肝干细胞分子生物学研究进展
- 格式:doc
- 大小:3.98 KB
- 文档页数:2




分子生物学与生物技术分子生物学和生物技术是现代生命科学中的两个重要方向,它们相互交织,共同推动生物科学的发展。
本文将从基础概念、应用领域、技术进展等多方面展开分析,全面地探讨这两个领域的内涵和发展。
一、基础概念分子生物学研究的是生物分子之间的相互作用和调控机制,包括DNA、RNA、蛋白质等大分子的结构、功能以及生物化学反应等方面。
生物技术则将分子生物学的研究成果应用到生物工程、生物医学、农业、环境保护等领域,从而实现生产和生活的可持续发展。
二、应用领域1. 生物医学领域生物技术在生物医学领域中有着广泛的应用,如基因治疗、干细胞治疗、生物药物开发等。
分子生物学的不断发展,让医学界能够更精准地定位和治疗疾病。
例如,基因测序技术的广泛应用使得医学家们可以更好地理解疾病的遗传基础,从而为针对性治疗提供了更多有力的依据。
干细胞技术则在组织工程和再生医学方面有着广泛的应用。
而生物药物的出现和不断发展,则彻底改变了传统药物治疗的理念和方式,为医药行业带来了巨大的商业利润。
2. 生物农业领域生物技术在农业领域也有着广泛的应用,例如转基因技术的应用。
转基因作物被开发出来,既能提高农产品的产量,又能提高农产品的质量和抗病能力,从而促进农业的可持续发展。
同时,转基因技术也被应用于动物育种,改善了养殖品种的基因质量和繁殖效率。
3. 生态环保领域生物技术的广泛应用,为生态环保领域的发展带来了新的契机和可能性。
例如,优化菌株和生物反应器技术的发展,可以用于污染水的处理。
生物地球化学循环的研究,则有助于我们更好地理解生态系统和生物圈的运行机制,从而更好地维护人类生存环境的可持续性。
三、技术进展1. 基因编辑技术随着基因编辑技术的不断发展,分子生物学和生物技术的应用领域也得到了极大拓展。
2012年,CRISPR-Cas9技术被发现,在基因治疗、基础生物研究等领域中引发了轰动效应。
这种趋于可控的基因编辑技术,可以通过改变DNA序列,精准地调控基因表达,从而为治疗基因病、提高生物产量等方面带来实质性的变革。

《CLDN6通过调控JAK2-STAT3信号通路对肝癌细胞侵袭、迁移和凋亡的影响及其作用机制的研究》CLDN6通过调控JAK2-STAT3信号通路对肝癌细胞侵袭、迁移和凋亡的影响及其作用机制的研究一、引言肝癌是一种常见的恶性肿瘤,其高侵袭性、高迁移性以及抗凋亡的特性给治疗带来了巨大的挑战。
近年来,细胞间黏附分子(CLDN)家族中的CLDN6逐渐受到研究者的关注。
本文将着重探讨CLDN6如何通过调控JAK2/STAT3信号通路对肝癌细胞的侵袭、迁移和凋亡产生影响,并进一步探讨其作用机制。
二、材料与方法1. 材料本部分研究使用肝癌细胞系作为研究对象,并收集了相关分子生物学试剂和实验设备。
2. 方法通过采用免疫组化、RT-PCR、Western Blot等技术手段,以及细胞培养、细胞侵袭迁移实验、流式细胞术等实验方法,研究CLDN6对肝癌细胞的影响及其作用机制。
三、结果1. CLDN6的表达与肝癌细胞的侵袭、迁移和凋亡的关系研究发现,CLDN6在肝癌组织中的表达与肝癌细胞的侵袭、迁移能力呈正相关,而与肝癌细胞的凋亡能力呈负相关。
2. CLDN6对JAK2/STAT3信号通路的影响通过实验发现,CLDN6能够显著调控JAK2/STAT3信号通路的活性。
具体来说,CLDN6能够促进JAK2的磷酸化,进而激活STAT3,促进其下游靶基因的表达。
3. CLDN6影响肝癌细胞侵袭、迁移和凋亡的机制经过深入研究,我们发现CLDN6通过调控JAK2/STAT3信号通路,影响肝癌细胞的侵袭、迁移和凋亡。
具体而言,JAK2/STAT3信号通路的激活能够促进肝癌细胞的侵袭和迁移,同时抑制肝癌细胞的凋亡。
而CLDN6的表能够通过对该信号通路的调控,实现对肝癌细胞生物学行为的调节。
四、讨论本研究表明,CLDN6在肝癌细胞中发挥着重要的调控作用,其通过影响JAK2/STAT3信号通路的活性,进而影响肝癌细胞的侵袭、迁移和凋亡。
干细胞、癌症和肿瘤干细胞干细胞生物学已经成熟。
证明造血系统中存在干细胞的研究已让位于对一些组织特异性干细胞和祖细胞的分离与研究、对它们的特征和基因表达程序的阐述,以及在再生医学中对它们应用的研究。
也许干细胞最重要和最有用的特征是它的自我更新能力。
通过这一特征能够发现干细胞和癌细胞之间有惊人的相似:肿瘤可能通常起源于正常干细胞的转化,相似的信号通路可能既调节干细胞也调节癌细胞的自我更新,且癌细胞中可能包含有“肿瘤干细胞”(cancer stem cells)----它们是一些极少的具有自我更新不定潜能的驱使肿瘤形成的细胞。
干细胞被定义为永存的细胞,能通过自我更新和在特异组织中分化产生成熟细胞。
在多数组织中特异性干细胞是极少的。
为了研究干细胞的特征,干细胞需经过仔细纯化和预期的鉴定。
尽管有理由认为每种组织来自于一种组织特异性干细胞,但对这些干细胞的严格的鉴定和分离仅在极少的组织中成功的完成。
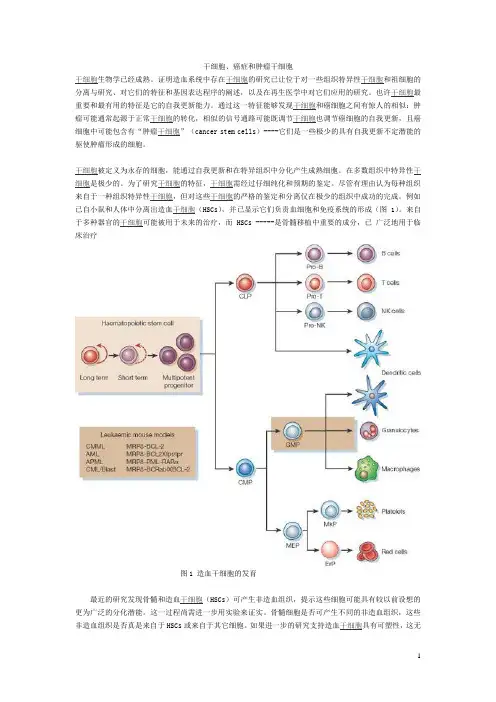
例如已自小鼠和人体中分离出造血干细胞(HSCs),并已显示它们负责血细胞和免疫系统的形成(图1)。
来自于多种器官的干细胞可能被用于未来的治疗,而 HSCs -----是骨髓移植中重要的成分,已广泛地用于临床治疗图1 造血干细胞的发育最近的研究发现骨髓和造血干细胞(HSCs)可产生非造血组织,提示这些细胞可能具有较以前设想的更为广泛的分化潜能。
这一过程尚需进一步用实验来证实。
骨髓细胞是否可产生不同的非造血组织,这些非造血组织是否真是来自于HSCs或来自于其它细胞。
如果进一步的研究支持造血干细胞具有可塑性,这无疑将开创对HSCs的发展潜能的了解,并进一步为临床治疗开辟新的途径。
作为HSCs的特征,在较早的综述中已涵盖了关于它们的分化潜能和临床的应用,在此我们只讨论干细胞生物学中出现的能够提供对肿瘤生物学新认识的证据。
我们将特别聚焦于干细胞与癌细胞相关的三个方面进行讨论。
第一,干细胞与癌细胞在自我更新时有相似的调节机制;第二,瘤细胞有可能来源于正常干细胞的可能性;第三,肿瘤中可能含有“肿瘤干细胞”的观念。

分子生物学研究技术Modul e 1:分子生物学研究技术cDNA文库cDNA文库,代表了生物体某一器官或组织mRNA中所有的或绝大部分的遗传信息。
其构建过程总共有5个步骤(5项技术):1、总RNA 的提取;2、mRNA的纯化;3、cDNA的合成;4、cDNA文库构建;5、基因文库筛选。
详细:1、总RNA的提取:目前常用的提取方法是异硫氰酸胍-苯酚提取法(Trizol提取法),提取步骤:(1)首先用液氮研磨材料成匀浆,加入Trizol试剂,进一步破碎并溶解细胞;(2)加入氯仿抽提,离心,收集含有RNA的水相;(3)用异丙醇沉淀,初步纯化RNA,获得样品用于下一步mRNA的纯化;(PS:实验中还常将含有RNA的细胞破碎物液通过硅胶膜纯化柱后,再通过低盐浓度下从硅胶膜上直接洗脱RNA,获得纯度较高的总RNA);总RNA的浓度、纯度测定:通过分光光度计测其OD260和OD280值,OD260为1时相当于RNA总量为40μg/ml;而OD260/OD280的比值如果在1.8~2.0之间,则表示所提取的RNA纯度较好。
2、mRNA的纯化:纯化原理,将真核细胞的mRNA分子具有5‘端帽子(m7G)和3’端poly (A)尾巴的特征结构,作为提取时的选择性标记。
纯化提取方法:常用寡-纤维素柱层析法获,该方法利用mRNA3‘末端的poly(A)尾巴的特点,当RNA流经寡-纤维柱时,在高盐缓冲液的作用下,mRNA或特异性结合到柱子上,之后再用低盐溶液洗脱mRNA,经过两次层析后可获得较高纯度的mRNA。
3、cDNA的合成:利用RT-PCR技术,常以oligo(dT)为引物,甲基化的dCTP(保证新合成的cDNA链被甲基化,防止构架克隆时被限制性内切酶切割)。
合成基本过程:以mRNA为模板链,在逆转录酶的催化下以甲基化dCTP为原料合成第一条cDNA链;之后再以第一条cDNA 链为模板,在DNA聚合酶催化下合成第二条cDNA(常用RNase H切割mRNA-cDNA杂链中的mRNA序列所产生的小片段为引物合成的第二条cDNA片段,再同过连接酶的作用连接成完整的DNA链。

成体干细胞治疗疑难病成体干细胞是指存在于一种已经分化组织中的未分化细胞,这种细胞能够自我更新并且能够特化形成组成该类型组织的细胞。
成体干细胞存在于机体的各种组织器官中。
成年个体组织中的成体干细胞在正常情况下大多处于休眠状态,在病理状态或在外因诱导下可以表现出不同程度的再生和更新能力。
成体干细胞的研究始于20世纪60年代人们对造血干细胞(hematopoietic stem cells, HSC)的研究。
HSC 是目前研究得最为清楚、应用最为成熟的成体干细胞,它移植治疗血液系统及其他系统恶性肿瘤、自身免疫病和遗传性疾病等均取得令人瞩目的进展,极大促进了这些疾病的治疗,同时也为其他类型成体干细胞的研究和应用奠定了坚实的基础。
成体干细胞的应用研究是再生医学的一个重要组成部分,是很多疾病可供选择的治疗手段。
骨髓来源的干细胞在特定环境中可向肝脏、胰腺、肌肉及神经细胞分化;成体干细胞则可从患者自身骨髓获得,而不存在组织相容性的问题,治疗时可避免长期应用免疫抑制剂对患者的伤害。
骨髓血液中含有大量造血干细胞造血干细胞移植。
人类造血干细胞形态上类似于小淋巴细胞,在骨髓中仅占有核细胞的1%左右。
人类造血干细胞来自胚胎期卵黄囊的间皮细胞,是人体内最独特的体细胞群。
具有极高的自我更新、多向分化与重建长期造血的潜能及损伤后自我修复的能力。
另外还具有广泛迁移和特异的定向(所谓"归巢")特性,能优先定位种植于适当的微环境(如骨髓等处)内,并以非增殖状态和缺乏系列相关性抗原的方式存在。
干细胞移植的安全问题干细胞(stem cells,SC)是一类具有自我复制能力(self-renewing)的多潜能细胞,干细胞(Stem Cell)是一种未充分分化,尚不成熟的细胞,具有再生各种组织器官和人体的潜在功能,医学界称为“万用细胞”。
干细胞分类按照分化潜能的大小,干细胞基本上可以分为以下三种类型:全能性干细胞(totipotent stem cells)它具有形成完整个体的分化潜能。

第二节细胞生物学发展简史与研究动态一、细胞生物学发展简史人类第一次发现细胞到现在已有三百多年的历史。
随着科学技术和实验手段的进步,人们对细胞的认识由浅入深、由表及里,导致了当今细胞生物学的兴起与发展。
根据其发展过程,可分为四个时期,即细胞学说的创立、细胞学的经典时期、实验细胞学的发展和细胞生物学的兴起。
(一) 细胞学说的创立1665 年,英国的物理学家胡克 (R. Hooke) 用自制的显微镜观察了软木( 栎树皮 ) 和其他植物组织,发表了《显微图谱》 (micrographia) 一书,描述了软木是由许多小室组成,状如蜂窝,称之为“细胞” (cell 原意为小室 ) 。
实际上,胡克在软木组织中所看到的仅是植物死细胞的细胞壁。
这是人类第一次看到细胞轮廓,人们对生物体形态的认识首次进入了细胞这个微观世界。
1675 年 (A.V.Leeuwenhoekia) 用自制的高倍放大镜先后观察了池塘水中的原生动物、动物的精子,在蛙鱼的血液中发现了红细胞; 1683 年,他又在牙垢中看到了细菌。
1831 年,布朗 (R. Brown) 在兰科植物的叶片表皮细胞中发现了细胞核。
1835 年,迪雅尔丹 (E.Dujardin) 在低等动物根足虫和多孔虫的细胞内首次发现了透明的胶状物质的内含物,称之为“肉样质” (sarcoide) 。
1836 年,瓦朗丁 (Valentin) 在结缔组织细胞核内发现了核仁。
至此,细胞的基本结构都被发现了。
在 19 世纪以前,许多学者的工作,都着眼于细胞的显微结构方面,主要从事于形态上的描述,而对各种有机体中出现细胞的意义,均未作出理论上的阐述和概括。
1838-1839 年,德国植物学家施莱登 (M.J.Schleiden) 和动物学家施旺(T · Schwann) 根据自己研究和总结前人的工作,首次提也了细胞学说(cell theory) 。
他们认为“一切生物从单细胞到高等动、植物都是由细胞组成的;细胞是生物形态结构和功能活动的基本单位”。

细胞生物学开展简史一细胞的发现和细胞学说的建立1665年,英国学者Robert Hooke用自制的显微镜观察栎树软木塞的薄切片,发现了其中有许多蜂窝状的小室,并将这些小室命名为cell。
实际上当时看到的是植物细胞的细胞壁。
此后,生物学家用cell一词描述生物体的根本构造单位,中文翻译为细胞。
Hooke对有关细胞的首次描述见于1665年他的?显微图谱?中,因此人们认为细胞的发现是在1665年。
真正观察活细胞的是荷兰科学家Antony von Leeuwenhoek,他用设计较好的显微镜观察池塘水中的原生动物、蛙肠道的原生动物、人类和哺乳动物的精子,并于1674年在观察鱼的红细胞时描述了细胞核的构造。
由以上可见,细胞生物学的根底建立于17世纪,并且Hooke和Leeuwenhoek两位科学家为此做出了重要奉献。
在Hooke发现细胞后的近170年中,人们用光学显微镜相继发现了一些不同类型的细胞,但对细胞的认识根本上没什么新的进展。
直到19世纪30年代,显微镜制造技术有了明显的改良,分辨率提高到1μm以;同时还由于切片机的制造成功,从而对细胞的观察有了许多新的进展,细胞核、核仁、细胞的原生质等被提醒,人们才真正认识到细胞的生物学意义。
1838~1839年,德国植物学家Scheleiden〔1838年〕和动物学家Schwann〔1839年〕总结前人的工作,综合了植物和动物组织中细胞的构造,提出了“细胞学说〔cell theory〕〞,指出“一切生物,从单细胞生物到高等动、植物都是由细胞组成的;细胞是生物体构造和功能的根本单位〞。
后来德国科学家Rudolf Virchow(1855年)明确提出“一切细胞只能来自原来的细胞〞的论点。
此外,他还指出机体的一切病理现象都是基于细胞的损伤,从而论证了生物界的统一性和共同起源。
二细胞学的形成和开展〔一〕细胞学说的建立把生物学的注意力引向细胞,有力地推动了对细胞的研究。
【关键词】 肝干细胞 分子 机制
0 引言 肝干细胞的可塑性及其分化机制的 研究 在基础 理论 和临床 应用 等方
面均有重要意义。一方面,肝干细胞可能参与肝脏损伤的修复与重建,研究肝干细胞的分化
机制有助于阐明肝脏的发育机制;另一方面,肝干细胞分化为具有功能的成熟肝细胞,将为
肝细胞移植和生物型人工肝提供重要的细胞来源。同时,阐明这些 问题 也可为临床肿瘤等
疾病的基因 治疗 提供新的治疗手段[1]。肝干细胞分化过程和机制较为复杂,关于肝干细胞
分子机制的研究成为肝干细胞研究的重要方面,现将其进展做一综述。编辑。 肝干细胞
的基本特征可概括为两点:(1)具有双向分化能力,可向肝细胞和胆管细胞分化;(2)具有自
我更新能力。 目前 肝干细胞范畴的界定还比较混乱,但较为一致的观点是:卵圆细胞(oval
cell)、胎肝细胞、小肝细胞和骨髓造血干细胞都是肝干细胞的候选者。
1 卵圆细胞 肝脏受损后的再生一般情况下由肝实质细胞分裂增殖来完成,而当肝实
质细胞严重受损不能增生或受到有丝分裂原抑制剂损伤时,位于肝内低分化的卵圆细胞即被
激活,进一步增殖、分化为肝细胞和胆管细胞来完成肝脏的结构与功能重建,卵圆细胞的形
态学和免疫组织化学方面与胆管上皮细胞相似,形态呈卵圆形。有学者[2]建立了一种新的促
肝脏卵圆细胞增殖的小鼠模型,研究发现,这种卵圆细胞在表达scal、cd34及cd35的同
时,还表达a6及afp,说明scal、cd34及cd35是小鼠肝卵圆细胞的特异标志。由于目
前尚未发现肝干细胞的特异性表面标志,故有关肝干细胞的分离纯化仍不成熟,近来有人[3]
利用绿色荧光蛋白转基因小鼠的肝细胞的荧光活性纯化并富集肝干细胞,并发现
cd45(-)ter-ll9(-)afp(+)的高表达gfp的幼稚细胞,具有增殖分化潜力,可以分化为肝细胞
和胆管细胞。tnf在肝干细胞的活化及肿瘤的发生中有重要作用。研究者[4]观察到,在缺乏
胆碱而补充乙硫氨基酪酸饮食的大鼠卵圆细胞增殖中,tnf具有正调节作用,同时卵圆细胞
本身也表达tnf。在tnfir基因敲除小鼠,卵圆细胞的增殖被大大削弱,肿瘤的形成明显减
弱。生长因子受体酪氨酸激酶信号在正常肝细胞生长的调节中有重要作用,研究发现[5]受体
型酪氨酸激酶家族中tie2、cmet、flk1在实验性大鼠肝癌细胞中比正常肝细胞中
显著增高。其中后两者在gstp(+)的癌前病变中过量表达,而tie2在内皮细胞和卵圆细
胞上有表达,考虑tie2、cmet、flk1与肝癌的发生有关。在人类肝脏,卵圆细胞
的数量随肝损伤的加重而增多。这种卵圆细胞的增生是肝干细胞对肝损伤的反应而不是受损
肝细胞的管样化生[6]。
2 胚胎肝细胞 有人[7]将胚胎13.5天的胎肝细胞用绿色荧光蛋白(egfp)标记,移植
入小鼠肝中,结果发现移植后胎肝细胞afp的表达逐渐降低,而alb的表达逐渐升高。为了
了解肝脏发育过程中肝干细胞标志的表达情况,有学者[8]选用干细胞标志thy1和肝细胞标
志ck18进行研究,分别选取胚胎第16、18、20天及新生大鼠的肝脏,分离提取肝细胞,采
用磁珠分离法纯化胎肝细胞,结果发现,在磁珠分离前,所有的胎肝细胞均表达thy1,磁珠
分离后,仅在部分胎肝细胞表达thy1,而ck18在磁珠分离前后的胎肝细胞上均表达,说明
在胎肝中有不同的肝细胞群同时存在,其中一部分具有干细胞特征。有学者[9]成功分离人胎
肝上皮细胞,在体外培养数月后,通过门静脉移植入小鼠肝脏,在移植后一小时,小鼠肝脏
中有1.5%的移植细胞,继而发现14%~55%的移植细胞进入小鼠肝脏,移植细胞占小鼠肝脏细
胞总数的1%。
4 骨髓造血干细胞 近年发现成年个体的造血干细胞可塑性很大,可以分化为所有种
类的组织细胞。有学者[11]首先在大鼠中发现一些骨髓细胞来源的卵圆细胞和肝细胞,研究
者将雄性大鼠的骨髓细胞移植入经放射线致死照射的雌性大鼠,然后用2-乙酰氨基芴和四氯
化碳损伤此受体鼠,此后在受体鼠的肝脏中发现了y染色体阳性的卵圆细胞和肝细胞。有学
者[12]对骨髓来源肝干细胞研究发现,thy1(+)、β2微球蛋白(-)的骨髓细胞能分化为形
态和功能都与肝实质细胞相同的细胞,分化细胞在超微结构上与肝实质细胞完全相同,而且
在大鼠及人的正常肝脏及病肝中也可发现thy1(+)、β2微球蛋白(-)细胞的存在。所以
thy1(+)、β2微球蛋白(-)的骨髓细胞被认为具有肝干细胞的特征。 骨髓中造血干
细胞一方面可以在病肝中分化为肝实质细胞,从而恢复肝功能,一方面又可以作为肝病基因
治疗的载体。美国研究者[13]将大鼠异体骨髓造血干细胞和成熟肝细胞分别通过门静脉注射
入原位肝移植后发生排斥反应的大鼠肝脏,结果发现骨髓造血干细胞中约62%的细胞定位于
肝脏,而成熟肝细胞只有约2.5%定位于肝脏,而且移植的造血干细胞表现出肝细胞和胆管细
胞的特征。有学者[14]报道人骨髓中的造血干细胞也可以分化为肝细胞,用dna特异性探针
在一曾接受男性骨髓移植的女性病人肝脏中发现有y染色体阳性的肝细胞,而将曾接受男性
骨髓移植的女性病人肝脏再移植给另一男性患者,结果在被移植的肝组织中发现有y染色体
阳性的肝细胞,提示骨髓造血干细胞可以定位于肝脏并分化为肝细胞。