细胞周期,癌症与诺贝尔奖
- 格式:doc
- 大小:76.50 KB
- 文档页数:4





分子生物学是一门研究分子基础的生物学领域,包括研究生命过程中使用的分子机制,如基因表达、蛋白质合成和信号传导。
以下是一些与分子生物学有关的诺贝尔奖得主:
•1969年,阿尔伯特·舒尔茨和朱利安·韦伯获得生理或医学奖,因为他们发现了如何利用细胞内的酶来把DNA复制成RNA。
•1989年,罗伯特·基因、孟德尔和爱德华·摩尔获得生理或医学奖,因为他们发现了基因的结构和功能。
•1993年,罗伯特·霍金斯、埃利奥特·拉扎罗和菲利普·范德比尔特获得生理或医学奖,因为他们发现了DNA修饰的分子机
制。
•2006年,罗伯特·费希尔、约翰·科斯塔和爱德华·摩尔获得生理或医学奖,因为他们发现了细胞周期的分子机制。
•2009年,约翰·科斯塔、伊莎贝尔·哈伯和路易斯·托马斯获得生理或医学奖,因为他们发现了细胞内的质膜受体的作用。
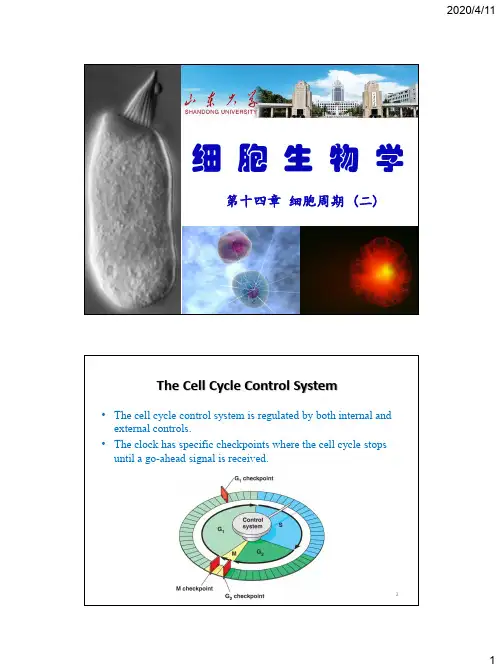
细胞生物学第十四章细胞周期(二)The Cell Cycle Control System•The cell cycle control system is regulated by both internal and external controls.•The clock has specific checkpoints where the cell cycle stops until a go-ahead signal is received.2细胞周期调控一、细胞周期调控因子的发现二、细胞周期调控因子三、细胞周期调控的机制四、其他3一、细胞周期调控因子的发现1、有丝分裂促进因子(mitosis promoting factor,MPF)2、成熟促进因子(maturation promoting factor,MPF)3、细胞分裂周期基因(cell division cycle gene,cdc)4、细胞周期蛋白(cyclin)41、MPF 的发现5G1期PCC S 期PCC G2期PCC1970年,Johnson RT和Rao PN 发现M 期HeLa 细胞与间期细胞融合形成早熟染色体凝集(PCC )。
这一现象提示在M 期细胞中存在诱导染色体凝集的因子,称为有丝分裂促进因子(mitosis promotingfactor ,MPF )。
•1971年,Masui Y 和Markert CL 通过非洲爪蟾卵实验发现成熟卵母细胞细胞质中含有促卵母细胞成熟的因子,称为成熟促进因子(maturation promoting factor ,MPF )。
卵细胞成熟示意图:细胞质移植实验发现MPF :MPF 的发现•1988年,James Maller 实验室在爪蟾卵中分离出MPF ,证明MPF 由p32和p45两种蛋白构成,p32和p45结合后表现出蛋白激酶的活性。
7MPF 激酶p32p45MPF 的组成•1970s ,Leland Hartwell 利用芽殖酵母发现了几十个细胞分裂周期(cell division cycle,cdc )基因,如cdc28。


诺贝尔生理学或医学奖历年获奖者(1901-2019)年份得主国家得奖原因1901年埃米尔·阿道夫·冯·贝林德国“对血清疗法的研究,特别是在治疗白喉应用上的贡献,由此开辟了医学领域研究的新途径,也因此使得医生手中有了对抗疾病和死亡的有力武器”1902年罗纳德·罗斯[英国“在疟疾研究上的工作,由此显示了疟疾如何进入生物体,也因此为成功地研究这一疾病以及对抗这一疾病的方法奠定了基础”1903年尼尔斯·吕贝里·芬森丹麦“在用集中的光辐射治疗疾病,特别是寻常狼疮方面的贡献,由此开辟了医学研究的新途径”1904年伊万·巴甫洛夫俄罗斯“在消化的生理学研究上的工作,这一主题的重要方面的知识由此被转化和扩增”1905年}罗伯特·科赫德国“对结核病的相关研究和发现”1906年卡米洛·高尔基意大利“在神经系统结构研究上的工作”圣地亚哥·拉蒙-卡哈尔西班牙*1907年夏尔·路易·阿方斯·拉韦朗法国“对原生动物在致病中的作用的研究”1908年伊拉·伊里奇·梅契尼科夫俄罗斯“在免疫性研究上的工作”保罗·埃尔利希德国1909年埃米尔·特奥多尔·科赫尔瑞士“对甲状腺的生理学、病理学以及外科学上的研究”1910年阿尔布雷希特·科塞尔德国“通过对包括细胞核物质在内的蛋白质的研究,为了解细胞化学做出的贡献”1911年阿尔瓦·古尔斯特兰德—瑞典“在眼睛屈光学研究上的工作”1912年亚历克西·卡雷尔法国“在血管结构以及血管和器官移植研究上的工作”1913年夏尔·罗贝尔·里歇法国“在过敏反应研究上的工作”1914年@罗伯特·巴拉尼奥地利“在前庭器官的生理学与病理学研究上的工作”1919年朱尔·博尔代比利时“免疫性方面的发现”1920年奥古斯特·克罗丹麦“发现毛细血管运动的调节机理”~1922年阿奇博尔德·希尔英国“在肌肉产生热量上的发现”奥托·迈尔霍夫德国“发现肌肉中氧的消耗和乳酸代谢之间的固定关系”1923年弗雷德里克·格兰特·班廷加拿大%“发现胰岛素”约翰·麦克劳德加拿大1924年威廉·埃因托芬荷兰“发明心电图装置”1926年约翰尼斯·菲比格,丹麦“发现鼠癌”1927年朱利叶斯·瓦格纳-尧雷格奥地利“发现在治疗麻痹性痴呆过程中疟疾接种疗法的治疗价值”1928年查尔斯·尼柯尔法国“在斑疹伤寒研究上的工作”1929年"克里斯蒂安·艾克曼荷兰“发现抗神经炎的维生素”弗雷德里克·霍普金斯爵士英国“发现刺激生长的维生素”1930年卡尔·兰德施泰纳奥地利“发现人类的血型”【1931年奥托·海因里希·瓦尔堡德国“发现呼吸酶的性质和作用方式”1932年查尔斯·斯科特·谢灵顿爵士英国“发现神经元的相关功能”埃德加·阿德里安英国1933年托马斯·亨特·摩尔根美国“发现遗传中染色体所起的作用”1934年乔治·惠普尔美国“发现贫血的肝脏治疗法”乔治·迈诺特\美国威廉·莫菲美国1935年汉斯·斯佩曼德国“发现胚胎发育中的组织者(胚胎发育中起中心作用的胚胎区域)效应”1936年,亨利·哈利特·戴尔爵士英国“神经冲动的化学传递的相关发现”奥托·勒维奥地利1937年圣捷尔吉·阿尔伯特匈牙利“与生物燃烧过程有关的发现,特别是关于维生素C和延胡索酸的催化作用”·1938年海门斯比利时“发现窦和主动脉机制在呼吸调节中所起的作用”1939年格哈德·多马克德国“发现百浪多息(一种磺胺类药物)的抗菌效果”1943年亨利克·达姆丹麦】“发现维生素K”爱德华·阿德尔伯特·多伊西美国“发现维生素K的化学性质”1944年约瑟夫·厄尔兰格美国“发现单神经纤维的高度分化功能”赫伯特·斯潘塞·加塞\美国1945年亚历山大·弗莱明爵士英国“发现青霉素及其对各种传染病的疗效”恩斯特·伯利斯·柴恩英国>霍华德·弗洛里爵士澳大利亚1946年赫尔曼·约瑟夫·马勒美国“发现用X射线辐射的方法能够产生突变”1947年卡尔·斐迪南·科里美国“发现糖原的催化转化原因”格蒂·特蕾莎·科里美国贝尔纳多·奥赛阿根廷“发现垂体前叶激素在糖代谢中的作用”1948年保罗·赫尔曼·穆勒瑞士…“发现DDT是一种高效杀死多类节肢动物的接触性毒药”1949年瓦尔特·鲁道夫·赫斯瑞士“发现间脑的功能性组织对内脏活动的调节功能”安东尼奥·埃加斯·莫尼斯葡萄牙“发现前脑叶白质切除术对特定重性精神病患者的治疗效果”1950年菲利普·肖瓦特·亨奇}美国“发现肾上腺皮质激素及其结构和生物效应”爱德华·卡尔文·肯德尔美国塔德乌什·赖希施泰因瑞士1951年&马克斯·泰累尔南非“黄热病及其治疗方法上的发现”1952年赛尔曼·A·瓦克斯曼美国“发现链霉素,第一个有效对抗结核病的抗生素”1953年汉斯·阿道夫·克雷布斯英国“发现柠檬酸循环”弗里茨·阿尔贝特·李普曼美国“发现辅酶A及其对中间代谢的重要性”1954年约翰·富兰克林·恩德斯美国“发现脊髓灰质炎病毒在各种组织培养基中的生长能力”弗雷德里克·查普曼·罗宾斯美国托马斯·哈克尔·韦勒美国1955年阿克塞尔·胡戈·特奥多尔·特奥雷尔瑞典“发现氧化酶的性质和作用方式”1956年安德烈·弗雷德里克·考南德、美国“心脏导管术及其在循环系统的病理变化方面的发现”沃纳·福斯曼德国迪金森·伍德拉夫·理查兹美国1957年)达尼埃尔·博韦意大利“发现抑制某些机体物质作用的合成化合物,特别是对血管系统和骨骼肌的作用”1958年乔治·韦尔斯·比德尔美国“发现基因功能受到特定化学过程的调控”爱德华·劳里·塔特姆美国乔舒亚·莱德伯格美国“发现细菌遗传物质的基因重组和组织”1959年阿瑟·科恩伯格美国“发现核糖核酸和脱氧核糖核酸的生物合成机制”塞韦罗·奥乔亚美国1960年弗兰克·麦克法兰·伯内特爵士澳大利亚“发现获得性免疫耐受”彼得·梅达沃英国1961年盖欧尔格·冯·贝凯希(美国“发现耳蜗内刺激的物理机理”1962年佛朗西斯·克里克英国“发现核酸的分子结构及其对生物中信息传递的重要性”詹姆斯·杜威·沃森美国【莫里斯·威尔金斯英国1963年约翰·卡鲁·埃克尔斯爵士澳大利亚“发现在神经细胞膜的外围和中心部位与神经兴奋和抑制有关的离子机理”艾伦·劳埃德·霍奇金英国安德鲁·赫胥黎英国1964年康拉德·布洛赫美国“发现胆固醇和脂肪酸的代谢机理和调控作用”费奥多尔·吕嫩德国1965年方斯华·贾克柏法国“在酶和病毒合成的遗传控制中的发现”安德列·利沃夫法国贾克·莫诺(法国1966年裴顿·劳斯美国“发现诱导肿瘤的病毒”查尔斯·布兰顿·哈金斯美国“发现前列腺癌的激素疗法”1967年|拉格纳·格拉尼特瑞典“发现眼睛的初级生理及化学视觉过程”霍尔登·凯弗·哈特兰美国乔治·沃尔德美国%1968年罗伯特·W·霍利美国“破解遗传密码并阐释其在蛋白质合成中的作用”哈尔·葛宾·科拉纳美国马歇尔·沃伦·尼伦伯格美国1969年马克斯·德尔布吕克美国“发现病毒的复制机理和遗传结构”阿弗雷德·赫希美国萨尔瓦多·卢瑞亚*美国1970年朱利叶斯·阿克塞尔罗德美国“发现神经末梢中的体液性传递物质及其贮存、释放和抑制机理”乌尔夫·冯·奥伊勒瑞典。

1、1901年,埃米尔•阿道夫•冯•贝林(德国)。
利用血清疗法治疗白喉。
2、1902年,Ronald Ross(英国)。
关于疟疾的研究。
3、1903年,Niels Ryberg Finsen(丹麦)。
利用光辐射治疗狼疮。
4、1904年,巴甫洛夫(俄国)。
在神经生理学方面,提出了著名的条件反射和信号学说。
5、1905年,R.柯赫(德国)。
关于结核方面的研究和发现。
6、1906年,C.高尔基(意大利),桑地牙哥•拉蒙卡哈(Santiago Ramón y Cajal,西班牙)。
关于神经系统结构的研究。
7、1907年,Charles Louis Alphonse Laveran(法国),发现原生动物在引起疾病中的作用。
8、1908年,Ilya Ilyich Mechnikov(俄国),Paul Ehrlich(德国)。
关于免疫方面的研究。
9、1909年,Emil Theodor Kocher(瑞士)。
关于甲状腺生理学,病理学和外科学方面的研究10、1910年,艾布瑞契•科塞尔(Albrecht Kossel,德国)。
关于细胞化学尤其是蛋白质和核酸方面的研究11、1911年,Allvar Gullstrand(瑞典)。
关于眼睛屈光学方面的研究。
12、1912年,Alexis Carrel(法国。
关于血管缝合以及血管和器官移植方面的研究。
13、1913年,Charles Robert Richet(法国)。
关于过敏反应的研究。
14、1914年,Robert Bárány(奥地利。
关于内耳前庭装置生理学及病理学方面的研究。
15、1915年-1918年,未颁奖,奖金划拨到生理医学奖专门的基金上。
16、1919年,Jules Bordet(比利时)。
关于免疫方面的研究。
17、1920年,Schack August Steenberg Krogh(丹麦)。
发现毛细血管运动的调节机制。

历年诺贝尔生理医学奖获奖者:1901年,(德国)。
利用疗法医治。
1902年,Ronald Ross(英国)。
关于的研究。
1903年,(丹麦)。
利用光辐射医治。
1904年,(俄国)。
在方面,提出了闻名的和信号学说。
1905年,R.(德国)。
关于方面的研究和发觉。
1906年,C.(意大利),桑地牙哥·拉蒙(Santiago Ramón y Cajal,西班牙)。
关于结构的研究。
1907年,Charles Louis Alphonse Laveran(法国),发此刻引发疾病中的作用。
1908年,Ilya Ilyich Mechnikov(俄国),(德国)。
关于方面的研究。
1909年,(Emil Theodor Kocher)(瑞士)。
关于生理学,病理学和外科学方面的研究。
1910年,艾布瑞契·(Albrecht Kossel)(德国)。
关于细胞化学尤其是和方面的研究。
1911年,(瑞典)。
关于眼睛学方面的研究。
1912年,Alexis Carrel(法国)。
关于血管缝合和血管和方面的研究。
1913年,Charles Robert Richet(法国)。
关于的研究。
1914年,Robert Bárány(奥地利)。
关于装置生理学及病理学方面的研究。
1915年-1918年,未颁奖,奖金划拨到生理医学奖专门的基金上。
1919年,Jules Bordet(比利时)。
关于免疫方面的研究。
1920年,Schack August Steenberg Krogh(丹麦)。
发觉运动的调剂机制。
1921年未颁奖,奖金划拨到生理医学奖专门的基金上。
1922年,Archibald Vivian Hill(英国),关于肌肉发烧方面的研究;Otto Fritz Meyerhof (德国),发觉肌肉中耗氧与乳酸代谢之间相关性。
1923年,(Frederick Grant Banting)(加拿大)、约翰·詹姆斯·理查德·麦克劳德(Macleod, John James Rickard (muh KLOWD)(苏格兰)。

历年诺贝尔生理医学奖获奖者:1901年,埃米尔·阿道夫·冯·贝林(德国)。
利用血清疗法治疗白喉。
1902年,Ronald Ross(英国)。
关于疟疾的研究。
1903年,尼尔斯·吕贝里·芬森(丹麦)。
利用光辐射治疗狼疮。
1904年,巴甫洛夫(俄国)。
在神经生理学方面,提出了著名的条件反射和信号学说。
1905年,R.柯赫(德国)。
关于结核方面的研究和发现。
1906年,C.高尔基(意大利),桑地牙哥·拉蒙卡哈(Santiago Ramón y Cajal,西班牙)。
关于神经系统结构的研究。
1907年,Charles Louis Alphonse Laveran(法国),发现原生动物在引起疾病中的作用。
1908年,Ilya Ilyich Mechnikov(俄国),保罗·埃尔利希(德国)。
关于免疫方面的研究。
1909年,埃米尔·特奥多尔·科赫尔(Emil Theodor Kocher)(瑞士)。
关于甲状腺生理学,病理学和外科学方面的研究。
1910年,艾布瑞契·科塞尔(Albrecht Kossel)(德国)。
关于细胞化学尤其是蛋白质和核酸方面的研究。
1911年,Allvar Gullstrand(瑞典)。
关于眼睛屈光学方面的研究。
1912年,Alexis Carrel(法国)。
关于血管缝合以及血管和器官移植方面的研究。
1913年,Charles Robert Richet(法国)。
关于过敏反应的研究。
1914年,Robert Bárány(奥地利)。
关于内耳前庭装置生理学及病理学方面的研究。
1915年-1918年,未颁奖,奖金划拨到生理医学奖专门的基金上。
1919年,Jules Bordet(比利时)。
关于免疫方面的研究。
1920年,Schack August Steenberg Krogh(丹麦)。
细胞周期循环的最主要的任务是将其基因组DNA在DNA合成期(S期)完整地复制成两份拷贝,而后在分裂期(M期)将这两份拷贝正确无误地分配给两个子代细胞.如果在这一过程中产生错误,又得不到及时纠正,那么将导致基因组的不稳定和变异.对单细胞生物,其后果是导致细胞增殖能力下降或细胞死亡;对多细胞生物则通常是引起肿瘤.在长期的进化过程中,细胞发展出了一套保证细胞周期中DNA复制和染色体分配质量的检查机制,通常称为细胞周期检验点(checkpoint).这是一类负反馈调节机制.当细胞周期进程中出现异常事件,如DNA损伤或DNA复制受阻时,这类调节机制就被激活,及时地中断细胞周期的运行.待细胞修复或排除了故障后,细胞周期才能恢复运转.根据“质量控制”的内容,可将细胞周期检查点分为三种:第一种负责查看DNA有无损伤,称为DNA损伤检查点;第二种负责DNA复制的进度,称为DNA复制检查点;第三类是管理染色体的正确分配与否,称为纺锤体组装检查点,因为染色体的分配主要依赖于纺锤体的作用.从检查点的工作方式来看,又可把检查点分为三个部分:第一个部分是探测器或传感器,它负责检查质量问题;第二个部分是信号传感器,它把“出了质量问题”的信号传递给第三部分———效应器,由效应器去中断细胞周期进程并开动修复机制.细胞周期的控制细胞周期调控是在各期的检验点上进行的.细胞周期中存在两个重要的检查点:一个位于G1/S交界处,在酵母菌中称为启动子,而在哺乳类动物细胞中称为限制点(R点),只要G1时相内的正调节因子累积达到一定程度,周期越过R 点,以后细胞就不再依赖于细胞外促生长因子而顺序完成整个细胞周期,可见R 点是影响细胞周期的最关键点;另一个位于G2/M交界处,它在染色体分开和细胞分裂前保证染色体均分的精确性.目前人们发现的细胞周期调控因子颇多,归纳起来可分三大类:周期素、周期素依赖性蛋白激酶、CDK抑制蛋白(CKIs).CDKs是细胞周期调控的中心环节,CDKs 的调节作用是通过使其底物(pRb及其相关蛋白p107、p130)的磷酸化来实现的;周期素是CDKs的正调节因子,能激活CDK活性;CKIs是负调节因子,可使CDK失活.下面主要从CDKs及其底物、周期素两方面阐述细胞周期调控因子在细胞周期调节中的作用.CDKs及其底物目前已发现的CDK有七种,分别命名为CDC2(CDK1)、CDK2~CDK7,它们在基因序列上的同源性超过40%.CDK的活性受细胞内外多种信号严密控制,活化的CDK包含催化亚单位(CDK)和一个正调亚单位(周期素),前者还需一个保守的Thr (苏氨酸)残基上的磷酸化和Tyr(酪氨酸)残基上的去磷酸化.CDK激活机制主要有:①CDK活化激酶(CDK-activated kinases,CAK)的作用.CAK自身也是CDK,它由一个催化亚单位CDK7/MO15和一个正调亚单位周期素H组成.CAK使CDK2的Thr161、CDK2的Thr160及CDK7的Thr170位点磷酸化而活化它们,CAK还能活化其他绝大多数CDK-周期素复合物.②周期素的扩增或过表达.③CKIs的突变、缺失或低表达.CDK失活机制有:Thr残基上的去磷酸化和Tyr残基上的磷酸化、与周期素s分离、同CKIs结合等途径.CDK被激活后,使其底物发生磷酸化.未磷酸化的pRb能够结合转录因子E2F,并抑制E2F启动S期相关基因的转录.磷酸化的pRb或pRb与某些转化蛋白如SV40的T抗原、腺病毒E1A蛋白、乳头瘤病毒E7蛋白结合,都能导致E2F的释放.游离的E2F与一种蛋白因子DP-1共同形成异源二聚体,能结合DNA的特定位点,正向激活S期相关基因转录,周期越过R点,细胞即从G1期进入S期.E2F分为五种类型.其中E2F1,E2F2,E2F3直接受pRb控制,E2F4和E 2F5主要受pRb相关蛋白p107、p130控制.p130和pRb在磷酸化调节方式上相同,但p130磷酸化的时间在pRb之前,因此认为p130的失活要早于pRb.目前,对p107了解尚少,但已知其和p130功能相似.周期素目前已发现的周期素有八种,分别命名为周期素A~H.根据细胞周期的时相,分为G1、G2、S和M时相以及作用尚未明确的周期素.周期素的过度表达可以激活CDK活性,造成细胞周期调节失控,导致肿瘤的形成,因此也称之为癌蛋白.G1时相周期素包括周期素C、D、E型,其中后两型是主要的.周期素D至少有D1、D2、D3三个亚型.目前周期素D1是研究的热点.周期素D1基因(又称CCND1、BCL-1、PRAD1或D11S128基因)编码的周期素D1由295个氨基酸组成.周期素D2基因(又称CCND2、vin-1基因)编码的周期素D2由290个氨基酸组成.周期素D3基因(又称CCND3基因)编码的周期素D3由292个氨基酸组成.三型周期素D氨基酸序列同源性达53.1%~63.5%,它们的C末端有一个富含脯氨酸(P)、谷氨酸(E)、天冬氨酸、丝氨酸(S)和苏氨酸(T)的残基序列,称之为PEST序列,与蛋白质的降解有关;它们的N末端有一个与某些转化蛋白如SV40的T抗原、腺病毒E1A蛋白、乳头瘤病毒E7蛋白共同的序列:亮氨酸Leu-X-半胱氨酸Cys-X-谷氨酸Glu,此序列同样可与pRb及pRb相关蛋白结合,随后在CDK4/CDK6的作用下,使pRb磷酸化,从而使细胞从G1时相进入S时相.一些生长因子(growth factors,GFs)可诱导它们的表达,一旦去除GFs,周期素D1的水平会迅速下降.因此,称它为生长因子感受器(growth factor sensors).在G1时相,给正常的成纤维细胞显微注射抗周期素D1抗体,能够阻止细胞进入S时相,但在接近G1/S交界处时则注射无效,这一结果表明周期素D1在G1的中晚期发挥着重要作用.周期素E在G1时相的表达晚于周期素D,在G1/S交界处达到高峰,进入S时相后逐渐下降,给细胞显微注射抗周期素E抗体,可阻止细胞进入S时相,因此被认为是调节G1/S时相转换的必需蛋白.细胞进入S时相后,周期素E降解,和它结合的CDK2被释放出来和周期素A结合.S时相周期素包括周期素A.它是周期素中最早被发现的,在G1时相晚期于周期素E之后表达,但直到S时相与CDK2结合后才被激活.给细胞显微注射抗周期素A抗体或反义周期素A能抑制细胞DNA的合成,表明周期素A在S时相有重要作用.另外,它在G2时相与M时相和CDC2(CDK1)结合,可能与有丝分裂有关.因此,周期素A也被称为M时相周期素.M时相周期素包括周期素A和B.周期素B又分为B1、B2、B3三种亚型, 它在S时相晚期和G2时相表达,且只和CDC2(CDK1)结合,在G2/M转换中发挥重要作用.其他周期素包括周期素F、G、H.它们的功能目前尚不完全清楚.对于所有的真核有机体,细胞周期的不同时相的相互协调是非常必要的.各时相要按正确的顺序进行;一个时相完成了,下一个时相才能开始.如果这种相互协调发生错误则会导致染色体的变异,染色体或染色体的一部分将丢失、重排或在两个子细胞之间不平等的分配.这种类型的染色体变异在癌细胞中经常可见.在生物和医学领域,理解如何控制细胞周期具有核心的作用.三位诺贝尔奖获得者在分子水平上认识了细胞周期中是如何从一时相转到下一时相.应用前景细胞周期调控机制将会在生物学、医学等诸多领域中得到广泛应用。
1901年,E . A . V . 贝林(德国人)从事有关白喉血清疗法的研究1902年,R.罗斯(英国人)从事有关疟疾的研究1903年,N.R.芬森(丹麦人)发现利用光辐射治疗狼疮1904年,I.P.巴甫洛夫(俄国人)从事有关消化系统生理学方面的研究1905年,R.柯赫(德国人)从事有关结核的研究1906年,C.戈尔季(意大利人)、S.拉蒙–卡哈尔(西班牙人)从事有关神经系统精细结构的研究1907年C.L.A.拉韦朗(法国人)发现并阐明了原生动物在引起疾病中的作用1908年P.埃利希(德国人)、E.梅奇尼科夫(俄国人)从事有关免疫力方面的研究1909年E.T.科歇尔(瑞士人)从事有关甲状腺的生理学、病理学以及外科学上的研究1910年A.科塞尔(德国人)从事有关蛋白质、核酸方面的研究1911年A.古尔斯特兰德(瑞典人)从事有关眼睛屈光学方面的研究1912年A.卡雷尔(法国人)从事有关血管缝合以及脏器移植方面的研究1913年C.R.里谢(法国人)从事有关抗原过敏的研究1914年R.巴拉尼(奥地利人)从事有关内耳前庭装置生理学与病理学方面的研究1915年——1918年未颁奖1919年J . 博尔德特(比利时人)作出了有关免疫方面的一系列发现1920年S.A.S.克劳(丹麦人)发现了有关体液和神经因素对毛细血管运动机理的调节1921年未颁奖1922年A.V.希尔(英国人)从事有关肌肉能量代谢和物质代谢问题的研究;迈尔霍夫(德国人)从事有关肌肉中氧消耗和乳酸代谢问题的研究1923年F.G.班廷(加拿大),J.J.R.麦克劳德(加拿大人)发现胰岛素1924年W.爱因托文(荷兰人)发现心电图机理1925年未颁奖1926年J.A.G.菲比格(丹麦人)发现菲比格氏鼠癌(鼠实验性胃癌)1927年J.瓦格纳–姚雷格(奥地利人)发现治疗麻痹的发热疗法1928年C.J.H.尼科尔(法国人)从事有关斑疹伤寒的研究1929年C.艾克曼(荷兰人)发现可以抗神经炎的维生素;F.G.霍普金斯(英国人)发现维生素B1缺乏病并从事关于抗神经炎药物的化学研究1930年K.兰德斯坦纳(美籍奥地利人)发现血型1931年O.H.瓦尔堡(德国人)发现呼吸酶的性质和作用方式1932年C.S.谢林顿、E.D.艾德里安(英国人)发现神经细胞活动的机制1933年T.H.摩尔根(美国人)发现染色体的遗传机制,创立染色体遗传理论1934年G.R.迈诺特、W.P.墨菲、G.H.惠普尔(美国人)发现贫血病的肝脏疗法1935年H.施佩曼(德国人)发现胚胎发育中背唇的诱导作用1936年H.H.戴尔(英国人)、O.勒韦(美籍德国人)发现神经冲动的化学传递1937年A.森特–焦尔季(匈牙利人)发现肌肉收缩原理1938年C.海曼斯(比利时人)发现呼吸调节中颈动脉窦和主动脉的机理1939年G.多马克(德国人)研究和发现磺胺药1940年——1942年没有颁奖1943年C.P.H.达姆(丹麦人)发现维生素K;E.A.多伊西(美国人)发现维生素K的化学性质1944年J.厄兰格、H.S.加塞(美国人)从事有关神经纤维机制的研究1945年A.弗莱明、E.B.钱恩、H.W.弗洛里(英国人)发现表霉素以及表霉素对传染病的治疗效果1946年H.J.马勒(美国人)发现用X 射线可以使基因人工诱变1947年C.F. 科里、G.T.科里(美国人)发现糖代谢中的酶促反应;B.A.何赛(阿根廷人)发现脑下垂体前叶激素对糖代谢的作用1948年P.H.米勒(瑞士人)发现并合成了高效有机杀虫剂DDT1949年W.R.赫斯(瑞士人)发现动物间脑的下丘脑对内脏的调节功能1950年E.C.肯德尔、P.S.亨奇(美国人)T.赖希施泰因(瑞士人)发现肾上腺皮质激素及其结构和生物效应1951年M.蒂勒(南非人)发现黄热病疫苗1952年S.A.瓦克斯曼(美国人)发现链霉素1953年F.A.李普曼(英国人)发现高能磷酸结合在代谢中的重要性,发现辅酶A;H.A.克雷布斯(英国人)发现克雷布斯循环(三羧酸循环)1954年J.F.恩德斯、T.H.韦勒、F.C.罗宾斯(美国人)研究脊髓灰质炎病毒的组织培养与组织技术的应用1955年A.H.西奥雷尔(瑞典人)从事过氧化酶的研究1956年A.F.库南德、D.W.理查兹(美国人)、W.福斯曼(德国人)开发了心脏导管术1957年D.博维特(意籍瑞士人)从事合成类箭毒化合物的研究1958年G.W.比德乐、E.L.塔特姆(美国人)发现一切生物体内的生化反应都是由基因逐步控制的;J.莱德伯格(美国人)从事基因重组以及细菌遗传物质方面的研究1959年S.奥乔亚、A.科恩伯格(美国人)从事合成RNA和DNA的研究1960年F.M.伯内特(澳大利亚人)、P.B.梅达沃(英国人)证实了获得性免疫耐受性1961年G.V.贝凯西(美国人)确立“行波学说”发现耳蜗感音的物理机制1962年J.D.沃森(美国人)、F.H.C.克里克、M.H.F.威尔金斯(英国人)发现核酸的分子结构及其对住处传递的重要性1963年J.C.艾克尔斯(澳大利亚人)、A.L.霍金奇、A.F.赫克斯利(英国人)发现与神经的兴奋和抑制有关的离子机构1964年K.E.布洛赫(美国人)、F.吕南(德国人)从事有关胆固醇和脂肪酸生物合成方面的研究1965年F.雅各布、J.L.莫诺、A.M.雷沃夫(法国人)研究有关酶和细菌合成中的遗传调节机构1966年F.P. 劳斯(美国人)发现肿瘤诱导病毒;C.B.哈金斯(美国人)发现内分泌对于癌的干扰作用1967年R.A.格拉尼特(瑞典人)、H.K.哈特兰、G.沃尔德(美国人)发现眼睛的化学及重量视觉过程1968年R.W.霍利、H.G.霍拉纳、M.W.尼伦伯格(美国人)研究遗传信息的破译及其在蛋白质合成中的作用1969年M.德尔布吕克、A.D.赫尔、S.E.卢里亚(美国人)发现病毒的复制机制和遗传结构1970年B.卡茨(英国人)、U.S.V.奥伊勒(瑞典人)J.阿克塞尔罗行(美国人)发现神经末梢部位的传递物质以及该物质的贮藏、释放、受抑制机理1971年E.W.萨瑟兰(美国人)发现激素的作用机理1972年G.M.埃德尔曼(美国人)、R.R.波特(英国人)从事抗体的化学结构和机能的研究1973年K.V.弗里施、K.洛伦滋(奥地利人)、N.廷伯根(英国人)发现个体及社会性行为模式(比较行为动物学)1974年A.克劳德、C.R.德·迪夫(比利时人)、G.E.帕拉德(美国人)从事细胞结构和机能的研究1975年D.巴尔摩、H.M.特明(美国人)、R.杜尔贝科(美国人)从事肿瘤病毒的研究1976年B.S.丰卢姆伯格(美国人)发现澳大利亚抗原;D.C.盖达塞克(美国人)从事慢性病毒感染症的研究1977年R.C.L.吉尔曼、A.V.沙里(美国人)发现下丘脑激素;R.S.雅洛(美国人)开发放射免疫分析法1978年W.阿尔伯(瑞士人)、H.O.史密斯、D.内森斯(美国人)发现限制性内切酶以及在分子遗传学方面的应用1979年A.M.科马克(美国人)、G.N.蒙斯菲尔德(英国人)开始了用电子计算机操纵的X 射线断层扫描仪(简称扫描仪)1980年B.贝纳塞拉夫、G.D.斯内尔(美国人)、J.多塞(法国人)从事细胞表面调节免疫反应的遗传结构的研究1981年R.W.斯佩里(美国人)从事大脑半球职能分工的研究;D.H.休伯尔(美国人)、T.N.威塞尔(瑞典人)从事视觉系统的信息加工研究1982年S.K.贝里斯德伦、B.I.萨米埃尔松(瑞典人)J.R.范恩(英国人)发现前列腺素,并从事这方面的研究1983年B.麦克林托克(美国人)发现移动的基因1984年N.K.杰尼(丹麦人)、G.J.F.克勒(德国人)、C.米尔斯坦(英国人)确立有免疫抑制机理的理论,研制出了单克隆抗体1985年M.S.布朗、J.L.戈德斯坦(美国人)从事胆固醇代谢及与此有关的疾病的研究1986年R.L.蒙塔尔西尼(意大利人)、S.科恩(美国人)发现神经生长因子以及上皮细胞生长因子1987年利根川进(日本人)阐明与抗体生成有关的遗传性原理1988年J.W.布莱克(英国人)、G.B.埃利昂、G.H.希钦斯(美国人)对药物研究原理作出重要贡献1989年J.M.毕晓普、H.E.瓦慕斯(美国人)发现了动物肿瘤病毒的致癌基因源出于细胞基因,即所谓原癌基因1990年J.E.默里、E.D.托马斯(美国人)从事对人类器官移植、细胞移植技术和研究1991年E.内尔、B.萨克曼(德国人)发明了膜片钳技术1992年E.H.费希尔、E.G.克雷布斯(美国人)发现蛋白质可逆磷酸化作用1993年P.A.夏普、R.J.罗伯茨(美国人)发现断裂基因1994年A.G.吉尔曼、M.罗德贝尔(美国人)发现G 蛋白及其在细胞中转导信息的作用1995年E.B.刘易斯、E.F.维绍斯(美国人)、C.N.福尔哈德(德国人)发现了控制早期胚胎发育的重要遗传机理,利用果蝇作为实验系统,发现了同样适用于高等增有机体(包括人)的遗传机理1996年P.C.多尔蒂(澳大利亚人)、R.M.青克纳格尔(瑞士人)发现细胞的中介免疫保护特征1997年S.B.普鲁西纳(美国人)发现了一种全新的蛋白致病因子——朊蛋白(PRION)并在其致病机理的研究方面做出了杰出贡献1998年R.F.福尔荷格特、L.J.依格那罗和F.穆莱德发现一氧化一氮在心血管系统中作为信号分子1999年Gunter Blobel发现控制细胞运输和定位的内在信号蛋白质2000年阿尔维德·卡尔松(瑞典人)、保罗·格林加德(美国人)、埃里克·坎德尔(奥地利人)在“人类脑神经细胞间信号的相互传递”方面获得的重要发现。
近20年诺贝尔生理和医学奖得主一览85年,美国纽约医生Michael S. Brown和南卡医生Joseph L.Geldstein 胆固醇的代谢调控86年意大利进化学家Rita Levi Montalcini美国斯坦福Stanley Cohen 发现生长因子87年美国麻省理工学院教授Susumu Tonegawa发现抗体多样性的遗传学原理88年英国James W. Black美国G. B. Elion & G. H. Hitchings关于药物研发相关原理的研究89年美国癌症中心Harold E.Varmus和加州理工学院Michael Bishop 发现逆转录病毒原癌基因(oncogene)90年美国E. Donnall Thomas和Joseph E. Murray 发明一种治疗疾病的新方法即细胞和器官移植91年德国Bert Sakmann和Erwin Neher发现细胞膜上离子通道的功能92年美国Edmond H.Fischer和Edwin G. Krebs蛋白质可逆磷酸化作为一种生物调节机制93年美国麻省Richard J. Roberts和Phillip A. Sharp Eukaryotic genes are split and spliced after transcription 94年美国Alfred G. Gilman和Martin RodbellG-蛋白及其在细胞信号传导中的作用95年美国Edward B. Lewis和Eric F. Wieschaus 德国Christiane Nüsslein-Volhard早期胚胎发育中的遗传调控机理96年澳大利亚Peter C. Doherty瑞士Rolf M. Zinkernagel 细胞中介的免疫保护特性97年美国Stanley B. Prusiner新的蛋白致病因子朊蛋白98年美国Robert F.Furchgott,Louis J. Ignarro和Ferid Murad 一氧化氮在心脏血管中的信号传递功能99年美国纽约洛克菲勒大学Günter Blobel蛋白质具有内在信号元件控制其胞内运输和定位00年瑞典Arvid Carlsson和美国Paul Greengard,Eric Kandel 神经系统信号传导方面的研究01年美国Leland.H.Hartwell和英国Tim Hunt,Sir Paul Nurse 细胞周期中的关键调节因子02年英国S Brenner,JE Sulston和美国H. R. Horvitz 器官发育和细胞程序性死亡的遗传调控机理03年 Paul C. Lauterbur和Sir Peter Mansfield 关于核磁共振成像的研究04年 Richard Axel和Linda B. Buck嗅觉的研究。
细胞周期,癌症与诺贝尔奖2001年的诺贝尔生理医学奖授予了3位研究细胞周期并取得卓越成就的科学家,他们的工作使我们对细胞增殖及其与癌症的关系有了更深刻的理解,从而为我们找到治愈癌症之路指明方向。
但是,他们的工作究竟有多重要,而有那么多的科学家在这一领域中工作,为什么独独是这3位而不是其它人得到这个全世界科学家都梦寐以求的这个荣誉呢?我们就来看看他们到底都做了些什么。
2001年诺贝尔生理医学奖获奖者(从左至右)Leland Hartwell、Tim Hunt和Paul Nurse。
一、细胞周期所谓细胞周期(cell cycle)是指连续分裂细胞从一次有丝分裂结束到下一次有丝分裂结束所经历的整个过程。
在这个过程中,细胞遗传物质复制并加倍,且在分裂结束时平均分配到两个子细胞中去。
细胞周期又可以分为间期(interphase)和有丝分裂期(M phase)。
从一次有丝分裂结束到下一次有丝分裂开始的时期就是间期。
这一时期,在光学显微镜下看不到细胞有明显的变化,但此时期的细胞内却正在进行一系列的生化活动,主要的活动围绕制造完全相同的又一套遗传物质展开。
这一期以DNA合成为标志,又分为G1期,S期和G2期。
而在光学显微镜下可以看到的只是M期,经过分裂期,加倍的染色体和其他细胞组分被平均分配到两个完全一样的子细胞中。
换句话,通过分裂,形成了一个新细胞。
事实上早在1841年,时任职于柏林大学的波兰神经内科学家和生物学家罗伯特·里麦克(Robert Remak,1815-1865)就报道了细胞分裂现象,并得出结论,细胞分裂是细胞增殖的方式也是机体生长发育的“根本动力”;更有意义的是,他在此时就已经认为肿瘤组织中细胞的形成机制“几乎与正常动物组织相同”。
不过,由于受观察手段得的限制,人们还不可能了解到有丝分裂间期中发生的生化事件,而又由于在显微镜下染色体的变化是如此规律,因此,认为细胞的增殖活动主要发生在形态变化明显的有丝分裂期就不难理解了。
直到1953年,Howard和Pelc才发现蚕豆根尖细胞分裂中遗传物质DNA的复制发生于静止期中的一个时期,这一时期与有丝分裂期在时间上存在前后两个间隙。
由此,他们第一次明确的提出了细胞周期的概念,并将细胞周期划分为上述的4个时期,其中的S期即是DNA合成的时期。
细胞在细胞周期中续惯的经过G1→S→G2→M而完成增殖。
1957年,3THdR放射自显影技术引入细胞周期的研究后,证明在动植物细胞中细胞周期存在普遍性,从而使对细胞增殖的研究引入到一个全新的时期,因此,细胞周期概念的提出成为20世纪细胞生物学的一个划时代成就。
细胞周期示意图G1期又称为DNA合成前期,此期DNA的合成还没有开始,但是这一时期中所进行的RNA 和蛋白质合成却是DNA复制所必须的,而且,S期中DNA合成的启动也在这一时期受到调控。
因此这一时期可以看作为DNA合成的准备时期。
S期中,DNA进行合成,同时,染色体形成所必需的组蛋白,非组蛋白等物质也在此期合成。
G2期主要合成一些与有丝分裂中新细胞形成所必需的物质,如作为细胞骨架重要成分的微管蛋白等。
M期中,细胞染色体形成并发生细胞分裂,新细胞在此期形成。
二、细胞周期调控细胞周期能够严格按照G1→S→G2→M的顺序运转是与相关调控基因的有序表达分不开(cell cycle checkpoint)的。
Leland Hartwell在20世纪70年代提出了“细胞周期检验点”概念。
这是作用于细胞周期转换过程的关键调控通路,从分子水平看就是一类调控基因及其表达产物对细胞是否以及如何分裂所进行的精细调节。
这些基因产物接受两种不同的信号,并作出反应。
一类信号是所谓“细胞内在信息”,这类信息反映细胞周期是否按照正确顺序进行,即细胞周期中的下一个时期是否在前一时期结束后才开始,换句话说,只有在完成前期所有的准备工作后,细胞分裂才可以开始,并且所有前期准备工作也只有在严格按部就班的情况下才可以完成。
另一类信号称为“细胞外在信息”,是指对细胞分裂与否至为重要的环境信号,只有当细胞所处的环境需要更多细胞时,细胞的分裂才是合适的,不和时宜的细胞分裂带来的就是肿瘤,因此细胞必须通过检验点机制识别这些信号,以避免不恰当的进入细胞周期。
Hartwell不仅提出了假设,他还通过对突变酵母细胞——在细胞周期研究中成功的引入并构建酵母这一重要的模型也是Hartwell的首创——的研究,证实了此类检验点的存在,其中尤值一提的是他对酵母细胞DNA合成开始前启动点(start)的研究,通过这一研究他进一步提出了细胞分裂基因的概念,即cdc(cell division cycle)基因。
此后,他与其同事并相继发现一系列此类基因,并进而指出,这些基因的表达受制于细胞中有关蛋白质和酶,通过在恰当的时间和位点出现这些调节蛋白和酶,使cdc基因激活,从而保证细胞周期中的关键事件高度准确的完成。
在他工作的基础上,为更好描述细胞分裂的这种复杂调控机制,有研究者提出了所谓“细胞周期中心控制体系”的概念,该理论综合了生理学和遗传学有关细胞周期的“时钟理论”和“骨牌理论”,前者认为细胞周期的控制体系类似钟摆,使细胞不断在有丝分裂期和间期来回循环;而后者则认为,细胞周期中的序贯事件在时间上是前后依赖的,即后一事件的启动依赖前一事件的完成,类似多米诺骨牌现象而得名。
以后的研究发现,这两种理论是对细胞周期中现象在不同层面上的认识,而真核细胞分裂调控的本质是一系列细胞内信号传导通路中分子的磷酸化和去磷酸化过程,故此,将其综合为前述细胞周期中心控制体系理论。
可以借用一个比喻来理解这一控制体系:“中心控制体系对细胞周期的调控作用类似于全自动洗衣机中的控制器,当它转到细胞周期中某特定点时,就启动该点的细胞进行一系列次级生化事件(如DNA复制、有丝分裂等);而且,它也接受正在进行的事件中传来的反馈信号而决定是否应启动下一个事件,从而防止在前一过程未完成时就开始下一过程,若失去接受反馈信号的功能,则将发生不正常的细胞分裂。
因此,中心控制体系也起到类似闸门样的关卡作用,控制着细胞周期中各事件的过渡。
”(引自《真核基因表达调控》p151,高等教育出版社-斯普林格出版社,1999,北京)中心控制体系的分子基础由一系列与真核细胞分裂有关的基因表达产物组成,其中包括钙调素(calmodulin)、周期蛋白(cyclin)家族及周期蛋白依赖性蛋白激酶(cyclin-dependent kinase,CDK)等。
又尤以后二者广为人所关注。
对细胞分裂调控的研究早在20世纪80年代即已经取得进展,Tim Hunt在对海胆和海蛤胚胎发育早期卵裂细胞的研究中发现,随细胞周期的进行,一个分子量约45-60KD的蛋白质随之发生合成剧烈增高又突然下降的震荡,它在间期开始合成而在G2→M期达到高峰,又在M期结束后陡然下降,下一轮间期又重新合成,故此将其命名为周期蛋白(cyclin),进一步的研究发现这是一类蛋白质,根据结构的差异将其予以分类,并视做一个蛋白质家族。
这一蛋白质可以诱导未成熟细胞进入M期,并发现在M期唯一需要合成的蛋白质就是这个周期蛋白,从而肯定这是一个起关键作用的调节蛋白。
在此之前的70年代中,人们已经在Hela 细胞和非洲爪蟾卵母细胞中发现存在可诱导有丝分裂和减数分裂的两种因子,并由于这二者在性质上十分相似,故统称之为“有丝分裂或减数分裂促进因子”(MPF),但在10年间,都未能将之纯化。
1988年,终于实现了MPF的纯化,发现这是由分子量分别为32KD和45KD 的二种蛋白质组成的复合物,其中45KD蛋白的测序发现,原来它就是周期蛋白。
而1990年,Paul Nurse发现MPF中作为丝氨酸/苏氨酸蛋白激酶的32KD产物在细胞从G2期向M期的过渡中,似乎起了决定性的作用,并进一步发现它与在80年代分离的裂殖酵母细胞分裂周期基因cdc2编码产物p34cdc2具有同源性,换句话,这二者又是同一个东西。
并且,Nurse 还发现,对于p34cdc2的活性而言,周期蛋白是必需的,所以将其命名为周期蛋白依赖性蛋白激酶(CDK)。
由此,Nurse在1990年提出:从酵母到无脊椎动物一直到人类,其所有真核细胞中存在一个共同的M期启动调节机制——“M期启动调节的普遍机制”。
至此,一整套认识细胞分裂增殖的逻辑和事实框架已经得到初步建立。
回顾一下细胞周期发现的历程,不难发现,Hartwell作为微生物学家,最先找到了用于研究真核细胞周期的合适模型——酵母细胞,同时他率先发现了cdc基因,为其后20年间细胞周期研究指明了方向——虽然他的这一工作的意义在10年后才真正显得尤为重要——并通过大量的研究为细胞周期理论的大厦奠定了基础。
而Tim Hunt和Paul Nurse在完全不同的方向上进行的工作揭示了细胞周期中的关键分子及其作用,最终这些工作与Hartwell的研究相整和,使人们得到了有关细胞分裂的主要概念框架,从而为与真核细胞增殖及其基因调控机制相关的研究提供了重要的切入点。
并且,也正是这些工作促成了对细胞增殖网络调节的精细认识。
今天回头看去,这三位科学家的成就的确具有划时代意义。
三、细胞周期与癌症癌症细胞本质上也来源于机体正常细胞,但由于在细胞分裂的不同阶段受各种因素的影响,导致其出现分化和增殖的异常,从而危及机体的整体安全。
目前对癌症的认识还不够清楚,但是,基于对细胞周期的了解,已经明确,细胞周期的正常运转有赖于细胞引擎分子——Cdks-cyclin、癌基因、抑癌基因和细胞周期调节蛋白彼此相互作用而构成的一个调节网络。
随着对这一调节网络的更深入了解,必将能对癌症的发病机制有透彻了解。
正是在这一意义上,细胞周期的研究使我们看到了攻克癌症的曙光。
由于这些成就不仅对于普通生物学的基础研究具有重要意义,也由于这些工作本身及其对后继学科和应用科学具有重要指导意义,因此,2001年的诺贝尔生理医学颁授给了前述3位科学家——Leland Hartwell、Tim Hunt与Paul Nurse。
选自:三思科学网站。