腺相关病毒知识(汇编)
- 格式:doc
- 大小:47.50 KB
- 文档页数:10

腺病毒知识点总结1. 腺病毒的分类腺病毒目前已知包括了六个属,分别为腺病毒属(Mastadenovirus)、阿卡洛病毒属(Atadenovirus)、索哈病毒属(Siadenovirus)等。
其中腺病毒属包括高度致病性的人类腺病毒、犬腺病毒等。
2. 腺病毒的结构腺病毒的外壳由蛋白质构成,内含双链DNA。
典型的腺病毒粒子呈多面体或类似无规则聚合的形状,直径约70-90纳米,表面均匀分布有纤维状的突起。
腺病毒粒子内含有DNA、蛋白质和酶。
3. 腺病毒的感染机制腺病毒通过呼吸道、消化道、眼部粘膜等途径进行感染。
一旦腺病毒进入人体,通过与细胞表面相关受体结合,进入细胞内。
腺病毒感染后,宿主细胞会被利用来合成病毒的DNA 和蛋白质,在宿主细胞内进行繁殖和增殖,破坏并利用宿主细胞的资源。
4. 腺病毒的致病性腺病毒引起的疾病种类繁多,常见的包括呼吸道感染、结膜炎、胃肠道感染、脑膜炎等。
不同种类的腺病毒可引起不同的疾病。
例如,腺病毒D型和E型主要引起呼吸道感染,腺病毒F型主要引起眼部感染。
5. 腺病毒的预防措施目前,预防腺病毒感染的主要措施包括加强个人卫生、保持良好的生活习惯、接种疫苗等。
通过保持清洁卫生,避免接触患者体液,保持良好的生活习惯,如良好的饮食习惯、充足的睡眠等,有助于预防腺病毒感染。
此外,疫苗接种也是预防腺病毒感染的有效手段。
目前,已有一些疫苗用于预防特定种类的腺病毒感染。
总之,腺病毒是一类广泛存在的病原体,对人类和动植物造成了严重威胁。
了解腺病毒的分类、结构、感染机制、致病性和预防措施等知识,有助于人们更好地认识腺病毒,预防和控制腺病毒感染,保障公共健康。
希望通过本文的介绍,读者能够更全面地了解腺病毒及其相关知识。

医疗卫系统公开招聘考试临床医学专业知识真题汇编2016年(1)一、判断题1. 胰岛素是促进合成代谢、调节血糖浓度的主要激素。
正确答案:A[解析] 胰岛素是由胰岛分泌的一种激素,胰岛素是促进物质合成(江南博哥)代谢,维持血糖水平稳定的关键激素。
2. 低钾血症可引起限制性通气障碍。
正确答案:A[解析] 低钾造成呼吸肌麻痹,从而导致限制性通气障碍。
3. 所有缺氧病人都会出现发绀现象。
正确答案:B[解析] 紫绀是缺氧的表现,但缺氧的病人不一定都有紫绀,例如贫血引起的血液性缺氧可无紫绀,缺氧指组织得不到充足的氧或不能充分利用氧时,组织的代谢、功能,甚至形态结构发生异常变化的病理过程。
紫绀主要是由于血液中血红蛋白氧合不全、还原血红蛋白增高所致。
任何原因使毛细血管内还原血红蛋白绝对含量超过50g/L时,即可出现发绀。
紫绀是缺氧的表现,但缺氧并不一定都发绀。
4. 阿托品对胆绞痛和肾绞痛的疗效较好。
正确答案:B[解析] 阿托品适用于各种内脏绞痛,对胃肠绞痛,膀胱刺激症状如尿频、尿急等疗效较好,但对胆绞痛或肾绞痛疗效较差,常需与阿片类镇痛药合用。
5. 铁的吸收部位主要在空肠后段和回肠。
正确答案:B[解析] 铁的吸收部位主要是在十二指肠和空肠上端。
6. 非淋菌性尿道炎的病原体以沙眼衣原体和支原体为主。
正确答案:A[解析] 非淋菌性尿道炎是指由淋菌以外的其他病原体,主要是沙眼衣原体和支原体等引起的一种性传播疾病。
7. 肾癌从肾小管上皮细胞发生。
正确答案:A[解析] 肾癌是肾小管上皮细胞发生的实质性恶性肿瘤,外有假包膜。
肾癌局限在包膜内恶性度较小,穿透包膜后可经血液和淋巴结转移,淋巴转移最先到。
肾蒂淋巴结。
8. 颅脑外伤造成的原发性脑损伤包括脑挫裂伤、颅内血肿、脑水肿和脑肿胀等。
正确答案:B[解析] 原发性脑损伤是指伤后立即发生的病理性损害,包括脑震荡、脑挫裂伤。
继发性脑损伤是指在原发性脑损伤的基础上逐渐发展起来的病理改变,主要是颅内血肿和脑肿胀、脑水肿。

腺病毒腺病毒(adenovirus)是一种没有包膜的直径为70~90nm的颗粒,由252个壳粒呈廿面体排列构成。
每个壳粒的直径为7~9nm。
衣壳的蛋白质分子共价结合,可以出现双链DNA的环状结构。
1腺病毒简介体腺病毒已知有52种,分别命名为adl~ad52,研究得最详细是ad2。
腺病毒基因组转录产生mRNA,已知的转录单位至少有5个:EⅠ区位于病毒基因组左侧,可再分成EⅠA和EⅠB,与细胞转化有关;EⅡ区编码DNA结合蛋白,参与病毒的复制;EⅢ区编码出现在宿主细胞表面的一种糖蛋白;EⅣ区位于ad2基因组右端,受EⅡ区编码的DNA结合蛋白质调控;第5个转录单位在病毒感染中期合成ad2蛋白质Ⅳ。
腺病毒对啮齿类动物有致癌能力,或能转化体外培养的啮齿类动物细胞。
使细胞转化只需要腺病毒基因组的一部分,这些基因位于基因组的左端,约占整个基因组的7%~10%。
尽管腺病毒分布很广,但对人体不出现致癌性。
人体细胞是一类允许细胞(permissive cell),即这类细胞允许感染入侵的病毒在细胞内复制增殖,最后细胞裂解死亡而释放出大量子代病毒。
在体外培养的多种人体肿瘤细胞中均未查出腺病毒颗粒,但在人的1号染色体上有adl2的整合位点,这意味着人体细胞对于腺病毒也可能是非允许细胞,即这类细胞在病毒感染后,病毒不能在细胞内复制增殖,但可整合在受感染细胞的基因组内。
这些细胞被病毒转化,表型发生改变,且可在体外无限期地培养传代。
2结构及化学组成编辑腺病毒是一种无包膜的双链DNA病毒,基因组长约25-45kb,理论上可编码22-40个基因。
衣壳(capsid)呈规则的20面体结构,直径约80-110nm。
衣壳含有240个六联体(hexon)、12个五联体(penton)及12根纤毛(fiber),除此之外还有其他一些小蛋白,如VI、VIII、IX、IIIa和IVa2等。
六联体是形成病毒衣壳20个三角形面的主要蛋白,12个顶端是5个五联体亚单位和3个纤毛蛋白构成的复合物,12根纤毛以五联体蛋白为基底由衣壳表面伸出,纤毛顶端形成头节区(knob)。


腺病毒和腺相关病毒有哪些区别?腺病毒和腺相关病毒(AAV)是用于基因传递的两种不同类型的病毒载体。
这两个重组病毒系统都具有感染广泛宿主的能力,包括分裂和非分裂细胞,而无需与宿主基因组整合。
它们两之间有几个主要区别:包装能力、水平,基因表达的起效和持续时间以及免疫应答。
腺病毒简介腺病毒有约8.5千碱基的容量,具有高水平的蛋白质表达和瞬时基因表达。
表达的发作时间可早于感染后16-24小时。
腺病毒系统的局限性主要在于靶细胞的高免疫应答。
尽管如此,由于它们对大多数组织有高效转导作用,仍被广泛用于研究中。
腺病毒不能整合至宿主基因组,仅能瞬时表达。
它可以对分裂期及非分裂期细胞进行感染。
其包装片段至多8 kb,浓缩滴度至多为1011-1012TU/mL,片段越大,滴度越低,表达水平还是比较高的。
腺病毒优势宿主范围广:能够感染分裂期细胞与静止期细胞;对上皮细胞有高度嗜性,尤其适用于感染原代细胞、悬浮细胞等难转染细胞类型感染效率高:优于其他病毒载体和转染系统的高感染能力,能达到近100%的效率包装容量大:最高可插入7.5 kb的外源片段表达水平高:1-2d即开始高水平表达安全系数高:去除病毒蛋白编码序列,降低细胞毒性和免疫反应;双质粒转染系统,生物安全性高无整合风险:无插入致突变性腺相关病毒简介AAV的包装容量约为4.5千碱基,蛋白质表达水平相对较低,有长效基因表达的潜力。
AAV的趋向性也可以通过不同的血清型增强。
AAV的主要缺点是其对目的基因的包装容量较小,并且表达起效较晚(体外2-7天,体内3-21天),然而,该传递系统所产生的免疫应答水平却非常低。
腺相关病毒可以稳定整合至宿主基因组,并且可以在特定基因产物存在的条件下整合至人类19号染色体中的特定位点。
它能对分裂期及非分裂期细胞进行感染。
其包装片段至多8 kb,浓缩滴度至多为1011-1013TU/mL,片段越大,滴度越低。
腺相关病毒可以应用Tet-on等诱导表达系统,表达水平也是比较高的。
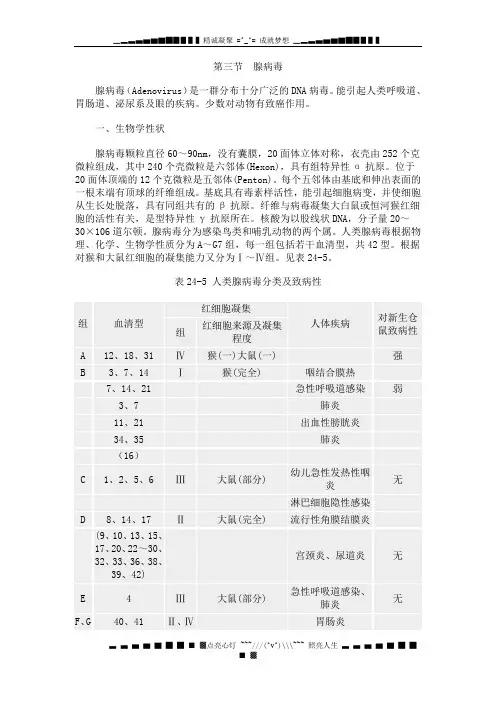
第三节腺病毒腺病毒(Adenovirus)是一群分布十分广泛的DNA病毒。
能引起人类呼吸道、胃肠道、泌尿系及眼的疾病。
少数对动物有致癌作用。
一、生物学性状腺病毒颗粒直径60~90nm,没有囊膜,20面体立体对称,衣壳由252个克微粒组成,其中240个壳微粒是六邻体(Hexon),具有组特异性α抗原。
位于20面体顶端的12个克微粒是五邻体(Penton)。
每个五邻体由基底和伸出表面的一根末端有顶球的纤维组成。
基底具有毒素样活性,能引起细胞病变,并使细胞从生长处脱落,具有同组共有的β抗原。
纤维与病毒凝集大白鼠或恒河猴红细胞的活性有关,是型特异性γ抗原所在。
核酸为以股线状DNA,分子量20~30×106道尔顿。
腺病毒分为感染鸟类和哺乳动物的两个属。
人类腺病毒根据物理、化学、生物学性质分为A~G7组,每一组包括若干血清型,共42型。
根据对猴和大鼠红细胞的凝集能力又分为Ⅰ~Ⅳ组。
见表24-5。
表24-5 人类腺病毒分类及致病性*括弧中型别的一般不致病人类腺病毒不能在鸡胚中增殖,上皮样人细胞系HeLa细胞和人胚原代细胞培养最敏感,能引起细胞肿胀、变圆、聚集成葡萄串状的典型细胞病变。
腺病毒对酸碱度及温度的耐受范围较宽,36℃7天病毒感染力无明显下降。
对脂溶剂和酶类均抵抗作用,但56℃30分钟可将其灭活。
二、致病性与免疫性人腺病毒中部分型别有致病性,最常的是1~7型。
幼儿急性上呼吸道感染约5%由腺病毒引起,成人感染很少发生于呼吸道。
腺病毒致病情况见表24-5。
传播以粪一口为主途径,也可通过呼吸道或污染物品传播。
病毒在咽、结膜尤其是小肠上皮细胞内增殖,偶尔波及其他脏器,隐性感染常见。
疾病一般为自限性,感染后可获得长期持续的型特异性免疫力,中和抗体损伤作用重要。
A、B组病毒在某些新生动物可诱发肿瘤,对人未发现致癌作用。
三、微生物学检查(一)病毒分离自急性期病人咽、直肠、结膜等处采取标本,迅速接种敏感细胞,根据特征性细胞突变及抗原性鉴定病毒。

什么是腺病毒
腺病毒是一类寄生于动物和人类的病毒,属于腺病毒科。
这种病毒在寄主细胞
内复制自身,引发一系列疾病。
腺病毒最常见的类型包括腺病毒5型、腺病毒7
型等,它们可以引发呼吸道感染、肠道感染和眼部感染等疾病。
腺病毒的特征
腺病毒是一种非常常见的病原体,易于传播。
它通过飞沫传播等途径感染人体,尤其容易在人口密集的地方传播。
腺病毒感染后,潜伏期较短,通常数天之内症状就会出现。
腺病毒感染的症状
腺病毒感染会引发不同的症状,主要表现为发热、咳嗽、流涕、喉咙痛、头痛、疲劳等。
严重的腺病毒感染还可能导致肺炎、腹泻等并发症。
如何预防腺病毒感染
为了预防腺病毒感染,我们可以采取以下措施:
1.注重个人卫生,勤洗手,尤其在接触食物、外出后要洗手。
2.避免和已感染腺病毒的人密切接触,尤其是在疫情流行期间。
3.保持室内空气流通,勤通风,保持室内清洁。
4.提高免疫力,多吃水果蔬菜,合理作息,增强体质。
腺病毒的治疗
目前尚无特效的抗病毒药物可以治疗腺病毒感染,治疗主要是对症支持疗法,
如饮食调理、适当的休息、多喝水等。
对于症状较重的患者,医生会根据具体情况采取相应的治疗措施。
综上所述,腺病毒是一种常见的病毒感染,预防措施尤为重要。
在日常生活中,注意个人卫生、保持良好的生活习惯,有助于降低感染的风险,有效控制腺病毒传播。


高考生物必背知识点汇编高考生物必背知识清单1.糖类、脂质、蛋白质和核酸共有的元素是C、H、O,除此之外,蛋白质中还含有N等元素,核酸中还含有N、P。
2.组成蛋白质的氨基酸约有20种,不同氨基酸理化性质差异的原因在于R基不同。
3.DNA和RNA在分子组成上的差异:DNA中含有脱氧核糖和胸腺嘧啶,而RNA中含有核糖和尿嘧啶。
4.DNA多样性的原因主要是碱基(脱氧核苷酸)的排列顺序不同;而蛋白质多样性的原因是组成蛋白质的氨基酸的种类、数目、排列顺序以及肽链的空间结构不同。
5.熟记实验中的颜色反应:蛋白质+双缩脲试剂→紫色; DNA +甲基绿染液→绿色;RNA +吡罗红(派洛宁)染液→红色; 还原糖+斐林试剂加热――→砖红色;脂肪+苏丹Ⅲ(Ⅳ)染液→橘黄色(红色); 线粒体+健那绿染液→蓝绿色。
6.乳糖和糖原只分布于动物细胞;蔗糖、麦芽糖、淀粉和纤维素只分布于植物细胞。
葡萄糖、核糖、脱氧核糖在动植物细胞中都有。
7.脂质主要包括脂肪、磷脂和固醇,其中固醇又包括胆固醇、性激素和维生素D 等。
8.脂肪的含氢量高于糖类,因此氧化分解时,耗O 2多,释放能量也多。
9.自由水/结合水的比值越大,生物新陈代谢越旺盛,但其抗逆性相对较小。
10.原核细胞没有核膜、核仁、染色体及除核糖体以外的细胞器。
11.各种生物膜都主要由脂质、蛋白质组成,细胞膜还含有少量糖类。
功能越复杂的膜中,蛋白质的种类和数量越多。
12.生物膜的结构特点是具有一定的流动性,功能特性是具有选择透过性。
13.生物膜系统包括细胞膜、核膜及具膜细胞器。
核糖体、中心体不是生物膜系统的组成成分。
14.内质网膜与核膜、细胞膜能直接转化,高尔基体膜与内质网膜、细胞膜通过囊泡发生间接转化。
15.根尖分生区细胞没有叶绿体和大液泡,低等植物细胞有中心体。
16.没有线粒体和叶绿体的细胞也可进行有氧呼吸和光合作用,如蓝藻(含叶绿素和藻蓝素)可以光合作用。
17.核仁与rRNA的合成及核糖体的形成有关,代谢旺盛的细胞中,核孔数目多,核仁较大。

必修(一)分子与细胞第一章走近细胞第一节从生物圈到细胞一、相关概念细胞:是生物体结构和功能的基本单位。
除了病毒以外,所有生物都是由细胞构成的。
细胞是地球上最基本的生命系统。
生物圈是地球上最大的生命系统。
病毒无细胞结构,必需要依赖活细胞生存。
生命系统的结构层次:细胞→组织→器官→系统→个体→种群→群落→生态系统→生物圈病毒不属于生命系统;不是所有生物都具有这些结构层次,单细胞生物仅有一个细胞构成,植物没有系统层次二、病毒的相关知识:1、病毒是一类没有细胞结构的生物体。
主要特征:①仅具有一种类型的核酸,DNA或RNA,没有含两种核酸的病毒;②结构简单,一般由核酸(DNA或RNA)和蛋白质外壳所构成。
③专营细胞内寄生生活;2、根据病毒所含核酸种类的不同分为DNA病毒和RNA病毒(常见的RNA病毒有:SARS 病毒、(HIV)[引起艾滋病(AIDS)]、烟草花叶病毒等。
第二节细胞的多样性和统一性一、细胞种类:根据细胞内有无以核膜为界限的细胞核,把细胞分为原核细胞和真核细胞二、原核细胞和真核细胞的比较:1、原核细胞:细胞较小,没有成形的细胞核;遗传物质(一个环状DNA分子)集中的区域称为拟核;没有染色体(染色质),DNA不与蛋白质结合,;细胞器只有核糖体;有细胞壁(支原体除外),成分与真核细胞不同。
2、真核细胞:细胞较大,有核膜、有核仁、有成形的细胞核;有一定数目的染色体(DNA与蛋白质结合而成);一般有多种细胞器(如线粒体、叶绿体,内质网等)。
3、真核细胞与原核细胞的统一性:都有细胞质、细胞膜、核糖体、遗传物质都是DNA3、原核生物:由原核细胞构成的生物。
如:蓝藻(包括蓝球藻、颤藻和、念珠藻及发菜)、细菌(如硝化细菌、乳酸菌、大肠杆菌、肺炎双球菌)、放线菌、支原体等都属于原核生物。
4、真核生物:由真核细胞构成的生物。
如动物(草履虫、变形虫)、植物、真菌(酵母菌、霉菌、磨菇等)等。
蓝藻是细胞内含有藻蓝素和叶绿素,是能进行光合作用的自养生物。

腺相关病毒、腺病毒、慢病毒、逆转录病毒的区别优缺点及应用场景前言:病毒系统知识点get,四种主流基因转导病毒工具:腺相关病毒、腺病毒、慢病毒、逆转录病毒的区别优缺点及应用场景随着病毒生物学的发展,种类繁多的病毒载体已越来越成为向各种实验系统(如细胞系、原代细胞、组织脏器等)转运核酸的重要工具。
除了在实验室的组织培养和动物模型生产中发挥重要作用,它们还被用于治疗遗传性疾病的临床试验中。
目前主流的病毒载体系统主要包括慢病毒(LV)、(γ-)逆转录病毒(RV)、腺病毒(Ad)和腺相关病毒(AAV)。
我们分述之。
慢病毒和逆转录病毒均属于逆转录病毒科。
其典型特征为其RNA 基因组能逆转录为 cDNA 副本,cDNA 副本又能稳定整合至宿主细胞基因组中(这就是常说的稳转)。
逆转录病毒通常分两种:简单的(有时又称为致癌病毒或γ- 逆转录病毒,如鼠白血病病毒)和复杂的(如慢病毒)。
这些亚型间主要的差异在于复杂的逆转录病毒存在一些附属基因和调控基因,而简单的则没有(下文有进一步的讨论)。
这两类病毒颗粒都含有两份正链RNA,RNA 上附有病毒反转录酶(RT),它们位于病毒的内核(图1)。
位于内核的还有结构蛋白和酶,包括核壳(NC)、衣壳(CA)、整合酶(IN)和蛋白酶(PR)。
内核由一圈外周蛋白层包围,外周蛋白包括基质蛋白(MA),基质蛋白又被来源于宿主细胞细胞膜插有包膜糖蛋白的腹膜所包围。
图1. 简单和复杂逆转录病毒粒子结构。
病毒颗粒含有两份正链RNA,RNA 上附有反转录酶(RT),它们位于病毒的内核。
除此之外,内核还包含核壳(NC)、衣壳(CA)、整合酶(IN)和蛋白酶(PR)。
内核由一圈基质蛋白(MA)所包围,基质蛋白又被来源于宿主细胞细胞膜插有包膜糖蛋白(ENV)的腹膜所包围。
图片来源:汉恒生物实验室常用的慢病毒(Lentivirus)载体是以HIV-1(人类免疫缺陷1型病毒)为基础发展起来的基因治疗载体。
认识腺病毒作者:暂无来源:《家庭医学(上半月)》 2021年第2期这几年冬春季节,人们时常可以听到腺病毒这个词。
这是一种什么疾病,会引起哪些症状,对人体健康的危害大吗?腺病毒最容易在幼儿园、小学引起小范围的暴发流行,需要家长和老师们予以重视。
限于篇幅,有关腺病毒的西医治疗将发于本刊3月上期“家庭医生”栏目,敬请注意察看。
策划/执行家庭医学刊社主任医师施弦(湖南省儿童医院湖南长沙 410007)1953年,Rowe医生在一次扁桃腺切除手术后,做常规组织培养时偶然发现了一种微生物;后来,其他医生从新兵急性呼吸道感染者扁桃腺中也发现分离出同种病原体。
因其可以导致原代培养细胞发生自发性退化,曾被称为腺退行性病原体;1956年,国际病毒命名委员会(ICTV)正式将其命名为腺病毒。
腺病毒科分哺乳动物腺病毒属、禽类腺病毒属、腺胸腺病毒属和唾液酸酶腺病毒属4个属。
哺乳动物腺病毒可引起人畜共患急性传染病;腺病毒病人、隐性感染者是腺病毒的主要传染源,病毒繁殖和复制的主要场所是肠上皮细胞,故主要传播方式是粪-口传播。
初期感染也可能通过呼吸道传播且发病初期传染性最强(在此再次强调一下戴口罩的重要性),传染期1周左右。
隐性感染者虽无临床症状,但因腺病毒可以长期存在于扁桃体、消化道等,且数年后仍可少量排毒,加之活动不受限制,故亦有传染性。
除人类之间传播外,个别情况下,猪、鸡和野生水鸟亦可能成为传染源,在一定条件下可引起动物与人之间的传播。
全球腺病毒流行呈模式多样化、流行地区广泛、人群普遍易感的特点。
腺病毒流行一年四季均可发生, 但以夏季及冬春季多见。
1950~1960年亚洲和欧洲有大规模暴发, 发病率和病死率均高。
1958年冬季和1963年冬季, 我国华北、东北、西北都发生了较大规模的腺病毒肺炎流行, 疫情严重。
暴发流行常发生于环境密闭、空气流动性较差、环境较差的健康儿童(尤其是5岁以下儿童),成人在特殊环境中(比如部队训练营)亦有传播。
高中生物知识点归纳汇编(全)第一章、生命的物质基础第一节、组成生物体的化学元素名词:1、微量元素:生物体必需的,含量很少的元素。
如:Fe(铁、Mn(门、B(碰、Zn(醒、Cu(铜、Mo(母,巧记:铁门碰醒铜母(驴。
2、大量元素:生物体必需的,含量占生物体总重量万分之一以上的元素。
如:C(探、0(洋、H (亲、N(丹、S(留、P(人people、Ca(盖、Mg(美K(家巧记:洋人探亲,丹留人盖美家。
3、统一性:组成细胞的化学元素在非生物界都可以找到,这说明了生物界与非生物界具有统一性。
4、差异性:组成生物体的化学元素在细胞内的含量与在非生物界中的含量明显不同,说明了生物界与非生物界存在着差异性。
语句:1、地球上的生物现在大约有200万种,组成生物体的化学元素有20多种。
2、生物体生命活动的物质基础是指组成生物体的各种元素和化合物。
3、组成生物体的化学元素的重要作用:①C、H、O、N、P、S 6种元素是组成原生质的主要元素,大约占原生质的97%。
②.有的参与生物体的组成。
③有的微量元素能影响生物体的生命活动(如:B能够促进花粉的萌发和花粉管的伸长。
当植物体内缺B时,花药和花丝萎缩,花粉发育不良,影响受精过程。
--------------------------------------------------------------------------------2:高中生物知识点归纳第二节、组成生物体的化合物名词:1、原生质:指细胞内有生命的物质,包括细胞质、细胞核和细胞膜三部分。
不包括细胞壁,其主要成分为核酸和蛋白质。
如:一个植物细胞就不是一团原生质。
2、结合水:与细胞内其它物质相结合,是细胞结构的组成成分。
7、自由水:可以自由流动,是细胞内的良好溶剂,参与生化反应,运送营养物质和新陈代谢的废物。
8、无机盐:多数以离子状态存在,细胞中某些复杂化合物的重要组成成分(如铁是血红蛋白的主要成分,维持生物体的生命活动(如动物缺钙会抽搐,维持酸碱平衡,调节渗透压。
腺相关病毒操作手册一、腺相关病毒(Adeno-Associated Viral Vector,AAV)简介腺相关病毒属微小病毒科(parvovirus),为无包膜的单链线状DNA病毒。
AA V的基因组约4700bp,包括上下游两个开放读码框架(ORF),位于分别由145个核苷酸组成的2个反向末端重复序列(ITR)之间。
基因组中有3个启动子(P5、P19和P40)和2个开放阅读读框(ORF),rep和cap,如图1所示。
rep编码4个重叠的多功能蛋白,即Rep78、Rep68、Rep52和Rep40,其中Rep78与Rep68参与AA V的复制与整合,Rep52和Rep40具有解螺旋酶和ATP酶活性,与Rep78、Rep68共同参与单链基因组的复制;cap编码的VP1、VP2、VP3是装配成完整病毒所需要的衣壳蛋白,它们在AA V病毒整合、复制和装配中其重要作用。
图1.AA V基因结构二、腺相关病毒的优点1.安全性高:迄今从未发现野生型AA V对人体致病,重组AA V基因组序列上去除了大部分的野生型AA V基因组元件,进一步保证了安全性;2.免疫原性低:AA V2的基因组仅4681个核苷酸,便于用常规的重组DNA技术进行操作,而且进行动物实验时造成的免疫反应小;3.宿主范围广:能感染分裂细胞和非分裂细胞;4.表达稳定:能介导基因的长期稳定表达;5.物理性质稳定:在60℃不能被灭活,能抗氯仿;(汉恒--AAV包装)三、重组腺相关病毒载体系统简介AAV是一种复制缺陷型微小病毒,其增殖复制需要腺病毒或疱疹病毒的辅助。
AAV无辅助病毒系统(AAV Helper-Free System)可以在无辅助病毒的条件下生产出重组腺相关病毒。
在AAV Helper-Free System中,生产具有感染性的AAV病毒颗粒所需的腺病毒基因产物(如:E2A,E4等基因)大部分由pHelper质粒提供,其余的腺病毒基因产物由稳定表达腺病毒E1基因的AAV-293宿主细胞提供。
腺相关病毒的特性及应用进展石亮【摘要】腺相关病毒(AAV)是一种重要的载体.它与反转录病毒、慢病毒、腺病毒相比有不同的特点,如长期表达,安全性高,免疫原性低,高滴度生产能力,对某些组织感染效率高等.经过十多年的研究,它的生物特性已经被深入了解.在医学上,广泛应用于疾病的基因治疗,如糖尿病、乙型肝炎、癌症及一些免疫性疾病.生物学上,应用于基因功能的研究,作为转基因的载体,动物模型的建立及癌症的研究等.该文就AAV的基本生物学特性和应用进展进行综述.【期刊名称】《医学综述》【年(卷),期】2016(022)011【总页数】5页(P2088-2092)【关键词】腺相关病毒;载体;血清型【作者】石亮【作者单位】中国医学科学院医学实验动物研究所,北京100021【正文语种】中文【中图分类】Q782腺相关病毒(adeno-associated virus,AAV)作为一种理想的基因载体,利用病毒对细胞的感染能力,将携带的外源基因转移到细胞中,效率大大高于非病毒载体系统。
因此,人们利用AAV进行基因治疗,基因过表达,基因沉默等。
其最大的特点是不但能感染分裂期的细胞而且还能感染非分裂期的细胞,在体内长期表达存在,没有致病性。
现就AAV的基本生物学特性及其应用进展进行概述。
AAV属于细小病毒科,无包膜,正二十面体,直径20~26 nm,基因组大小为4.7 kb的单链DNA[1]。
AAV本身为复制缺陷病毒,只有在辅助病毒(腺病毒、疱疹病毒等)作用下才能复制增殖[2]。
它有两个开放的阅读框,末端重复序列位于开放阅读框的两侧。
左侧rep基因编码四个非结构性蛋白,对病毒的包装和复制起重要作用。
右侧的cap基因编码三个结构蛋白,负责病毒颗粒的整合、复制和装配。
AAV不但能感染分裂细胞也能感染非分裂细胞,在体内具有广泛的组织转导性包括心脏、肝脏[3]、肺[4]、骨骼肌、大脑、眼睛等[5-6]。
其具有长期高效的表达,无致病性,低免疫性等优良特征。
第一节AAV病毒的生活周期腺病毒伴随病毒(adeno-associated virus, AAV)是微小病毒科(Parvoviridae)家族的成员之一。
这一家族成员是一类微小、无被膜及具有二十面体结构的病毒。
病毒颗粒的直径在20~26nm之间,含有大小在4. 7~6kb之间的线状单链DNA基因组。
从昆虫到人类都已分离到微小病毒。
AAV病毒属于依赖性病毒类(De pendovirus),最初是在纯化的腺病毒液中发现的一种污染成分(Atchinson et al. 1965), 顾而得名。
从鸟类到许多哺乳动物包括人的体内都分离到各种血清型的AAV病毒。
大多数成年人都感染过AAV病毒,但尚未发现该病毒是任何疾病的致病因素。
在多数情况下,AAV在培养的正常细胞中不发生产毒性感染,只有在有辅助病毒包括腺病毒或疱疹病毒共同感染时才发生产毒性感染(Hoggan et al. 1966; Buller et al. 1981)。
因此,AAV病毒长期以来被认为是一种缺陷性病毒。
进一步的研究发现AAV病毒并非缺陷性病毒,而是在正常细胞中偏向于建立潜伏感染,仅在宿主细胞受到刺激时才被诱发进行感染性增殖。
AAV的生活周期有两种不同的胞内期。
在无辅助病毒存在时,AAV病毒颗粒进入细胞,脱衣壳后AAV的调节蛋白发生有限的表达,并抑制病毒基因的进一步表达和病毒DNA的复制。
这种负调节作用的结果是促进病毒基因组整合到宿主的基因组中建立潜伏感染。
AAV病毒偏向于整合到人基因组19号染色体q臂的特定位置(Kotin et al. 1990,1991,1992; Samulski et al. 1993)。
研究被AAV病毒潜伏感染的细胞发现,AAV对细胞表型往往有微弱影响,并影响细胞对刺激的反应能力(Yalkinoglu et al. 1988; Yakobson et al. 1987,1989; Bantel-Schaal et al. 1992, 1991)。
AAV生活周期的另一种胞内期是产毒性感染,往往在有辅助病毒(腺病毒或疱疹病毒)感染时发生。
辅助病毒的感染可以在AAV感染之前、同时或之后进行。
腺病毒(Ad)和单纯疱疹病毒(HSV)中与辅助功能有关的基因都已确定。
腺病毒中参与提供辅助功能的基因包括E1a, E1b, E2a, E4和VA RNA(Muzyczka 1 992),但不包括与腺病毒DNA复制直接相关的E2b基因产物(DNA聚合酶和末端酶)。
1型单纯疱疹病毒(HSV-1)提供辅助功能的情形则有些不同,不仅涉及与HSV病毒基因调节有关的基因包括ICP0和ICP4,还需要HSV病毒DNA复制的所必需的基因产物包括HSV螺旋酶-引物酶复合物(UL5,UL8,UL52)和主要DNA结合蛋白UL29的参与(Weindler F et al. 1991)。
这种在辅助功能方面的不同可能反映了腺病毒感染和疱疹病毒感染期间对宿主DNA合成机制的不同影响。
目前的关于AAV工作模式有以下要点:(1)AAV是一种生活周期以潜伏感染为主的病毒;(2)通过潜伏感染, 病毒基因组得以同细胞共存;(3)只要宿主细胞正常,AAV基因表达就处于抑制而维持潜伏状态;(4)如果细胞受到刺激,细胞内环境发生改变而表达应激基因;(5)导致细胞应激反应基因表达的调节状况也使得AAV基因表达,从而使AAV病毒复制;(6)产生子代病毒并释放,又感染新的正常宿主细胞,建立新的潜伏状态。
用某些可损伤基因的因素如紫外线照射、γ射线照射、各种化学致癌剂或某些代谢抑制剂处理某种类型的哺乳动物细胞,可使之在无辅助病毒存在的情况下能够发生AAV产毒性感染。
虽然这种情形比有辅助病毒存在时每个细胞产生AAV病毒的量要低几个数量级,但上述实验提示AAV病毒并不是一种缺陷性病毒,而是一种强烈偏向于潜伏的病毒。
返回标题第二节AAV病毒的基因组结构和功能人2型腺病毒伴随病毒(AAV-2)的基因组为4681 个核苷酸的单链DNA,全部序列已测定。
正链和负链DN A均可被包装到AAV病毒颗粒中。
AAV-2基因组的结构如图3-1所示。
基因组的两末端为145bp的倒转末端重复序列(inverted terminal repeat, ITR)。
ITR序列之间为AAV病毒的编码区,左侧的ORF编码4种Re p蛋白;右侧的ORF编码3种Cap蛋白。
一.ITR的结构和功能AAV 病毒ITR的序列和结构见图3-2。
每个ITR长145bp。
其中前125bp可形成一个T形的发夹结构,由两个小回纹结构(palindrome)(图2 B和C)组成,旁边为一个较大的回纹结构(A)。
ITR的序列可有两种构型:flip(B回纹接近3’端)或flop(C回纹接近3’端)。
一个特定分子AAV的两个末端形成flip或flop 的机率都相等。
单链形式的AAV ITR容易形成如图2所示的T型发夹结构,还可能形成另外的构型。
ITR序列是AAV病毒的复制、整合、拯救和包装所必需的顺式作用元件。
二.Rep基因的结构和功能Rep基因编码至少4种非结构蛋白:Rep78, Rep 68, Rep 52, Rep 40。
由p5启动子起始的2种mRNA翻译成Rep78和Rep68;p19启动子起始的2种mRNA翻译成Rep52和Rep40(图3.1)。
Rep52和Rep40蛋白C末端的氨基酸序列分别与Rep78和Rep68相同。
Rep 78和Rep68蛋白与AAV基因表达的正负调控有关。
这两种蛋白都可以和ITR中的特定序列( 5’-GCTCGCTCGCTC-3’)结合。
类似的序列在AAV的3个启动子上游均可找到。
当ITR自我折叠时可加强这种结合作用。
Rep78/Rep68具有ATP酶和螺旋酶的功能。
当其与ITR结合时便成为在124位点核苷酸处特异切割的缺口酶,识别的位点是5’-T/TGG。
这两种蛋白对于AA V生活周期的每一时期都是必需的。
在非允许条件下(没有辅助病毒和刺激因素),Rep蛋白只有微量表达,这种表达又能抑制其进一步的表达。
另外,在非允许条件下Rep的表达似乎可抑制AAV基因组以直接方式复制。
近来的研究证明Rep蛋白的表达对位点特异性整合是必需的。
相反,在允许条件下,Rep蛋白的表达对AAV病毒从整合状态的拯救、各种AAV基因的表达和AAV DNA的复制都是必需的。
Rep52和Rep40参与了病毒的装配,它们在病毒双链DNA合成中是不需要的,但在单链DNA和病毒颗粒的累积中则是必需的。
三.Cap基因的结构和功能Cap基因编码衣壳蛋白,其转录从p40启动子开始,形成约2.6kb和2.3kb mRNA,拼接后分别编码三个结构蛋白VP1、VP2和VP3(图2) ,分子量分别为87、73和61kDa,在成熟毒粒中的比例为1:1:10。
没有VP1时,VP2和VP3就可以包装子代单链DNA。
但这样的毒粒感染性较低,提示VP1在病毒颗粒的稳定性或感染性上是需要的。
VP2在病毒样空颗粒的装配中起重要作用。
VP3 似乎需要与其它两个中的一个一起,完成核定位任务。
返回标题第三节重组AAV的产生原理及常规制备、检测方法在基因治疗研究中,重组逆转录病毒和重组腺病毒在基因转移载体中一直占主导地位。
但近年来这一情况有所改变,用AAV作基因转移载体已成为基因治疗研究的热点。
这是由于AAV具有以下特点:(1)安全性好。
迄今从未发现野生型AAV对人体致病;重组AAV去除了野生型AAV基因组的96%,进一步保证了安全性;(2)宿主范围广。
不仅可转导分裂细胞,而且可转导静止期细胞;(3)野生型AAV可整合到人基因组19号染色体q臂的特定位置,利用这一特点,有可能研制出具有特异性整合功能的AAV载体;(4)AAV-2的基因组仅4681个核苷酸, 便于用常规的重组DNA技术进行操作;(5)物理性质稳定。
在60°C不能被灭活, 能抗氯仿;(6)重组AAV(rAAV)可长期稳定地表达外源基因。
一.重组AAV的产生原理目前在基因治疗研究中所用的AAV载体均由2型AAV改造而来。
AAV-2的ITR中包含了复制、包装、拯救及整合的信号,这使rAAV的产生成为可能:外源基因表达盒可完全取代AAV的编码序列,rep和cap基因产物则可由另一个质粒反式提供。
参照野生型AAV的生活周期,表达外源基因的rAAV的产生需要4种成分参与:1. 载体DNA。
载体DNA质粒包括了145bp的AAV ITR序列,外源基因的表达单位(通常由转录启动子、目的基因和多聚腺苷酸序列组成)位于两个ITR之间。
2. AAV的复制蛋白(Rep)和外壳蛋白(Cap)。
通常由携带它们相应基因的质粒(称为包装质粒)反式提供。
3. 宿主细胞。
多种细胞可用作产生重组AAV的宿主细胞,包括293细胞、HeLa细胞和KB细胞等,其中29 3细胞用磷酸钙转染的效率最高,因此最常用。
4. 辅助病毒。
常用的辅助病毒为腺病毒,疱疹病毒则较少用到。
腺病毒的辅助功能由E1a、E1b、E2a、E4 orf6和VA RNA基因提供;HSV-1的辅助功能由UL5、UL8、UL52和UL29等基因提供。
在辅助病毒存在的条件下,Rep蛋白可以将含有外源基因表达单位的rAAV基因组从载体质粒上拯救出来,并加以复制,得到大量复制形式的rAAV DNA。
随后,Cap蛋白可将单链的rAAV基因组包装成感染性的病毒颗粒。
二.重组AAV的常规制备及纯化方法rAAV载体的常规制备过程见图3.3。
通常采用磷酸钙共转染的方法,将载体质粒和包装质粒共转染经腺病毒感染的HeLa或293细胞(Hermonat and Muzyczka, 1984; Samulski et al., 1987,1989)。
当细胞在腺病毒感染下完全病变后(通常为感染后48-72 h),收集细胞,低速离心、去上清。
此时绝大部分rAAV存在于细胞内,去掉上清没有明显损失。
将细胞团块冻融3~4次,裂解液在55~60°C加热30~60min以灭活腺病毒。
低速离心去除细胞碎片后,将rAAV的粗提物用若干次氯化铯平衡梯度离心进行纯化。
如果rAAV基因组与野生型AAV基因组的大小一致,则密度为1.41 g/cm3,可以与密度为1.35 g/cm3的腺病毒明显分离。
对于转导培养的细胞,用rAAV的粗提物即可;但如果用于动物实验,则需进行若干次氯化铯平衡梯度离心进行纯化,因为即使污染了很少量的腺病毒(无论灭活与否)都能造成病毒接种部位的局部炎症反应。
在rAAV制备过程中,需注意以下问题:1.rAAV 的基因组长度(包括两个ITR在内)应接近野生型AAV基因组4681bp的长度,即外源基因表达盒(包括启动子、目的基因及多聚腺苷酸序列)不超过4.3 kb。