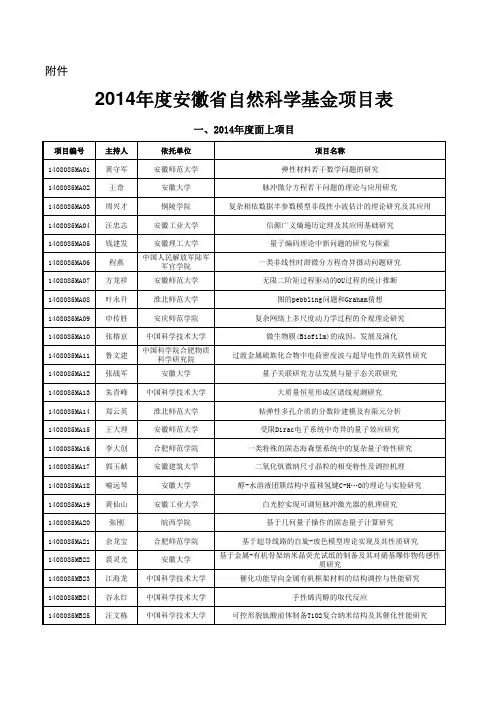
L_精氨酸高产菌的选育及基于代谢流量分布的育种机制_许正宏
- 格式:pdf
- 大小:224.23 KB
- 文档页数:6
第36卷第4期 2006年12月工业微生物IndustrialMicrobiology Vol.36No.4 Dec.2006
国家0十五0科技重点攻关项目(2004BA713B07-05)作者简介:许正宏(1971~),男,博士,副教授,主要从事微生物资源开发与利用的研究。*通讯作者,电话:86-510-85862412;E-mail:Wytao@sytu.edu.cn
L-精氨酸高产菌的选育及基于代谢流量分布的育种机制许正宏, 熊筱晶, 窦文芳, 陶文沂*(江南大学生物工程学院无锡214036)
摘 要 从分析钝齿棒杆菌(Corynebacteriumcrenatum)的精氨酸合成途径入手,提出了一种通过选育脯氨酸结构类似物抗性突变株以提高其精氨酸合成能力的育种思路。采用亚硝基胍(NTG)诱变处理出发菌株YD8(His-,SGr1.2mg/mL,D-Argr15mg/mL),经含15mg/mL的脯氨酸结构类似物S-甲基半胱氨酸(S-MC)的抗性筛选获得精氨酸高产突变株YDM403(His-,SGr
1.2mg/mL,D-Argr15mg/mL,S-MCr15mg/mL),产酸水平可达29.4g/L,较出发菌株YD8的产酸高出55.0%。通过代谢流量分布分析了菌株YD8和YDM403代谢网络的变化,结果表明,菌株YD8可能存在顺序反馈抑制作用,出发株YD8解除了Arg对Glu到Arg的反馈抑制,而YDM403又解除了Pro对Glu到Pro的反馈抑制,从而使中间物Glu累积量下降而对A-KG到Glu不再有反馈抑制,其通量提高,与此同时从Glu向Arg的代谢通量也相应增加。关键词:钝齿棒杆菌; 精氨酸; 代谢流量分布; 菌种选育
对于工业微生物细胞而言,在特定的生理条件下,其代谢表型(Metabolicphenotype)可以通过胞内代谢网络中各种代谢物的流量来进行表征[1]。代谢流量分布(metabolicfluxdistribution)是一种研究胞内代谢的方法,是根据胞内主要反应的化学计量模型和胞内代谢产物的质量平衡来计算胞内的代谢流量[2],通过较准确的代谢流的定量分析,找出影响目标代谢物过量合成的0瓶颈0因素是代谢工程的重要目标之一。氨基酸代谢是代谢工程研究领域的重要课题之一,国外已将代谢流分布的分析广泛用于研究氨基酸菌种在不同条件下代谢表型的改变,进而达到理性优化发酵工艺的目的,如谷氨酸[3]和赖氨酸[10]发酵条件的优化,谷氨酸棒杆菌在不同渗透压协迫条件下代谢流分布的改变[4]等。近年来,国内学者也有利用代谢流量分析来指导L-异亮氨酸发酵条件的优化[5],还有采用该技术手段分析L-缬氨酸育种机制的研究报道[6]等。 L-精氨酸(L-arginine,以下简称Arg)是一种碱性氨基酸,在医药和食品工业有较广泛的用途[7],近年来备受氨基酸工业的广泛关注。目前世界上仅日本等国实现了Arg的发酵法生产,其产量可达60g/L。而我国主要依靠毛发水解提取法制备Arg,发酵法生产的研发尚处于开始阶段。国际上有少量采用基因工程手段进行Arg菌种构建的报道,但从实际的产酸能力看,远未达到工业化生产的水平,因而国内外普遍采用的Arg菌种的选育技术仍以传统的诱变育种为主,其中又以Arg结构类似物抗性筛选为主要方法[8,9]。本文从分析钝齿棒杆菌YD8的Arg及相关氨基酸合成途径入手,提出了一种Arg高产菌株选育的新方法,并通过代谢流量分析比较了出发菌株YD8和高产菌株YDM403的代谢流分布的变化,解析了突变株代谢调节模式的可能的变化,为下一步采用代谢工程手段改良L-精氨酸高产菌株的研究奠定了一定的理论基础。
1 材料与方法1.1 菌株 钝齿棒杆菌CorynebacteriumcrenatumYD8
(His-,SGr1.2mg/mL,D-Argr15mg/mL)为本实验室保藏的L-精氨酸生产菌株,突变株YDM403
)1)(His-,SGr1.2mg/mL,D-Argr15mg/mL,S-MCr15mg/mL)由YD8经NTG诱变而来。1.2 培养基及培养条件 (1)完全培养基(g/L):葡萄糖20,牛肉膏10,蛋白胨10,酵母膏5,NaCl5,琼脂20。pH7.0~7.2,121e灭菌20min。 (2)基础培养基(g/L):葡萄糖10,(NH4)2SO43,KH2PO41,MgSO4#7H2O0.5,FeSO4#7H2O0.02,MnSO4#7H2O0.02,琼脂20,生物素3@10-5,硫胺素2@10-4,L-组氨酸5@10-4。pH7.0~7.2,121e灭菌20min。 (3)种子培养基(g/L):葡萄糖30,玉米浆20,(NH4)2SO420,KH2PO41,MgSO4#7H2O0.5,尿素1.5。pH7.0~7.2,121e灭菌20min,装液量30mL/250mL。 (4)发酵培养基(g/L):葡萄糖120,玉米浆10,(NH4)2SO450,KH2PO41.5,MgSO4#7H2O0.5,Fe-SO4#7H2O0.02,MnSO4#H2O0.02,生物素8@10-5,L-组氨酸5@10-4,CaCO330。pH7.0~7.2,121e灭菌10min,装液量15mL/250mL。 如无特别说明,种子培养采用30~31e,往复式摇床(无锡查桥轻机厂)96r/min培养15~16h后,以10%的接种量接入发酵培养基中,31e往复式摇床96r/min培养96h。5L全自动发酵罐(上海保兴生物工程有限公司)实验中接种量和培养温度同摇瓶实验,通风量为0.75vvm,搅拌速度为500r/min,自动流加氨水控制pH7.2左右。1.3 诱变方法和突变株的筛选 选用亚硝基胍(NTG)为诱变剂,采用适当浓度S-甲基半胱氨酸(S-MethylCysteine,简称S-MC)的平板作为筛选平板。 菌株YD8种子培养到对数生长中后期再以50%的接种量转接到新鲜种子培养基中培养5h使细胞达到同步培养。取培养液10mL无菌离心,收集菌体,用0.1mol/L,pH6.0的磷酸缓冲液洗涤2~3次,转入带玻璃珠的三角烧瓶中打散,得菌悬液,细胞浓度控制在108个/mL左右。称取少量亚硝基胍溶解在少量丙酮中,再加入磷酸缓冲液,制成浓度为1.0mg/mL的亚硝基胍溶液。取一定量的菌悬液中加入亚硝基胍溶液,使其终浓度约0.2mg/mL,30e振荡20min,诱变结束后快速离心终止反应,用0.1mol/L,pH6.0的磷酸缓冲液洗涤2~3次。最后取菌悬液按梯度稀释涂布筛选平板,31e恒温培养3~5d后随机挑选菌落进行初筛。 配制基本培养基,灭菌后分别加入S-MC使终浓度达到10mg/mL、12mg/mL、15mg/mL、18mg/mL。再在各个不同S-MC浓度的平板上涂布经适当稀释的未经诱变处理的YD8菌悬液,观察生长情况,确定出发菌株YD8耐受S-MC的浓度。然后将诱变处理的菌悬液涂布到高于此浓度的S-MC药物平板上,置于31e培养3~5d,随机挑选单菌落进行摇瓶发酵实验进行高产突变株的初筛。1.4 有机酸的测定 采用高效液相色谱仪HP1100(惠普公司)测定
发酵液中的有机酸含量,柱条件如下: 色谱柱为ZORBAXSBC18(4.6mm@250mm),
流动相为0.1%(v/v)磷酸溶液,流速为1mL/min,柱温25e,检测波长为210nm结构类似物。通过归一化法计算各有机酸的含量。1.5 氨基酸的测定 采用氨基酸分析仪1100(安捷伦公司)分析发
酵液中各氨基酸的含量。
2 结果与讨论2.1 Arg的代谢途径分析及育种策略的提出 不同的微生物在Arg生物合成途径及调节机制
上有所不同,已有文献对微生物的Arg生物合成途径进行了很好的综述[8,9]。本文选用的产Arg的出发菌株为实验室选育的钝齿棒杆菌YD8,该菌利用葡萄糖底物生物合成Arg的途径是A-酮戊二酸衍生类型,即谷氨酸类型。在棒杆菌中与Arg合成途径相关的主要分支途径为L-脯氨酸合成途径。这两个氨基酸不仅各自对经由谷氨酸合成自身的后续代谢途径中的酶具有终产物抑制作用,而且还会对它们的共同合成途径,即由葡萄糖底物合成谷氨酸的代谢途径中的酶起到协同反馈调节作用。结合代谢育种原理以及代谢网络调节的观点,Arg的育种策略除采用Arg结构类似物抗性来解除Arg对其代谢途径中的酶的反馈抑制外,还可以尝试通过结合选育脯氨酸的结构类似物抗性突变株来解除Arg和脯氨酸对共同途径中酶)2)
第4期 工 业 微 生 物第36卷 的协同反馈调节的作用,以增加前体物谷氨酸的合成量,从而达到过量合成Arg的目的。 针对上述假设,作者通过NTG诱变处理出发菌株YD8,进而通过筛选脯氨酸结构类似物)))S-甲基半胱氨酸(S-MC)(图1)抗性突变株,以期获得图1 S-甲基半胱氨酸和L-脯氨酸的结构式比较Arg的高产突变株。2.2 高产Arg的突变株YDM403的获得 根据1.3方法采用NTG处理出发菌株YD8,
经含15mg/mL的S-MC的抗性平板处理和摇瓶筛选获得Arg高产突变株YDM403(His-,SGr1.2mg/mL,D-Argr15mg/mL,S-MCr15mg/mL),后者的产酸水平可达29.4g/L,较出发菌株YD8的产酸水平高出55.0%。2.3 菌株YD8与菌株YDM403的发酵过程的比较及代谢流量分析 为进一步分析出发株和突变株在代谢网络上的
变化,在同一发酵条件下,采用5L全自动控制发酵罐实验分析比较了菌株YD8和YDM403的发酵过程,结果如图2、图3及图4:
图2 发酵过程中葡萄糖的消耗和Arg合成的变化曲线rGlucose tArginine A:YD8 B:YDM403
图3 菌株YD8发酵过程中副产物的生成曲线rSerine tLysine vLacticacidoAcetate @Pyruvate图4 菌株YDM403发酵过程中副产物的生成曲线
rAlanine tProline vLacticacid oAcetate @Pyruvate 由图2、3可以看出,菌株YD8在发酵过程中有丝氨酸和赖氨酸生成,由此可知,部分葡萄糖经旁路代谢生成了丝氨酸,且葡萄糖经TCA循环转化成A-酮戊二酸后,碳架流经过TCA循环部分转化为赖氨酸,赖氨酸的生成量为3.02g/L,而Arg的生成量
仅为12g/L。 由图2和4可知,出发菌株经过NTG诱变处理
得到的S-MC抗性突变株YDM403在相同发酵条件下,Arg的产量为32.8g/L,且在发酵过程中无丝氨酸和赖氨酸的生成,但是有少量的脯氨酸和丙氨
)3)
第4期许正宏,等:L-精氨酸高产菌的选育及基于代谢流量分布的育种机制第36卷