[66]以131个亚洲梨和欧洲梨为试验材料ꎬ在已被鉴定的病害相关QTL区域中找到41个核苷酸结合位点(NBS)编码基因(表9)ꎮ表8㊀苹果相关的数量性状座位(QTL)Table8㊀Quantitativetraitlocus(QTL)relatedtoapple亲本功能㊀㊀㊀群体大小(株)QTL数量(个)参考文献Malling9ˑRobus ̄ta5标记矮化的位点416[62]海棠㊁红富士㊁金冠㊁乔纳森标记果皮颜色的位点94229[63]红玉ˑ金冠标记果实质量的位点2972[64]红玉ˑ金冠标记果实酸度的位点2976[64]红玉ˑ金冠标记糖含量的位点2972[64]红玉ˑ金冠标记抗病的位点53429[67]BalengCrabˑM9标记耐盐碱的位点325842[68]MalusdomesticaˑM.asiatica标记果肉硬度的位点266432[69]MalusdomesticaˑM.asiatica标记果肉脆度的位点266430[69]FortuneˑMurcott标记苹果酸的位点14638[70]秦冠ˑ蜜脆标记抗旱的位点35055[71]3.3㊀与柑橘相关的QTL通过分子标记构建柑橘的遗传连锁图谱可以获得果实发育等农艺性状和应答逆境胁迫的QTLꎮ罗艾等[72]以晚蜜2号和梨橙2号的94株F1代材料为群体ꎬ进行QTL定位分析ꎬ发现4个与果实质量相关的QTL定位ꎬ7个与果实大小相关的QTL定位ꎮ马喜军[73]以晚蜜2号和梨橙2号的350株F1代为试验材料对柑橘抗寒性相关的QTL进行定位ꎬ得到7个与柑橘抗寒性相关的QTLꎬ分布于4个连锁群上ꎬ分别为LW1㊁LW2㊁LW3和LW8ꎮHuang等[74]对甜橙ˑ枳橙属间杂交的170株F1代进行基因分型分析ꎬ在枳遗传图谱上鉴定到4个与柑橘黄龙病相关的QTL(表10)ꎮ表9㊀梨相关的数量性状座位(QTL)Table9㊀Quantitativetraitlocus(QTL)relatedtopear亲本㊀㊀功能㊀㊀㊀群体大小(株)QTL数量(个)参考文献苹果梨ˑ八月红标记果实大小的位点1506[65]苹果梨ˑ八月红标记果肉硬度的位点1503[65]苹果梨ˑ八月红标记可溶性固形物的位点1506[65]苹果梨ˑ八月红标记可滴定酸的位点1502[65]苹果梨ˑ八月红标记果实质量的位点1505[65]亚洲梨ˑ欧洲梨标记抗病的位点13141[66]八月红ˑ砀山酥梨标记果实质量的位点1022[75]八月红ˑ砀山酥梨标记果实大小的位点1022[75]表10㊀柑橘相关的数量性状座位(QTL)Table10㊀Quantitativetraitlocus(QTL)relatedtocitrus亲本㊀㊀功能㊀㊀㊀群体大小(株)QTL数量(个)参考文献晚蜜2号ˑ梨橙2号标记果实质量的位点944[72]晚蜜2号ˑ梨橙2号标记果实大小的位点947[72]梨橙2号ˑ晚蜜2号标记抗寒的位点3507[73]甜橙ˑ枳橙标记抗病的位点1704[74]红橘ˑ枳壳标记果肉色泽的位点792[76]FortuneˑMurcott标记挥发性物质的位点116206[77]TrifoliataˑCleopatramandarin标记耐盐碱的位点98[78]3.4㊀与其他果树相关的QTL除常见果树外ꎬ其他果树重要性状相关的QTL定位研究也取得了很大进展ꎮCirilli等[79]评估133份桃种质ꎬ定位到1个可以推迟桃花期的QTLꎮ鲍荆凯[80]用JMS2和交城5号枣的150株F1代材料对其QTL定位进行分析ꎬ共获得104个QTL位点ꎬ其中与果实大小相关的QTL位点57个㊁与果实糖组分相关的QTL位点17个㊁与果实酸组分相关的QTL位点30个ꎮ刘春燕[81]以桂海4号和山梨猕猴桃的杂交后代开展QTL定位研究ꎬ共检测到44个QTL位点ꎬ其中与果实质量相关的QTL位点7个ꎬ与果实横纵径相关的QTL位点21个ꎮ史晓畅[82]以881江苏农业学报㊀2024年第40卷第1期山东大绵球和新宾软籽山楂的130株杂交F1代为材料ꎬ检测到2个与单果质量相关的QTL位点ꎬ6个与果皮穿刺硬度相关的QTL位点ꎬ3个与果实大小相关的QTL位点ꎬ5个与果皮脆性相关的QTL位点ꎬ5个与果肉平均硬度相关的QTL位点ꎮ这些研究(表11)不仅丰富了果树的遗传学研究ꎬ也为果实品质及果树抗逆的遗传机制和育种研究提供了理论基础ꎮ4㊀展望在果树的品种鉴定㊁辅助育种(早熟㊁无核㊁矮化㊁品质以及抗性等)㊁遗传图谱的构建和农艺性状基因定位等方面ꎬDNA分子标记被广泛应用ꎮDNA分子标记技术种类多样ꎬ大致可分为三类ꎮ第一类:以电泳技术和分子杂交技术为核心的分子标记技术ꎬ如RFLPꎻ第二类:以DNA聚合酶链式反应(PolymerasechainreactionꎬPCR)为基础的分子标记技术ꎬ包括RAPD㊁SSR㊁SCAR等ꎻ第三类:以DNA测序为核心的分子标记技术ꎬ如SNP标记[83]ꎮRFLP具有共显性且不需要先验序列信息ꎬ但它耗时长ꎬ需要大量纯DNAꎬ价格也比较昂贵[84]ꎮRAPD操作简单ꎬ所需DNA量少ꎬ多态显性ꎬ但需要高度纯化的DNA并且再现性低ꎮDNA的数量和质量㊁PCR缓冲液㊁氯化镁浓度㊁退火温度和TaqDNA聚合酶是影响RAPD标记再现性的一些重要因素[85]ꎮAFLP标记将RFLP和PCR技术结合在一起ꎬ先对DNA进行消化ꎬ然后进行PCRꎮAFLP标记物具有低成本ꎬ并且不需要先前的序列信息ꎮ在AFLP中ꎬ既可以使用高质量的DNAꎬ也可以使用部分降解的DNAꎬ但是ꎬ该DNA不能含有任何限制性内切酶或PCR抑制剂[86]ꎮ相较于RFLP㊁RAPD㊁AFLP等分子标记ꎬSSR标记具有多态性检出率高㊁基因组中分布广泛㊁结果稳定可靠等特点ꎬ是检测品种真实性㊁分析品种间遗传差异以及鉴定纯度的理想标记[87]ꎮSNP可以提供最简单和最大数量的标记ꎮSNP在植物和动物中大量存在[88 ̄91]ꎬ植物中的SNP频率为每100~300bp中有1个SNPꎮSNP成本低ꎬ在基因组中广泛分布ꎬ无需先验序列信息ꎬ再现性高ꎬ共显性标记ꎬ但其开发成本较高ꎮ人们基于不同的等位基因识别技术和检测平台已经开发了大量的SNP基因分型方法ꎬ其中ꎬRLFP(SNP ̄RFLP)是最简单的方法ꎬCAPS标记技术也可以应用于SNP检测[92]ꎮ分子标记种类繁多ꎬ功能优势也有所不同ꎬ根据试验的目的选择合适的分子标记有助于解决具体问题ꎮ表11㊀与其他果树相关的数量性状座位(QTL)Table11㊀Quantitativetraitlocus(QTL)relatedtootherfruittrees物种(属)㊀㊀㊀㊀㊀㊀亲本㊀㊀㊀㊀㊀功能㊀㊀㊀㊀㊀㊀㊀群体大小(株)QTL数量(个)参考文献李(PrunusL.)标记开花的位点1331[79]枣(ZiziphusMill.)JMS2ˑ交城5号标记果实大小的位点15057[80]JMS2ˑ交城5号标记果实糖组分的位点15017[80]JMS2ˑ交城5号标记果实酸组分的位点15030[80]猕猴桃(ActinidiaLindl.)桂海4号ˑ山梨猕猴桃标记果实质量的位点1747[81]桂海4号ˑ山梨猕猴桃标记果实大小的位点17421[81]山楂(CrataegusL.)山东大绵球ˑ新宾软籽标记果实质量的位点1302[82]山东大绵球ˑ新宾软籽标记果实大小的位点1303[82]山东大绵球ˑ新宾软籽标记果皮硬度的位点1306[82]山东大绵球ˑ新宾软籽标记果肉脆度的位点1305[82]葡萄(VitisL.)赤霞珠ˑ左优红标记抗寒的位点1818[93]枣(ZiziphusMill.)DavisˑGeorgiaBelle标记抗寒的位点21112[94]㊀㊀为同时提高育种效率㊁缩短育种周期ꎬ寻找与农艺性状密切相关的分子标记至关重要ꎮ分子标记辅助育种(MAS)的优势可以体现在以下3个方面ꎮ一㊁允许提前选择ꎮ在育种中ꎬ有些性状需要特定的生长环境和一定的生长周期ꎮ二㊁同一性状利用多个等位基因ꎮ在不同的育种材料中ꎬ可能存在多个基因影响同一性状(如抗病性和品质)ꎬ利用表型很难识别这些等位基因ꎮ三㊁允许同时选择多个性状ꎮ所选单株或品系不仅要在单株抗病㊁品质㊁产量等方面表现良好ꎬ综合性状也要相对较好ꎮ因此ꎬ有必要对育种的种群981孙雨桐等:果树分子标记辅助育种研究进展中每个目标性状逐一进行识别和筛选ꎮ以往的研究大多将目的基因的分子标记与育种工作分离ꎬ不能很好地应用于实际ꎮ今后分子标记技术的发展将与传统育种相结合ꎬ使其尽快为育种工作服务ꎮ这有助于提高果树作物育种效率ꎬ加快育种发展进程ꎮ参考文献:[1]㊀KATULA ̄DEBRECENIDꎬLENCSESAKꎬSZOKEAꎬetal.Marker ̄assistedselec ̄tionfortwodominantpowderymildewresistancegenesintrogressedintoahybridgrapepopulation[J].ScientiaHoriculturaeꎬ2010ꎬ126(4):448 ̄453.[2]㊀TARTARINIS.RAPDmarkerslinkedtotheVfgeneforscabre ̄sistanceinapple[J].TheoreticalandAppliedGeneticsꎬ1996ꎬ92:803 ̄810.[3]㊀杨亚州ꎬ王跃进ꎬ张剑侠ꎬ等.中国葡萄属野生种抗旱基因的分子标记及遗传分析[J].园艺学报ꎬ2007ꎬ34(5):1087 ̄1092. [4]㊀余智城ꎬ何雪娇ꎬ林秀香ꎬ等.琯溪蜜柚芽变种质遗传多样性的RAPD分析[J].福建热作科技ꎬ2022ꎬ47(3):38 ̄41. [5]㊀RAJAPAKSESꎬBELTHOFFLEꎬHEGꎬetal.GeneticlinkagemappinginpeachusingmorphologicalꎬRFLPandRAPDmarkers[J].TheoreticalandAppliedGeneticsꎬ1995ꎬ90(3/4):503 ̄510. [6]㊀WARBURTONMLꎬBECERRA ̄VELáSQUEZVLꎬGOFFREDAJCꎬetal.UtilityofRAPDmarkersinidentifyinggeneticlinkagestogenesofeconomicinterestinpeach[J].TheoreticalandAppliedGeneticsꎬ1996ꎬ93(5/6):920 ̄925.[7]㊀黄晗达ꎬ杨静慧ꎬ龚无缺ꎬ等.7个樱桃品种亲缘关系的RAPD分析[J].天津农学院学报ꎬ2018ꎬ25(1):5 ̄8.[8]㊀HIMABINDUAꎬRAJASEKHARM.CharacterizationoftraditionalmangogermplasmofcoastalAndhraPradeshusingRAPDmarkers[J].ElectronicJournalofPlantBreedingꎬ2021ꎬ12(4):1261 ̄1267.[9]㊀董美超ꎬ杨㊀帆ꎬ李进学ꎬ等.90份鳄梨种质资源AFLP遗传多样性分析[J].福建农业学报ꎬ2020ꎬ35(1):13 ̄19.[10]LAICWꎬLINYLꎬZHOUXJꎬetal.AFLPandMSAPanalysisof LaneLate navelorangeanditsbudsportpumpkin ̄likenavelorange[J].PlantDiseasesandPestsꎬ2022ꎬ13(1):20 ̄25. [11]王晓英ꎬ郭廷松ꎬ王新花ꎬ等.4个苹果品种的AFLP分子标记研究[J].山东农业大学学报(自然科学版)ꎬ2018ꎬ49(1):90 ̄93.[12]王㊀平ꎬ唐小浪ꎬ马培恰ꎬ等.辐射诱变和芽变柑橘品种(系)的AFLP分析[J].果树学报ꎬ2012ꎬ29(1):130 ̄134.[13]赖春旺ꎬ周小娟ꎬ米兰芳ꎬ等.脐橙早熟芽变及其早熟性状回复型材料的AFLP和MSAP分析[J].果树学报ꎬ2022ꎬ39(8):1346 ̄1357.[14]张㊀慧ꎬ张世鑫ꎬ吴绍华ꎬ等.猕猴桃属33份种质资源的AFLP遗传多样性分析[J].生物学杂志ꎬ2018ꎬ35(2):29 ̄33. [15]王立新ꎬ张小军ꎬ史星雲ꎬ等.苹果栽培品种SSR指纹图谱的构建[J].果树学报ꎬ2012ꎬ29(6):971 ̄977.[16]KIMURATꎬSHIYZꎬSHODAMꎬetal.Identificationofasian ̄pearvarietiesbySSRanalysis[J].BreedingScienceꎬ2002ꎬ52(2):115 ̄121.[17]刘国彬ꎬ姚砚武ꎬ曹㊀均.利用荧光SSR标记构建欧李种质分子身份证[J].东北林业大学学报ꎬ2022ꎬ50(10):10 ̄17. [18]魏姗姗ꎬ杨敏生ꎬ梁海永.桃品种遗传多样性SSR分析[J].耕作与栽培ꎬ2022ꎬ42(1):1 ̄5ꎬ9.[19]胡光明ꎬ张㊀琼ꎬ韩㊀飞ꎬ等.猕猴桃属植物通用型SSR分子标记引物的筛选及应用[J].中国农业科学ꎬ2022ꎬ55(17):3411 ̄3425.[20]MAHJBIAꎬOUESLATIAꎬBARAKETGꎬetal.AssessmentofgeneticdiversityofTunisianorangeꎬCitrussinensis(L.)osbeckusingmicrosatellite(SSR)markers[J].GeneticsandMolecularResearchꎬ2016ꎬ15(2):1 ̄12.[21]王雷存ꎬ樊红科ꎬ高㊀华ꎬ等.苹果酸度基因(Ma)SSR标记及遗传分析[J].园艺学报ꎬ2012ꎬ39(10):1885 ̄1892.[22]寿园园.苹果抗褐斑病性遗传分析与SSR分子标记[D].哈尔滨:东北农业大学ꎬ2009.[23]AKKURTMꎬÇAKIRAꎬSHIDFARMꎬetal.UsingSCC8ꎬSCF27andVMC7f2markersingrapevinebreedingforseedlessnessviamarkerassistedselection[J].GeneticsandMolecularRe ̄searchꎬ2012ꎬ11(3):2288 ̄2294.[24]ADAM ̄BLONDONAFꎬROUXCꎬCLAUXDꎬetal.Mapping245SSRmarkersontheVitisviniferagenome:atoolforgrapege ̄netics[J].TheoretischeandAngewandteGenetikꎬ2004ꎬ109(5):1017 ̄2227.[25]KUCZMOGAꎬGALAMBOSAꎬHORVáTHSꎬetal.MappingofcrowngallresistancelocusRcg1ingrapevine[J].TheoretischeandAngewandteGenetikꎬ2012ꎬ125(7):1565 ̄1574.[26]刘嘉艺ꎬ岳俊阳ꎬ刘永胜.基于毛花猕猴桃基因组的性别相关SSR分子标记的开发[J].合肥工业大学学报(自然科学版)ꎬ2022ꎬ45(8):1135 ̄1138ꎬ1146.[27]董星光.梨抗黑星病基因的分子标记研究[D].北京:中国农业科学院ꎬ2009.[28]冯㊀涛ꎬ刘㊀娟ꎬ华夏雪.利用SSR㊁SRAP分子标记鉴定桃早熟芽变[J].江苏农业科学ꎬ2017ꎬ45(6):42 ̄44.[29]孙叶红ꎬ张㊀媛ꎬ李中勇ꎬ等.苹果砧木耐盐性基因SRAP标记的鉴定及序列分析[J].华北农学报ꎬ2015ꎬ30(2):59 ̄63. [30]POLATIꎬKACARYAꎬYESILOGLUTꎬetal.Molecularcharac ̄terizationofsourorange(Citrusaurantium)accessionsandtheirrelativesusingSSRandSRAPmarkers[J].GeneticsandMolecu ̄larResearchꎬ2012ꎬ11(3):3267 ̄3276.[31]尚晓星ꎬ张安世ꎬ刘㊀莹ꎬ等.玫瑰香系葡萄种质资源SRAP遗传多样性分析及指纹图谱构建[J].分子植物育种ꎬ2020ꎬ18(6):1916 ̄1922.[32]XUANDTKꎬNGUYENQTꎬKHANGNHMꎬetal.Molecularcharacterizationofcoconut(CocosnuciferaL.)varietiesinViet ̄namusingsequence ̄relatedamplifiedpolymorphism(SRAP)markers[J].Biologiaꎬ2022ꎬ77(11):183 ̄191.[33]祁㊀楠ꎬ万怡震ꎬ高㊀华ꎬ等.苹果抗斑点落叶病基因的一个091江苏农业学报㊀2024年第40卷第1期。

线粒体DNA分子标记在动物种群遗传学研究中的应用随着人们对生物学,特别是遗传学的研究越加深入,越来越多的科学家开始关注动物种群遗传学,这涉及到基因在种群中的分布和演化。
而在这个领域中,线粒体DNA分子标记的应用变得越来越普遍和重要。
一、什么是线粒体DNA?线粒体是生物细胞中可以自主繁殖的细胞器。
除了有自主繁殖的能力,线粒体还有着不同于其他细胞器的基因,这些基因以环形的方式存在于线粒体DNA中。
线粒体DNA只存在于母亲的卵细胞内,而不会从父亲的精子中传递给下一代,这种遗传方式也被称为无性遗传。
因此,线粒体DNA可以被用来研究动物种群演化中母亲线ages的演化历程。
二、线粒体DNA的分析方法线粒体DNA的研究方法主要有两种:限制性片段长度多态性(RFLP)研究和DNA条形码。
这两种方法在使用上存在一些不同。
RFLP研究的方法主要是通过测量线粒体DNA的长度和序列变异,来区分同种或不同种之间线粒体DNA的差异。
这种方法通过对目标基因进行摄影并用化学荧光或其他技术处理,可以得到独特的串段图。
RFLP方法的主要优势在于可以半定量地测量线粒体DNA的序列变异,并对不同的基因型进行分类和定位。
同时,使用这种方法还可以识别出潜在的基因漏洞和致病突变。
DNA条形码方法则是使用PCR(聚合酶链式反应)将线粒体DNA从样本中扩增并鉴定序列,再将其转化为一种特定的AluI酶切位点P1和P2之间的短序列。
通过比较这些序列相似性来比较不同种物种,在这种技术下,样本的数量可以多达数百万份,而处理和分析每份样本也非常快捷并且高效。
三、线粒体DNA在动物种群遗传学中的应用通过分析不同动物种内的线粒体DNA差异,可以揭示它们在已知的地质历史和事件中是如何相互交流和进化的。
动物的地理分布和分化是由于基因演化的结果,而线粒体DNA对于识别物种的分化和地理分布具有高度传递性。
下面,就来谈一谈线粒体DNA在不同环境下的应用。
1.谱系分析通过分析线粒体DNA,可以确定某一动物物种的谱系,比如说,白头海鹰、斑点猫、印度狮、非洲象都是由多个谱系组成的。


1132017年34卷第02期 SWINE INDUSTRY SCIENCE 猪业科学遗传改良GENETIC IMPROVEMENT精品思想 市场战略分子遗传标记技术及其在动物育种中的研究进展宋志芳1,于国生1,,解佑志1,芦春莲1,2,曹洪战1,2*(1.河北农业大学动物科技学院,河北 保定 071000;2.河北农业大学猪业科学研究所,河北 保定 071000)摘 要:遗传标记经历了从传统的标记即形态学标记、细胞学标记、生物化学标记到现代分子标记的发展,分子标记具有很多优势,也促进了动植物育种、人类医学、基因定位以及构建遗传图谱的改革。
遗传标记能应用于畜禽的遗传多样性分析、种质资源的鉴定、亲缘关系的研究、遗传图谱的构建、分子标记辅助选择和QTL 定位等领域,文章主要综述了分子标记在标记辅助选择的应用。
关键词:标记辅助选择;分子育种;分子标记作者简介:宋志芳(1992-),女,山东菏泽人,研究生,研究方向:动物遗传育种,E-mail :187********@1 分子标记分子标记作为一种遗传标记,以个体间核苷酸序列的变异为基础,能够直接反映出DNA 水平的遗传多态性,有广义分子标记和狭义分子标记之分。
广义的分子标记一般指DNA 序列或蛋白质,能够遗传且可检测;狭义的分子标记一般指特异性DNA 片段,能够反映生物个体或种群间基因组中的差异。
理想的分子标记必须达到以下几个要求:具有高多态性;共显性遗传(即利用分子标记可鉴别二倍体中的基因型);能明确辨别等位基因;遍布整个基因组;要求分子标记在整个基因组中分布均匀;即无基因多效性;检测手段简单、快速;成本低;重复性好。
但是在实际实验过程中,分子标记很难达到理想状态。
随着生物技术的发展,目前已经出现了种类不同的分子标记,比如限制性片段长度多态性、小卫星序列、微卫星序列或简单重复序列、随机扩增多态性DNA、扩增片段长度多态性、特定序列位点、DNA 单链构象多态性、单核苷酸多态性以及脉冲场电泳等。

杂种绵羊繁殖与生长性状的DNA分子标记研究的开题报告一、研究背景及意义绵羊作为我国重要的畜禽之一,在养殖业发展中具有不可忽视的地位。
随着农业科技的发展,人们不仅重视绵羊的育种和繁殖性能,而且也开始关注其遗传特征,希望通过DNA分子标记研究,探究绵羊繁殖和生长性状的遗传基础,进而提高绵羊产量和质量。
二、研究现状和存在问题目前,国内外学者对杂种绵羊的繁殖和生长遗传特征的研究取得了一些成果。
国内外学者通过对绵羊微卫星DNA分子标记研究,发现了许多绵羊群体的遗传多样性、亲缘关系和基因多态性等问题。
此外,还有学者通过对绵羊生长发育基因的研究,揭示了肌肉生长、繁殖性能和产奶量等性状的遗传基础。
但是,目前国内对杂种绵羊的DNA分子标记研究还不充分,而且当前绵羊基因组测序和芯片技术还不够成熟,需要进一步探讨和研究。
三、研究目的和内容本研究旨在通过PCR和电泳技术,筛选适宜研究杂种绵羊繁殖和生长性状的微卫星DNA分子标记,并通过对标记位点的检测和测序,探究其在遗传多样性、亲缘关系和基因多态性等方面的特征,进而探究绵羊繁殖和生长性状的遗传基础。
具体内容包括:1.收集杂种绵羊标本样本。
2.筛选适宜研究的微卫星DNA分子标记,设计PCR引物。
3.实验室条件下进行PCR扩增和电泳分析。
4.对标记位点进行检测和测序,分析群体的遗传多样性和亲缘关系。
5.通过群体表型和基因型数据的分析,揭示绵羊繁殖和生长性状的遗传基础实际。
四、研究方案1.选取50只杂种绵羊样本,收集血样进行DNA提取。
2.通过绵羊基因组数据库和前期文献调研,筛选出适宜研究的微卫星DNA分子标记,设计合适的PCR引物。
将所有PCR制品混合后,电泳检测PCR扩增产物。
3.对于标记位点不同的分离纯化的PCR扩增产物,采用ABI3730再生技术进行测序,并进行序列分析和比对。
4.通过测序和分析,文章对样本进行遗传多样性和亲缘关系分析。
利用SPSS软件对群体表型和基因型数据进行统计分析,揭示绵羊繁殖和生长性状的遗传基础。



dna分子标记技术在甘薯遗传育种研究中的应用DNA分子标记技术在甘薯遗传育种研究中的应用一、DNA分子标记技术的介绍DNA分子标记技术是一种在分子水平上识别和区分生物个体之间差异的技术。
它可以帮助科研人员更深入地了解生物的遗传变异和遗传演化,对于植物育种研究尤为重要。
DNA分子标记技术主要包括随机扩增多态性DNA标记 (RAPD)、简单重复序列(SSR)标记、单核苷酸多态性(SNP)标记等。
这些技术具有快速、准确和高效的特点,为甘薯遗传育种研究提供了重要的分子工具。
二、甘薯遗传育种研究中DNA分子标记技术的应用在甘薯遗传育种研究中,DNA分子标记技术被广泛应用于种质资源鉴定、遗传多样性分析、基因定位和分子标记辅助选择等方面。
种质资源是育种研究的重要基础,通过DNA分子标记技术可以准确地鉴定甘薯品种之间的遗传差异,为育种新品种提供了更多选择。
利用DNA分子标记技术进行遗传多样性分析,可以全面了解甘薯种质资源的遗传多样性和遗传结构,为合理利用种质资源提供了科学依据。
DNA分子标记技术还可以帮助科研人员在甘薯中定位关键性状的基因,探索其遗传规律和分子机制。
通过基因定位,可以加速甘薯育种过程,提高选择效率和育种精度。
更重要的是,DNA分子标记技术的应用还可以实现分子标记辅助选择,即在育种材料中直接以分子标记来选择目标性状,大大加快了育种进程。
三、个人观点和理解从我个人的角度来看,DNA分子标记技术在甘薯遗传育种研究中的应用极大地丰富和拓展了育种研究的手段和方法。
它提供了更加准确、快速和高效的分子工具,为甘薯新品种的培育提供了科学依据和技术支持。
这些技术的广泛应用也为甘薯遗传育种研究注入了新的活力和动力,加快了育种进程,促进了甘薯产业的发展和壮大。
四、总结回顾DNA分子标记技术在甘薯遗传育种研究中的应用,极大地促进了甘薯育种研究的深度和广度。
它为甘薯遗传多样性的鉴定、关键性状的基因定位和分子标记辅助选择等方面提供了重要支持,成为育种研究的得力助手。
第23卷第2期周口师范学院学报2006年3月V o.l23No.2Journa l o f Zhoukou No r m alUn iversity M ar.2006分子标记的发展及应用马克世,胡炳义(周口师范学院生命科学系,河南周口466000)摘要:随着分子生物学的迅速发展,DNA分子标记越来越受到重视.本文介绍了分子标记的发展,论述了DNA分子标记在基因组作图、基因克隆、物种亲缘关系分析及疾病诊断等方面的应用.关键词:DNA;多态性;分子标记中图分类号:Q523文献标识码: A 文章编号:16719476(2006)020081-04DNA分子标记是以生物大分子的多态性为基础的遗传标记,是DNA水平遗传变异的直接反映,克服了传统的形态标记、细胞学标记和生化标记易受外界因素、生物个体发育阶段及器官组织差异的影响,因而具有广泛的应用前景.随着分子生物学技术的发展,DNA分子标记的研究愈加活跃,相继有数10种不同的分子标记技术问世.1 分子标记的发展1.1 第一代分子标记限制性片段长度多态性(restr i ction fragm ent leng t h po ly m orphis m s,RFLP)是最早应用的第一代分子标记技术[1],是检测DNA在限制性内切酶酶切后形成的特定DNA片段的大小的方法,反映了DNA分子上不同酶切位点的分布情况.因此, DNA序列上的微小变化,甚至1个核苷酸的变化,也能引起限制性内切酶切点的丢失或产生,导致酶切片段长度的变化. R FLP标记的等位基因具有共显性的特点,结果稳定可靠,重复性好,特别适用于构建遗传连锁图.但是,在进行RFLP分析时,需要对该位点的DNA片段做探针,通常用放射性同位素及核酸杂交技术,这样既不安全又不易自动化.另外,RFLP对DNA多态性检出的灵敏度也不高,RFLP连锁图上还有很多大的空间区(G aps).为了克服RFLP技术上的缺点,W illia m s等人于1990年建立了随机扩增多态性DNA(R andom Amp lified Po ly m orphic DNA s,RAPD)技术[2],由于其独特的检测DNA多态性的方式使得RA PD技术很快渗透于基因研究的各个领域.RA PD是建立在PCR基础之上的,用一个随机引物(8~10个碱基)非定点地扩增DNA片段,然后用凝胶电泳分离扩增片段来进行DNA多态性研究.对任一特定引物而言,它在基因组DNA序列上有其特定结合位点,一旦基因组在这些区域发生DNA片段插入、缺失或碱基突变,就可能导致这些特定结合位点的分布发生变化,从而导致扩增产物的数量和大小发生改变,表现出多态性.与RFLP相比,RA PD方便易行,DNA用量少,设备要求简单,不需DNA探针,设计引物也不需要预先克隆标记或进行序列分析,不依赖于种属特异性和基因组的结构;合成一套引物可以用于不同生物基因组分析,用一个引物就可扩增出许多片段,并且不需要同位素,安全性好.但RA PD技术中影响因素很多,实验的稳定性和重复性差[3].扩增片段长度多态性(Amp lifi ed F ragment L ength Po ly m orphis m s,A FLP)是1992年由荷兰K eygene公司科学家Zabeau和V os发明的一种DNA分子标记新技术[4].其基本原理是基因组DNA经限制性内切酶双酶切后,形成分子量大小不等的随机限制性片段,将特定的接头(adap ter)连接在这些DNA片段的两端,形成一个带接头的特异片段,通过接头序列和PCR引物3 末端的识别,特异性片段经变性、退火和延伸周期性循环而扩增,最终通过聚丙烯酰胺电泳分子筛作用,将这些特异的限制性片段分离开来.AFLP融合了RFLP和RAPD两种技术的优点,既具有RFLP的可靠性,又具有RA PD的灵敏性.它的出现是分子标记技术又一重大突破,被认为是目前一种较为理想、有效的分子标记.此外,在RA PD和RFLP技术基础上建立了SCAR(Sequence character i zed a m plifi ed reg i ons,序列特异性扩增区域), CPA S(C l eaved amp lifiedpo l ymo rph ic sequence,酶切扩增多态序列)和DAF(DNA a mp lified fi ngerpr i nts,DNA扩增指纹)等标记技术[3,5].这些技术的出现,进一步丰富和完善了第一代分子标记,增加了DNA多态性的研究手段.收稿日期:20051228;修回日期:20060216作者简介:马克世(1974),男,河南项城人,硕士,讲师,研究方向:动物生物化学.E m a i:l m adg2000@.82 周口师范学院学报2006年3月1.2 第二代分子标记第二代分子标记利用存在于真核生物基因组中的大量重复序列,如重复单位长度在15~65个核苷酸左右的小卫星DNA(m i nisa tellite DNA),重复单位长度在2~6个核苷酸左右的微卫星DNA(m icrosa tellite DNA),后者又称为简短重复序列(sho rt tande m repeat po l ymo rph is m缩写为STR,STR P,或者si m ple sequence l eng th po ly m orphis m缩写为SSLP)[1].小卫星和微卫星DNA分布于整个基因组的不同位点.由于重复单位的大小和序列不同以及拷贝数不同,从而构成了长度多态性.微卫星多态性有助于遗传图谱的建立,为比较基因组分析奠定了基础[6].微卫星特点[7]:(1)保守性.微卫星标记在哺乳动物种间,特别是关系亲缘较近的物种间具有一定程度的保守性.(2)共显性遗传.微卫星标记的等位基因呈孟德尔共显性遗传,可以区别纯合显性和杂合显性个体.(3)多态信息量大.微卫星DNA在基因组中分布随机,几乎遍布整个基因组,可获得比RFLP更多的多态性.(4)检测容易,省时,重复性较好.在基因组多态性分析中,可采用VNTR(V ar i ble number o f tan de m repea ts,数目可变的串联重复多态性)标记技术区别这些小卫星或微卫星DNA.VNTR基本原理与RFLP大致相同,只是所用的探针核苷酸序列必须是小卫星或微卫星序列,经分子杂交和放射自显影后,即可获得反映个体特异性的DNA指纹图谱.VNTR同RFLP标记一样,实验操作程序繁杂,检测时间长,成本较高.M oo re等于1991年结合PCR技术创立了SSR(S i m p le Sequence R epeat,简单重复序列).SSR即微卫星DNA,是一类由几个(多为1~5个)碱基组成的基序(mo tif)串联重复而成的DNA序列,其长度一般较短,广泛分布于基因组的不同位置,如(CA)n,(A T)n,(GGC)n等重复.不同遗传材料重复次数的可变性,导致了SSR长度的高度变异性,这一变异性正是SSR 标记产生的基础.SSR标记的基本原理:根据微卫星重复序列两端的特定短序列设计引物,通过PCR反应扩增微卫星片段.由于重复的长度及数目变化极大,所以这是检测多态性的一种有效方法.在SSR技术中,采用PCR反应必须知道扩增DNA片段两侧序列,在大多数情况下,某些序列本身或其旁侧序列并不清楚,这就限制了SSR技术的应用.1994年,Z i e t k ie w i cz等对SS R技术进行了发展,建立了加锚微卫星寡核苷酸(A ncho red m i crosatellite o ligonucleoti des)技术[5,8].他们用加锚定的微卫星寡核苷酸作引物,即在SS R的5 端或3 端加上2~4个随机选择的核苷酸,这可引起特定位点退火.这样就能导致与锚定引物互补的间隔不太大的重复序列间的基因组片段进行PCR扩增.这类标记又被称为简单重复序列间扩增ISSR(i nter s i m p l e sequence repeat),锚定简单重复序列扩增A SSR(anchored si m p l e sequence repeats).在所用的两端引物中,一个可以是锚定引物,另一个是随机引物.1.3 第三代分子标记单核苷酸多态性(si ng l e nuc leo ti de po l ymo rph is m,SNP)标记被称为第三代DNA分子标记.这种分子标记是分散于基因组中的单个碱基的差异.这种差异包括单个碱基的缺失和插入,但更常见的是单个核苷酸的替换[9,10].SNP的分布密集,在人类基因组中被认为有300万个以上的SNP遗传标记,这可能达到了人类基因组多态位点数目的极限,因此被认为是应用前景较好的遗传标记物.其优点是:(1)SNP在种群中是二等位基因性的,在任何种群中其等位基因频率都可估计出来;(2)位点丰富,S N P几乎遍布于整个基因组;(3)部分位于基因内部的S N P可能会直接影响蛋白质的结构或基因表达水平,因此它们本身可能就是疾病遗传机制的候选改变位点;(4)遗传稳定性高;(5)易于进行自动化分析[11].目前,已有2000多个标记定位于人类染色体上.S N P与第一代的RFLP及第二代的STR标记的不同之处有两个方面:一是它不再以DNA片段的长度变化作为检测手段,而直接以序列变异作为标记;二是S N P标记分析完全摒弃了经典的凝胶电泳,代之以最新的DNA芯片[12]技术.2 分子标记的应用领域2.1 基因组作图和基因定位研究长期以来,各种生物的遗传图谱几乎都是根据诸如形态、生理和生化等常规标记来构建的,所建成的遗传图谱仅限少数种类的生物,而且图谱分辨率大多很低,图距大,饱和度低,因而应用价值有限.DNA分子标记用于遗传图谱构建是遗传学领域的重大进展之一.1987年,D onis K e ller等发表了第一张人类的RFLP s连锁图,其饱和度远远超过了经典的图谱.近来,L i L i等[13]运用A FLP技术构建太平洋牡蛎(Crasso strea gigas)的一个雄性和一个雌性遗传图谱,雄性图包括96个标记,定位在10个连锁群中,覆盖1258c m;而雌性图谱包括119个标记,分布在11个连锁图中,覆盖993c m.在遗传图谱中大部分的标记很少是对于纯合体的,且彼此间有着紧密的联系,表明在太平洋牡蛎中存在着隐性有害的基因.目前,几乎所有重要农作物,如拟南芥、番茄、玉米、水稻、甜菜、小麦、大豆、马铃薯、棉花、油菜等的RFL P图谱均已构成[14,15].随着新的DNA标记技术的发展,生物遗传图谱名单上的新成员将不断增加,图谱上标记的密度也将越来越高.建立起完整的高密度的分子图谱,就可以定位感兴趣的基因.迄今在玉米、水稻、大豆、小麦和棉花等各种经济作物上已定位了许多农艺性状和经济性状的基因,包括农作物的抗病性、抗虫性、抗逆性、早熟性等,为开展这些基因的分子育种奠定了基础[16].2.2 基于图谱克隆基因图位克隆(M ap based c l on ing)是近几年随着分子标记遗传图谱的相继建立和基因分子定位而发展起来的一种新的基因克隆技术[1,14].利用分子标记辅助的图位克隆无需事先知道基因的DNA 序列,也不必了解基因的表达产物,就可以直接克隆基因.图位克隆技术首先在拟南芥中取得成功,后来在番茄、大麦、小麦、甜菜、水稻的研究中也利用了这一方法.图位克隆是最为通用的基因识别途径,至少在理论上适用于一切基因.基因组研究提供的高密度遗传图谱、大尺寸物理图谱、大片段基因组文库和基因组全序列,已为图位克隆的广泛应用铺平了道路.2.3 物种亲缘关系和系统分类中的应用分子标记广泛存在于基因组DNA 的各个区域,通过对随机分布于整个基因组的分子标记的多态性进行比较,就能够全面评估研究对象的多样性,并揭示其遗传本质.利用遗传多样性的结果可以对物种进行聚类分析,进而了解其系统发育与亲缘关系.1997年,G uiford 等仅用了3个SS R 标记区分出21个苹果品种.2002年,Ivand ic 等用已知图谱位置的33个SSR 对来自以色列、土耳其和伊朗的39份野生型大麦基因型遗传多样分析表明,大多数野生型大麦能按其起源的国家归类[17].Y ang 利用RAPD 技术对23个不同品种的芹菜进行研究,根据其多态性,划分出不同类型,绘出了其进化关系,与以前用蛋白质或用同工酶标记分类的结果一致[18].李太武等[19]用20条能产生清晰可重复扩增产物的随机引物,对皱纹盘鲍、杂色鲍进行RA PD 分析,计算出各群体扩增位点的多态性比例、群体遗传杂合度和群体间的遗传距离,表明皱纹盘鲍与杂色鲍的亲缘关系较远,K linbu ng a 等[20]用R FL P 和RA PD 技术,对泰国的3种鲍H aliatis asinina ,H.ov ina,H.var ia 的18S ,16SrDNA 进行研究,通过小样本图谱分析表明H.as i n ina 和H.ovina 在遗传上比H.varia 近,H.varia 与H.ovina 有明显的遗传上的差异.分子标记的发展为研究物种亲缘关系和系统分类提供了有力的手段.2.4 用于疾病诊断和遗传病连锁分析1980年,Boste i n 等成功的将RFLP 技术用于镰刀型贫血症的诊断分析,开创了基因诊断的先河.RFLP 是以孟德尔方式遗传,因此可以作为染色体上致病基因座位的遗传标志.目前,许多与RFLP 相连锁的致病基因得以定位[21].小卫星和微卫星DNA 因其高度多态性而被广泛用于疾病诊断和遗传病的连锁分析.刘湘帆等采用多个微卫星位点进行血友病B 基因诊断分析,发现6个S TR 位点与血友病B 基因有关[22].近几年研究发现,核苷酸重复长度变异可导致许多疾病.已经证实,Hunti ngon 舞蹈症、脆性X 染色体综合症,肌强直性营养不良等与之有关[23,24].第三代分子标记SNP 被认为是一种稳定遗传的早期突变,因此将S N P 用于单基因与多基因病的连锁分析,显示出广阔的应用前景[9,25].M arti n 等研究表明,A poE 基因附近7个SNP 位点与老年性痴呆症(A lzhei m e r d i sease)有关[26].将S N P 标记技术用于与心血管疾病、神经精神病及肿瘤相关基因的分析均有较多报道[27,28].随着高通量检测SNP 技术方法的出现,作为数量最多且易于批量检测的多态标记,SNP 在连锁分析与基因定位,包括复杂疾病的基因定位、关联分析、个体和群体对环境致病因子与药物的易感性研究中将发挥愈来愈重要的作用.3 分子标记技术的展望目前,分子遗传标记技术已飞速发展,并被广泛应用于动植物的遗传研究中.分子标记中的S N P 已在玉米、大豆、鸡、猪等动植物育种和生产中有许多应用研究,主要集中在基因定位、辅助育种、疾病治疗等方面的应用研究工作,取得了一些应用成果[29~31].DNA 分子标记技术的开发是近年来分子生物学领域研究的热点.随着分子生物学理论与技术的迅猛发展,必将研发出分析速度更快、成本更低、信息量更大的分子标记技术.而分子标记技术与DNA 提取程序化、电泳胶片分析自动化、信息(数据)处理计算机化的结合,必将加速遗传图谱的构建、基因定位、基因克隆、物种亲缘关系鉴别及与人类相关的致病基因的诊断和分析.参考文献:[1]朱玉贤,李毅.现代分子生物学[M ].第二版.北京:高等教育出版社,2002:371-376.[2]W illia m J C K,Kubeli k A R,L i vak K J .DNA po l ymo rph is m a m plifi ed by arb itrary pri m ers are use f u l genetic m arkers[J].N ucle i c A c i ds R es ,1990,18:6531-6537.[3]王志林,赵树进,吴新荣.分子标记技术及其进展[J].生命的化学,2001,21(4):39-42.[4]V e lappam N,Sondgrass J L,H akov irta J R.R apid i den tificati on o f pathogenic bacter i a by si ng l e enzy m e a m plified frag m entleng t h po ly m orphis m ana l ys i s[J].D iagnostic M i c robiology and i nfecti ons D isease ,2001,39:77-83.[5]黎裕,贾继增,王天宇.分子标记的种类及其发展[J].生物技术通报,1999,15(4):19-22.[6]Serap i on J ,K ucudtasH,F eng J ,e t a.l B i o i nfo r ma ti c m i n i ng oft ype I m i ncro sate llite fro m expressed sequence tage o f channe l catfis h(lcta l urns puncta t us)[J].A n i n G enet ,2003,34(6):445.[7]张丽娟,张保军,耿社民.微卫星标记与动物遗传育种[J].黄牛杂志,2003,29(2):49.83第23卷第2期马克世,等:分子标记的发展及应用84 周口师范学院学报2006年3月[8]冯龙仁,牛安欧.微卫星简单重复序列锚定PCR扩增技术(SSR PCR)及其在寄生虫基因研究中的应用[J].实用寄生虫杂志,2000,8(1):33-34.[9]杨昭庆,洪坤学,褚嘉佑.单核苷酸多态性的研究进展[J].国外医学:遗传学分册,2000,23(1):4-8.[10]张效萌,韩金祥.单核苷酸多态性的检测及其在医药领域的应用[J].生命的化学,2001,21(4):325-327.[11]李艳杰,龙火生,李影,等.单核苷酸多态性(S N P)的研究进展和应用[J].A n i m a l Science V eter i ngany M edic i ne,2003,22(4):l6.[12]帕提古丽,罗薇.基因芯片的应用[J].西南民族大学学报,2004,30(3):341.[13]L i L,i G uo X i m i ng.A FLP based gene tic linkage m aps of t he P ac ific O yster Crasso sstrea G i g as T hunbe rg[J].M ar i ne B iotechno,l2004,6:26.[14]张德水,陈受宜.DNA分子标记、基因组作图及其在植物遗传育种上的应用[J].生物技术通报,1998(5):915-912.[15]王建波.ISSR分子标记及其在植物遗传学研究中的应用[J].遗传,2002,24(5):613-616.[16]危文亮,赵应忠.分子标记在作物育种上的应用[J].生物技术通报,2000,22(2):12-16.[17]冯宗云,张义正,凌宏清.大麦基因组中的微卫星标记及应用[J].遗传,2002,24(6):727-733.[18]Y ang X,Q u i ros C F.Identifi cation and classificati on o f ce l ery cultivars w it h RA PD m arkers[J].Theor A pp l G enet,1993,86:205-212.[19]李太武,杨文新,宋林生,等.皱纹盘鲍和杂色鲍遗传多样性的RA PD研究[J].海洋与湖沼,2003,34(7):444.[20]K linbunga S,P ripue P,K ha m nam tong N,et a.l G enetic d i versity and m o l ecular o f the trop i ca l abalone(H a li o ti s asi n i na)i nT hail and[J].M arine B i otech no,l2003(5):505.[21]卢大儒,邱信芳,薛京伦.医学分子遗传学[M].上海:复旦大学出版社,1998:505-506.[22]刘湘帆,王学锋,樊绮诗.联合多个微卫星DNA位点进行血友病B基因诊断[J].中华血液学杂志,2002,23(3):147-150.[23]蔡善荣,刘希永,郑树.人类基因组STR及其应用[J].国外医学:遗传学分册,2000,23(2):60-62.[24]聂胜洁,景强.短串联重复序列的研究及应用[J].国外医学:遗传学分册,2002,25(5):255-259.[25]W eiss K M.H o w m any disease does it take to m ap a gene w it h S N Ps[J].N a t ure genet,2000,26(2):151-155.[26]L aws S M,H one E,G andy S.Expand i ng the associati on bet w een the APOE gene and the risk o fA lzhei m er s'd i sease:possi blero l es fo r APOE prom oter po ly m orphis m s and a lterati ons i n APOE transcr i ption[J].J N eurochem istry,2003,84(6):1215-1236.[27]刘芳,周新.单核苷酸多态性与高脂血症[J].国外医学:临床生物化学与检验医学,2001,22(4):169-170.[28]柯杨.相关基因的单核苷酸多态性与肿瘤的遗传易感[J].中华肿瘤杂志,2002,24(2):105-106.[29]聂庆华,张细权,雷明明.单核苷酸多态性及其在鸡QTL定位上的应用[J].遗传,2003,25(6):729-734.[30]侯娅丽,周倩,刘文忠.单核苷酸多态性及其猪育种中的应用[J].A n i m al Science V eter i nary M ed i c i ne,2004,6:46.[31]郑汗式,刘良国,易祖盛.RA PD分子标记与鱼类种质资源和遗传育种研究[J].广州大学学报,2003,2(6):5l8.D evel op m ent and applicati on of DNA m olecular markersMA Ke shi,HU B i n g y i(D epart m ent o f L ife Sc ience,Zhoukou N orma lU n i ve rsity,Zhoukou466000,Ch i na)Abstrac t:W ith the rap i d deve l op m en t of m o lecular b i o l ogy,DNA mo lecu l a r m arke rs rece i ves m ore and m ore atten tion i n t he w orld.T his paper i ntroduces the deve l op m en t i n DNA m olecu lar m arkers and its appli cation i n genom e m app i ng,gene cloni ng,a nalysis o f relationsh i p of spec i es relationsh i p,as w e ll as d i sease diagnosis.K ey word s:DNA;po l ymo rph i s m;m o l ecular m arke r。
大黄鱼DNA分子标记技术及其在遗传育种中应用研
究的开题报告
针对大黄鱼这一重要经济鱼类的遗传育种瓶颈问题,本研究将探索
利用DNA分子标记技术在大黄鱼遗传育种中的应用,以期为大黄鱼遗传
育种提供理论基础和技术支撑。
本研究将从以下几个方面进行探索:
一、大黄鱼的遗传特征分析
针对大黄鱼的遗传特性,本研究将采用PCR技术,按照已经确定的
大黄鱼基因序列,设计特异性引物,通过PCR扩增出大黄鱼DNA样本中的目标基因段,再通过限制性酶切,测定大黄鱼样本基因型,并进行基
因型比较和分析,为后续的育种工作提供遗传特性分析的依据。
二、大黄鱼的SNP分子标记技术的建立
根据大黄鱼的基因序列,本研究将开发大黄鱼的SNP分子标记技术。
具体操作为在大黄鱼基因序列中筛选出SNP位点,并设计特异性引物,
进行PCR扩增,再通过基因测序,获得目标SNP位点的基因序列数据,最后通过SNP切点的测定,建立大黄鱼SNP分子标记库。
三、大黄鱼遗传育种的应用研究
针对大黄鱼的SNP分子标记技术,本研究将结合大黄鱼遗传特征,
进行遗传育种的应用研究。
首先将对大黄鱼不同品系的DNA样本进行SNP分子标记检测,获得其基因型数据,并对其进行遗传特性分析。
然后,将对大黄鱼的性状进行测定,采集大量不同表现型的大黄鱼个体
DNA样本,进行SNP分子标记检测,筛选出具有良好性状的个体,并进
行遗传育种。
通过本研究的开展,不仅可以为大黄鱼遗传育种提供技术支撑,还
能够为其他重要经济鱼类的遗传育种提供借鉴和启示。
DNA分子标记研究进展第 7 卷第 1 期天津农学院学报 17, 11 N o V o l, 20002000 年 3 月 Jo u rna l o f T ian jin A g r icu ltu ra l Co llege M a rch () 文章编号: 1008- 5394 200001- 0021- 04Ξ分子标记研究进展 DN A1 2 王晓梅杨秀荣 ( )1 天津农学院水产科学系, 天津 300384; 2 天津市植保所, 天津 300112摘要: 分子标记是继形态标记、细胞标记和生化标记之后发展起来的一种新的较为理想的遗传标记。
本文概述了几种主要的 DN A 分子标记的基本原理和特点, 同时对它们的优缺点进行了比较。
关键词: 分子标记原理特点中图分类号: Q 75 文献标识码: A遗传标记是基因型的特殊的易于识别的表现形式, 它是生物分类学、育种学、遗传学和物种起源与 1 进化等研究的主要技术指标之一。
理想的遗传标记主要应具备以下几条标准: ?具有较强的多态性;() ?表现为共显性, 因而能鉴别出纯合基因型和杂合基因型; ?选择中性即无基因多效性; ?对主要的农艺性状无影响; ?经济方便和易于观察记载。
传统上一直沿用形态标记, 生物学研究进入细胞水帄后,() 人们开始利用细胞标记作为遗传标记, 该标记是指染色体核型染色体的数目、大小、着丝粒位置等和()带型带、带、带等。
随着同工酶技术的发展, 又广泛应用了从蛋白质水帄反映生物遗传差异的生 C G N化标记。
这三种标记的数目有限, 往往不能满足研究的需要, 并且它们有些是基因表达加工后的产物, 其标记易受环境条件和发育阶段等因素的影响。
进入80 年代, 随着分子生物学及分子克隆技术的发展和完善, 诞生了直接从水帄上进行遗传分析的分子标记, 在人类基因组计划的推动下, 分子标记的 DN A 研究与应用得到了迅猛的发展。
分子标记直接以的形式表现, 不受环境条件和发育阶段的影响, DN A标记的数目多, 多态性高。
并且有许多分子标记表现为共显性, 能提供完整的遗传信息, 当前分子标记已广泛应用于基因组研究的各个方面。
()1 限制性片段长度多态性 , R FL P R e st r ic t io n F ragm en t L en g th Po lym o rp h ismR FL P 是 80 年代中期发展起来的一种最早的分子标记。
1980 年 Bo ste in 首先提出了利用 R FL P 作为标记构建遗传图谱, 直到 1987 年 D o n is K e lle r 等人才构建出第一张人的 R FL P 图谱。
R FL P 是由于分子中限制性内切酶酶切识别序列中出现碱基的插入、缺失、置换、倒位而导致酶切位点的增减所 DN A2, 3 引起的基因型间限制性片段长度的差异。
它的基本原理是: 不同材料的用已知的限制性内切 DN A 酶消化后, 产生许多大小不等的片段, 电泳分离、印迹转移到硝酸纤维膜上, 用放射性标 DN A So u th e rn记的探针与膜上变性的酶切进行杂交, 放射自显影, 即可显示出不同材料的多态性图谱, 即显示 DN A出所分析的序列间的。
目前, 有 2 种方法可进行的分析。
一是上述原理所述, DN A R FL PDN A R FL P其中用作的探针可以是特殊基因的克隆、克隆、随机的基因组克隆和合成的 R FL P DN A cDN A DN A() 低聚核苷酸克隆。
二是对那些分子量较小的样本如线粒体、核糖体等, 可在酶切后 DN A DN A DN A 4 对其产物直接电泳, 将不同大小的限制性酶切片段分离, 从而得到该的图谱。
DN A DN A R FL PR FL P 标记呈孟德尔式遗传, 大多数 R FL P 的等位基因为共显性, 在任何分离群体中能区分纯合基Ξ 收稿日期: 1999211229 ( ) ) ( , 女汉族, 讲师, 硕士, 主要从事分子生物学、组织学等方面的研究工作。
作者简介: 王晓梅 1962-因型与杂合基因型。
标记的数目多且稳定, 可用于构建高密度的遗传图谱。
但该技术在操作过程中涉及了的提取、酶切、电泳分离、探针的制备和杂交等一系列分子生物学技术, 步骤繁杂, 工 DN A So u th e rn() 作量大, 且分析中需要的量大 2, 10 , 因此在很大程度上限制了技术的应用。
DN A ΛgR FL P()2 随机扩增多态 , RA PD R an dom A m p lif ied Po lym o rp h ic DNA DN A是 1990 年由和领导的 2 个研究小组几乎同时发展起来的建立在基 RA PD W illiam s W e lsh PCR 5, 6 () 础上的一种新型的分子标记技术。
它的基本原理是: 采用随机合成的寡核苷酸通常 10 作 DN A bp反应的引物, 对所研究生物基因组进行扩增。
一般说某一引物在真核基因组中可能有 PCR DN A PCR很多的结合位点, 但只有在 4 000 之内, 存在反向帄行的与该引物互补的双链才能将合成的新 , bp DN A 链作下一次合成的模板, 经 35, 40 个循环, 即可得到大量的片段, 扩增产物经琼脂糖凝胶电泳、 DN A染色, 紫外下显示带纹。
EB RA PD探针; ?使用随机引物, 合成引物时无需预知 RA PD 技术具有其自身独特的优点: ?不需要 DN A被研究的生物的基因组序列, 一套引物可用于不同生物的研究; ?RA PD 技术操作简便、快速, 可免去中的克隆制备、同位素标记、杂交等步骤; ?分析所需量少, 每个反应仅需R FL P So u th e rn RA PD DN A样本受限制的情几十纳克的 DN A , 仅为 R FL P 的1ƒ1 000, 1ƒ200, 这对生物早期取样鉴定或在 DN A 况下大有益处。
但 RA PD 技术也有自身的局限性, 主要表现在两个方面。
第一, RA PD 标记为显性分子标记, 因此不能在代区分纯合体与杂合体, 必须进行分析; 第二, 扩增中退火温度较常规的 F 2 F 3 RA PD 反应低, 一般为 36 ?左右, 这样能保证短核苷酸引物与模板的稳定配对并允许适当的错配, 增大 PCR了引物在基因组中配对的随机性, 提高了对基因组的分析效率, 但是由于采用的引物短, 值DN A Tm低, 扩增结果易受外界因素的影响, 实验的重复性较差, 因此在实验中要严格控制实验条件, 才能得到可重复、理想的扩增效果。
()3 扩增片段长度多态性 , A FL P A m p lif ied F ragm en t L en g th Po lym o rh ism 7 是 1993 年由荷兰公司科学家等人发明的一种分子标记技术, 该技 A FL P k eygen e Zab eau DN A术的基本原理是: 对基因组进行限制性酶切片段的选择性扩增。
主要步骤是: 将基因组进行DN A DN A酶切片段的两端, 从而形成一个带接头的特异片段, 通过接限制性酶切后, 将特定的接头连接在 DN A’ 头序列和引物 3端的识别, 进行扩增, 最终经过变性的聚丙烯酰胺凝胶电泳分离, 通过银染 PCR PCR或放射自显影检测。
技术实际上是将和相结合的一种技术。
该技术既继承了的稳定性, 又具 A FL P R FL P PCR R FL P( )有反应快速、灵敏的特点, 同时克服了和的缺点, 且扩增的带纹多 50, 100 条。
PCR R FL P RA PD的大多数扩增片段与基因组的单一位置相对应, 实验重复性高, 该标记为孟德尔式遗传。
但该技 A FL P术目前在国内多用试剂盒来完成整个实验, 实验成本高; 此外该技术已申请专利, 目前人们只能自由地将该技术用于非赢利性的科学研究, 因此在某种程度上也限制了该技术的应用。
()4 简单序列重复 , SSR S im p le Sequ en ce R ep ea t在真核生物基因组中, 散在分布着由 1, 4 个碱基对组成的简单重复序列, 即它又称为微卫星, SSR()。
序列的侧翼具有保守的序列, 由于重复基数的数目变化大, 因此 DN A m ic ro sa te llite DN A SSR DN ASSR 显示出较强的多态性。
人们把以微卫星 DN A 标记建立的遗传连锁图称为继 R FL P 作图后的第二代遗传连锁图。
1985 年已从人肌红蛋白位点获得了3 核心序列作探针, 与人的酶切后的总 Ieff rey s bp体进行杂交, 得到了杂交带谱, 大多数带纹代表人基因组内不同座位上的卫星它们组合在 , DN A DN A 一起成为人的指纹图, 它有高度的个体差异, 且按孟德尔方式稳定遗传, 此法流行了近 10 , 在法 DN A a8 医鉴定方面作出了贡献。
1993 年和改用小卫星探针显示带谱的方法, 创造了用侧W u T an k sley SSR第 1 期 23? ?王晓梅, 等: DN A 分子标记研究进展翼序列作引物的一部分, 用来扩增重复序列本身, 由于重复序列的长度变化极大, 因此是检测多态性的一种有效的方法。
该技术一般检测到的是一个单一的多等位基因位点, 此标记为共显性, 所得的结果重复性高。
为了提高分辨率, 常使用变性的聚丙烯酰胺凝胶电泳, 它可检测出单拷贝差异。
该标记比 R FL P 8 的多态性更丰富, 且操作上比简便快速, 因此正在被广泛应用。
R FL P ()5 2 SCA R Sequ en ceC h a rac te r ized A m p lif ied R eg io n 标记是 1993 年在技术的基础上发展起来的。
基本流程是: 先作分析, 然后把 SCA R RA PD RA PD 目标的片段回收、克隆和测序, 再根据该序列设计特定引物, 此引物通常包含有原来的引 RA PD RA PD9 物序列, 多为 20, 24 , 再用该引物对所研究的基因组进行扩增, 这样就可以把与原来 bp DN A PCR的片段相对应的单一位点鉴定出来。
该标记为共显性遗传。
由于标记使用的引物长, 因而 RA PD SCA R实验的可重复性高, 因此比和其它利用随机引物的方法在基因定位和作图中的应用要好。
RA PD()6 2单链构象多态性 , SSC P S in g leS t ran d Co n fo rm a t io n Po lym o rp h ism技术的基本原理是根据双链经变性处理后, 进行非变性的聚丙烯酰胺凝胶电泳, 由于SSC P DN A可形成二级结构, DN A 分子在凝胶中的迁移率与其分子量和空间构型有关, 在非变性胶中, 单链 DN A 而这种二级结构与单链 DN A 的序列有关, 且单链 DNA 的迁移率受到其二级结构的影响。