糖的生物合成
- 格式:doc
- 大小:423.50 KB
- 文档页数:25

糖的生物合成与代谢途径糖是生命中不可或缺的重要物质,它是生物体的主要能量来源之一,也是构成生物体的重要组成部分。
糖的生物合成与代谢是一系列复杂而精细的过程,它们通过一定的途径在细胞内进行。
在本文中,我们将探讨糖的生物合成与代谢的主要途径和相关机制。
第一节糖的生物合成糖的生物合成是细胞利用光能或化学能将无机物合成糖类化合物的过程。
主要的合成途径有光合作用和糖异生两种形式。
光合作用是指细胞通过叶绿体内的光化学反应,将二氧化碳和水转化为葡萄糖等有机物的过程。
在光照条件下,叶绿体中的叶绿素可以吸收太阳能,光合色素体可将太阳能转化为化学能,进而促使光合作用的进行。
光合作用分为光反应和暗反应两个阶段。
光反应发生在光合体系中,通过光合色素体捕捉光能,产生氧化还原电位,将光能转化为高能物质膜内的质子激励。
暗反应指的是光合作用中的还原和碳固定反应,主要在叶绿体基质内进行。
通过一系列酶的作用,将光反应所得的ATP和NADPH利用碳源还原为葡萄糖或其他有机物。
糖异生是指细胞在无光照条件下,通过有机物合成糖的过程。
糖异生主要发生在细胞质基质内,包括糖异生途径的两个重要过程:糖酵解和有机酸循环。
糖酵解是指将葡萄糖分解为丁醛酸,再将丁醛酸氧化为甲酸,最终合成糖的过程。
有机酸循环是指细胞质基质内的一系列反应,将葡萄糖分解为丙酮酸、柠檬酸等有机酸,最终通过一系列酶的作用合成糖。
第二节糖的代谢途径糖的代谢指的是细胞对糖化合物进行分解和利用的过程。
糖的代谢途径包括糖酵解、糖异生和糖氧化三个主要途径。
糖酵解是指细胞内部一系列酶的作用,将葡萄糖分解为丙酮酸或乙酸,产生ATP和还原能力分子NADH的过程。
糖酵解包括糖原糖酵解和异物糖酵解两种形式。
糖原糖酵解是指细胞内糖原被酵解,通过一系列的反应将糖原分解为葡萄糖,再进一步分解为丙酮酸,转化为乙酸最终释放能量。
异物糖酵解是指细胞利用外源性的碳水化合物,如蔗糖、木糖等进行糖酵解的过程。
糖异生是指细胞利用非糖类有机物合成糖的过程。


第七章糖的生物合成7.1 光合作用7.1.1 光合作用概述7.1.2 光能的吸收、转变和同化力产生7.1.3 光合的碳素途径(卡尔文循环)7.1.4 C4途径7.2 糖异生作用7.2.1 糖异生途径7.2.2 糖酵解和糖异生的互补调节7.3 蔗糖和多糖的生物合成7.3.1 糖核苷酸的作用7.3.2 蔗糖的生物合成7.3.3 淀粉(糖原)的合成7.3.4 纤维素的生物合成7.3.5 半纤维素的生物合成7.3.6 果胶的生物合成7.4 植物糖代谢的调节7.4.1 植物光合细胞丙糖、蔗糖、淀粉的相互转化7.4.2 果糖-2 , 6-二磷酸(F - 2 ,6 - BP)对糖酵解的调节7.4.3 光合作用形成的能量和还原力的外运7.4.4 植物光合细胞中糖酵解及蔗糖和淀粉合成的调节7.1 光合作用7.1光合作用光合作用(photosynthensis)是生物界中规模最大的有机合成过程,通过光合作用使太阳能转变为化学能贮存于碳水化合物中,每年约为8.36×1018 kJ。
放出的氧气约5 35×1011 t,同化的碳素约2×1011 t。
7.1.1光合作用概述光合作用的基本过程可用下式表示。
式中CO2是碳的氧化态,而生成物碳水化合物(CH2O)中的碳是相对还原态,因此,这是一个氧化还原反应。
CO2为氧化剂,在反应中被还原,H2O为还原剂,本身被氧化而提供CO2还原所需的电子。
CO2/(CH2O)系统的E′为-0.4 V,而O2/H2O的E′是+0.82 V,显然,在电子从水转移至CO2分子时是逆电势梯度(+1.22 V),因此,不能自发进行。
要使这一过程进行,必须供给能量。
在光合作用中,这些能量是由叶绿素吸收的光能提供的。
7.1.2光能的吸收、转变和同化力产生7.1.2.1光合色素和光化学反应1光合色素高等植物叶绿体中含有两类色素分子:叶绿素和类胡萝卜素。
叶绿素包括叶绿素a和b;类胡萝卜素包括胡萝卜素和叶黄素。




糖类生物合成途径及其应用研究糖类是人类和其他生物体内不可或缺的重要营养物质,也是许多药物的基础。
糖类的合成和利用涉及多种生物化学反应,其中最重要的是糖类的生物合成途径。
本文将介绍糖类的生物合成途径及其应用研究。
一、糖类生物合成途径1. 糖原生物合成途径糖原是一种储存多余能量的多糖,也是人体内最重要的能量储备物质。
糖原的生物合成途径包括两种途径:糖原合成途径和糖原分解途径。
糖原合成途径主要涉及到葡萄糖,通过多个酶催化反应将葡萄糖转化为α-1,4- -D-葡萄糖苷键之间的分枝多糖分子,最终形成糖原。
糖原分解途径,则是糖原的分解过程,将其转化为葡萄糖分子释放能量。
2. 葡萄糖合成途径葡萄糖是生命活动所必需的主要能量源,其生物合成途径也是多种反应的复杂组合。
葡萄糖的生物合成途径同样需要多种酶的参与,在体内主要通过六碳糖的环化来合成葡萄糖分子。
此外,生命体需要维持体内葡萄糖水平的稳定,因此在葡萄糖的生物合成途径中,还需要进行调节糖联的产生和分解等。
3. 糖类的修饰途径糖类的修饰起到了重要的作用,可以改变糖类的结构、功能、稳定性、相互作用等等。
常见的糖类修饰途径包括糖基化、乙酰化、硫化、酯化等。
其中,糖基化是最为常见和复杂的一种修饰方式,通过酶的催化反应将糖分子与蛋白质、核酸等生物大分子连接,形成糖蛋白、糖核酸等新的复合生物大分子,所修饰的糖类不仅可做生物活性调节剂,同时也被广泛应用于医药、农业等领域。
二、糖类合成途径在医药、化妆品等领域的应用研究1. 新型药物开发糖类合成途径在新型药物开发领域有着广泛的应用。
糖蛋白、糖核酸等复合生物大分子是人体内最基本的分子之一,其糖基化修饰的差异常常会影响到人体生理状况。
因此,针对人体糖基化修饰失调的疾病,如糖尿病、肿瘤等,研究人员可以开发新型药物,调节糖基化修饰的平衡,减轻疾病症状。
2. 化妆品制造糖类作为功能性成分,除了在医药领域广泛应用外,在化妆品领域也有着广泛的应用。


7糖的生物合成一、名词解释1、光合作用:含光合色素主要是叶绿素的植物和细菌,在日光下利用无机物质(CO2、H2O、H2S)合成有机物质,并释放氧气或其他物质的过程。
2、天线色素:全部叶绿素b、类胡萝卜素和大部分叶绿素,吸收光能并传递到作用中心色素分子。
3、作用中心色素:位于内囊体膜上具有特殊状态和光化学活性的少数叶绿素a分子,利用光能产生光化学反应,将光能转变成电能。
4、光合色素:5、光合磷酸化:在叶绿体ATP合成酶催化下依赖于光的由ADP和Pi合成ATP的过程。
6、糖异生:由简单的非糖前体转变为糖的过程。
糖异生不是糖酵解的简单逆转。
虽然由丙酮酸开始的糖异生利用了糖酵解中的7步近似平衡反应的逆反应,但还必须利用另外4步糖酵解中不曾出现的酶促反应绕过糖酵解中的三个不可逆反应。
二、填空1、光合作用分为光反应和暗反应两个阶段。
第一阶段主要在叶绿体的类囊体膜部位进行,第二阶段主要在叶绿体的基质部位进行。
2、高等植物光反应的最终电子供体是H2O,最终电子受体是NADP。
3、光合电子传递链位于叶绿体类囊体膜上,呼吸电子传递链位于线粒体内膜上。
4、光合磷酸化有环式和非环式两种类型。
5、在光合碳循环中,每固定6CO2形成葡萄糖,需消耗12NADPH+H+和18ATP。
6、C4植物的Calvin循环在维管束鞘细胞中进行,而由PEP固定CO2形成草酰乙酸是在叶肉细胞中进行。
7、糖异生主要在肝脏(细胞溶胶)中进行;糖异生受Pi、AMP、ADP抑制,被高水平ATP、NADH激活。
8、在糖异生作用中由丙酮酸生成PEP,在线粒体内丙酮酸生成草酰乙酸是丙酮酸羧化酶催化的,同时要消耗ATP;然后在细胞质内经PEP羧激酶催化,生成磷酸烯醇丙酮酸,同时消耗GTP。
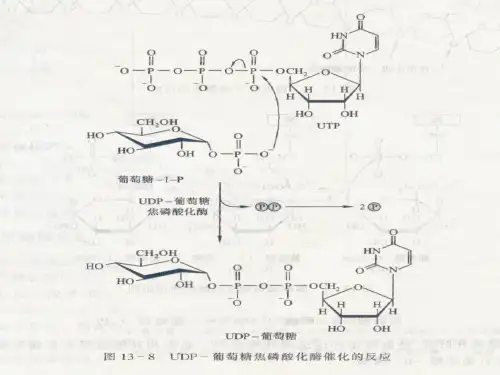
9、植物体内蔗糖合成酶催化的蔗糖生物合成中葡萄糖的供体是UDPG,葡萄糖基的受体是果糖。
10、合成糖原的前体分子是UDPG,糖原分解的产物是G-1-P。
三、单项选择题1、用于糖原合成的葡萄糖-1-磷酸首先要经什么化合物的活化?A、ATPB、CTPC、GTPD、UTPE、TTP2、RuBisCO催化RuBP羧化反应的产物是(RuBisCO-核酮糖-1,5-二磷酸羧化酶;RuBP—核酮糖-1,5-二磷酸;PGA-3-磷酸甘油酸)A、PGAB、PEPC、OAAD、IAA3、不能经糖异生合成葡萄糖的物质是:(乙酰CoA只能进入TCA分解,不能经糖异生合成葡萄糖)A、α-磷酸甘油B、丙酮酸C、乳酸D、乙酰CoAE、生糖氨基酸4、丙酮酸羧化酶是那一个途径的关键酶:A、糖异生B、磷酸戊糖途径C、胆固醇合成D、血红素合成E、脂肪酸合成5、动物饥饿后摄食,其肝细胞主要糖代谢途径:A、糖异生B、糖有氧氧化C、糖酵解D、糖原分解E、磷酸戊糖途径6、下面哪种酶在糖酵解和糖异生中都起作用:A、丙酮酸激酶B、丙酮酸羧化酶C、3-磷酸甘油醛脱氢酶D、己糖激酶E、果糖1,6-二磷酸酯酶7、糖异生途径中哪一种酶代替糖酵解的己糖激酶?A、丙酮酸羧化酶B、磷酸烯醇式丙酮酸羧激酶C、葡萄糖-6-磷酸酶D、磷酸化酶8、光合作用中Calvin循环是在叶绿体的:A、外膜上进行B、基粒上进行C、基质中进行D、类囊体腔内进行9、电子在环式光合电子传递链中传递时可产生:A、NADPHB、O2C、ATPD、NADH10、非环式光合电子传递链中,最终的电子受体是:A、H2OB、NADC、NADPD、ADP11、光合作用中,将CO2还原为糖类的“同化力”来源于:A、光反应B、暗反应C、光呼吸D、暗呼吸12、在光合作用的光反应中,作用中心分子的作用是将:A、电能转变为化学能B、光能转变为电能C、光能转变为化学能D、化学能转变为电能13、光合作用释放的O2来源于:A、H2OB、CO2C、RuBPD、PEP14、下列那个是各糖代谢途径的共同中间产物:A、6-磷酸葡萄糖B、6-磷酸果糖C、1,6-二磷酸果糖D、3-磷酸甘油醛E、2,6-二磷酸果糖(葡萄糖经过激酶的催化转变成葡萄糖-6-磷酸,可进入糖酵解途径氧化,也可进入磷酸戊糖途径代谢,产生核糖-5-磷酸、赤鲜糖-4-磷酸等重要中间体和生物合成所需的还原性辅酶Ⅱ;在糖的合成方面,非糖物质经过一系列的转变生成葡萄糖-6-磷酸,葡萄糖-6-磷酸在葡萄糖-6-磷酸酶作用下可生成葡萄糖,葡萄糖-6-磷还可在磷酸葡萄糖变位酶作用下生成葡萄糖-1-磷酸,进而生成糖原。

糖原合成途径糖原合成途径是指生物合成糖原的过程,它是维持机体能量平衡的重要途径之一。
糖原合成途径发生在细胞内,主要是在肝细胞和肌肉细胞中进行。
糖原是由多个葡萄糖分子连接而成的聚糖,保存在肝脏和肌肉中,用于身体需要增加能量供应的时候释放出来。
糖原合成途径主要基于胰岛素的调节。
当血糖水平升高时,胰岛素被释放出来,刺激肝脏和肌肉细胞开始合成糖原,将多余的葡萄糖转化为储存形式。
糖原合成途径主要分为以下几个步骤:1. 葡萄糖转化为葡萄糖-6-磷酸葡萄糖经过磷酸化反应变成葡萄糖-6-磷酸,这个反应需要消耗ATP,同时需要一种叫做磷酸果糖激酶的酶来催化反应。
2. 葡萄糖-6-磷酸转化为葡萄糖-1-磷酸葡萄糖-6-磷酸再经过一次磷酸化反应变成葡萄糖-1-磷酸,这个反应的催化剂是磷酸葡萄糖异构酶。
这个过程是可逆的,也就是说,葡萄糖-1-磷酸也可以通过磷酸果糖激酶反应变成葡萄糖-6-磷酸。
3. 葡萄糖-1-磷酸转化为UDP-葡萄糖葡萄糖-1-磷酸经过羧化反应变成UDPG。
这个反应由一个酶叫做UDP-葡萄糖合成酶催化。
4. UDP-葡萄糖与糖原核心部分相连核心糖原分子由苏打岩附着蛋白组成,UDP-葡萄糖与糖原核心部分相连后,就形成一个长链的糖原分子。
这个过程由一种酶叫做糖原合成酶催化。
5. 分支酶调节糖原合成的分支情况糖原分子的自我调节可以通过分支酶进行。
分支酶可以在糖原的表面上挂上分支,增加糖原的表面积,更方便地储存更多的葡萄糖。
这个过程由一种酶叫做糖原分支酶催化。
总的来说,糖原合成途径是一个复杂的过程,需要多种酶的参与,同时也需要外部的调节因素控制。
当人体需要能量时,这些糖原分子就会被释放出来,分解成单糖分子,供给身体需要能量的器官。
而当血糖水平升高时,胰岛素就会调节体内葡萄糖的合成和储存,维护人体的稳态。
合成糖概念
一、糖的合成
糖的合成是指通过化学或生物合成的方法,将不同的碳源、氢源和氮源等基本原料转化为糖的过程。
根据所需合成的糖的类型不同,可以采用不同的合成方法。
常见的糖类包括单糖、双糖和多糖,其中单糖是最简单的糖类,也是其他糖类的基本单元。
二、合成糖的方法
1.化学合成法:化学合成法是利用不同的化学原料,通过一系列的化学反应,
最终得到目标糖的过程。
该方法具有反应速度快、产率高等优点,但同时也存在反应条件苛刻、需要使用大量有机溶剂等缺点。
2.生物合成法:生物合成法是利用微生物或酶催化剂,将不同的碳源、氢源
和氮源等基本原料转化为糖的过程。
该方法具有反应条件温和、对环境友好等优点,但同时也存在反应速度慢、产率低等缺点。
三、合成糖的应用
1.食品工业:合成糖在食品工业中应用广泛,如糖果、饮料、甜点等食品的
生产。
通过使用合成糖,可以控制食品的甜度、口感和质地等方面,提高食品的品质和口感。
2.制药工业:合成糖在制药工业中也有着广泛的应用,如抗生素、抗病毒药
物和抗肿瘤药物等的生产。
通过使用合成糖,可以控制药物的化学结构和药理活性,提高药物的疗效和安全性。
3.生物技术领域:合成糖在生物技术领域中也有着重要的应用,如糖蛋白、
糖脂和多糖等生物分子的合成。
通过使用合成糖,可以研究这些生物分子的结构和功能,为疾病诊断和治疗提供新的思路和方法。
4.材料科学领域:合成糖在材料科学领域中也有着一定的应用,如生物降解
塑料、生物医用材料和生物粘合剂等的制备。
通过使用合成糖,可以改善这些材料的生物相容性和降解性能,提高其安全性和实用性。
糖的合成与降解途径糖是一种重要的碳水化合物,广泛存在于自然界中。
它不仅是人体能量的主要来源,还在许多生物过程中起着关键的作用。
本文将探讨糖的合成和降解途径,并介绍其在生物体内的重要性。
一、糖的合成途径1. 光合作用:光合作用是植物通过光能转化为化学能的重要过程。
在光合作用过程中,植物通过光能和二氧化碳的参与,合成葡萄糖等糖类物质。
光合作用包括光能捕捉、光反应和暗反应三个阶段。
其中,暗反应是主要的糖合成过程,通过酶的催化,将二氧化碳和水转化为葡萄糖,并释放出氧气。
2. 糖异生:糖异生是指利用非糖底物合成糖的过程。
主要发生在肝脏、肾脏和肠道等器官。
在糖异生过程中,葡萄糖的前体物质如乳酸、甘油和氨基酸等被逐步转化为葡萄糖,以满足机体对能量的需求。
糖异生在长时间禁食或低血糖状态下起着重要作用,维持血糖水平的稳定。
3. 糖核酸代谢:在糖核酸代谢过程中,糖原、DNAs、RNAs等被分解为核苷酸,然后通过一系列酶的催化作用,再合成成糖核酸。
糖核酸代谢不仅提供了细胞所需的能量,还参与了遗传物质的传递和遗传信息的编码。
二、糖的降解途径1. 糖酵解:糖酵解是指在没有氧气的条件下,将葡萄糖分解为乳酸或乙醇和二氧化碳。
糖酵解途径主要发生在肌肉和红血球等缺氧环境下的细胞中,产生少量ATP的同时,通过再氧化生成的NAD+补充酵母菌在氧气缺乏时的能量需求。
2. 无氧糖解:无氧糖解是指在缺氧环境下,将葡萄糖转化为乳酸。
这种过程比糖酵解产生的ATP更少,但可以在氧气供应不足时提供急需的能量。
3. 糖酸循环:糖酸循环,又称为Krebs循环或三羧酸循环,是有氧呼吸的关键步骤。
它将葡萄糖酸分解为二氧化碳和能量丰富的电子转移物质NADH和FADH2,并产生一定量的ATP。
4. 脂肪酸合成:在饥饿或低血糖状态下,糖原储备不足,机体会将葡萄糖转化为脂肪酸以供应能量需求。
脂肪酸合成主要发生在肝脏和脂肪细胞中,通过乙酰辅酶A催化,将葡萄糖转化为三酰甘油。
基础生物化学Basic Biochemistry
9 糖的生物合成
9.1 光合作用
9.2 糖异生作用
9.3 蔗糖的生物合成
9.4 淀粉和糖原的生物合成
蔗糖的结构
α-D-吡喃葡糖基-(1→2)-β-D-呋喃果糖苷
9.3.1 糖核苷酸的作用
葡萄糖和果糖不能直接合成寡糖和多糖,必需经活化变成活化葡萄糖( UDPG、ADPG 、GDPG )才能参与反应。
9.3.2 蔗糖的生物合成
高等植物合成蔗糖的途径有两条:
①蔗糖合酶催化的合成途径
UDPG + 果糖→ 蔗糖+ UDP
该途径主要是起蔗糖分解的作用,在贮藏器官中主要起分解蔗糖产生UDPG,用于淀粉的合成。
蔗糖+ UDPG →淀粉
②磷酸蔗糖合酶催化的合成途径
UDPG + F-6-P → 磷酸蔗糖+ UDP
光合组织中磷酸蔗糖合酶活性高,在磷酸蔗糖磷酸酶的催化下,磷酸蔗糖水解生成蔗糖。
磷酸蔗糖+H
O→蔗糖+Pi
2
蔗糖合成的两条途径:
②
①
③蔗糖磷酸化酶催化的途径(微生物中)
G-1-P + 果糖 蔗糖+ Pi。
7 糖的生物合成第七章糖的生物合成7.1 光合作用7.1.1 光合作用概述7.1.2 光能的吸收、转变和同化力产生7.1.3 光合的碳素途径(卡尔文循环)途径7.1.4 C47.2 糖异生作用7.2.1 糖异生途径7.2.2 糖酵解和糖异生的互补调节7.3 蔗糖和多糖的生物合成7.3.1 糖核苷酸的作用7.3.2 蔗糖的生物合成7.3.3 淀粉(糖原)的合成7.3.4 纤维素的生物合成7.3.5 半纤维素的生物合成7.3.6 果胶的生物合成7.4 植物糖代谢的调节7.4.1 植物光合细胞丙糖、蔗糖、淀粉的相互转化7.4.2 果糖-2 , 6-二磷酸(F - 2 ,6 - BP)对糖酵解的调节7.4.3 光合作用形成的能量和还原力的外运7.4.4 植物光合细胞中糖酵解及蔗糖和淀粉合成的调节7.1 光合作用7.1光合作用光合作用(photosynthensis)是生物界中规模最大的有机合成过程,通过光合作用使太阳能转变为化学能贮存于碳水化合物中,每年约为8.36×1018 kJ。
放出的氧气约535×1011 t,同化的碳素约2×1011 t。
7.1.1光合作用概述光合作用的基本过程可用下式表示。
式中CO2是碳的氧化态,而生成物碳水化合物(CH2O)中的碳是相对还原态,因此,这是一个氧化还原反应。
CO2为氧化剂,在反应中被还原,H2O为还原剂,本身被氧化而提供CO2还原所需的电子。
CO2/(CH2O)系统的E′为-0.4 V,而O2/H2O的E′是+0.82 V,显然,在电子从水转移至CO2分子时是逆电势梯度(+1.22 V),因此,不能自发进行。
要使这一过程进行,必须供给能量。
在光合作用中,这些能量是由叶绿素吸收的光能提供的。
7.1.2光能的吸收、转变和同化力产生7.1.2.1光合色素和光化学反应1光合色素高等植物叶绿体中含有两类色素分子:叶绿素和类胡萝卜素。
叶绿素包括叶绿素a和b;类胡萝卜素包括胡萝卜素和叶黄素。
这些色素分子与叶绿体类囊体膜上的蛋白质形成色素蛋白复合物,完成对光能的吸收、传递和光化学反应。
根据色素的作用可将其分为天线色素(辅助色素)和作用中心色素。
天线色素(antenna pigment)包括全部叶绿素b、类胡萝卜素和大部分叶绿素a,它们的功能是吸收光能并传递到作用中心色素分子。
作用中心色素(reaction centre pigment)是位于类囊体膜上具有特殊状态和光化学活性的少数叶绿素a分子,其作用是利用光能产生光化学反应,将光能转变成电能。
2 光化学反应根据吸收光波长的不同,把作用中心色素分为两类:P700(700 nm)和P680(680 nm),它们分别是色素蛋白复合物光系统Ⅰ(photosystem Ⅰ,PSⅠ) 和光系统Ⅱ(photosystemⅡ,PSⅡ)的光合作用中心色素。
在高等植物中光合作用中心是指叶绿体中进行光合作用原初反应的最基本的色素蛋白结构,至少包括一个作用中心色素分子P(代表P680或P700)、一个原初电子受体(A)和一个原初电子供体(D)。
A和D分别是直接接受或供给作用中心色素电子的物质。
光化学反应发生时,作用中心色素P接受光能被激发成激发态P*,此时P*的一个电子被激发处于高能轨道,极易失去。
P*把1个电子传给原初电子受体A,使A变成A-,P*失去电子后回到基态变成P+,P+对电子有极大的吸引力,再从原初电子供体D得到一个电子,本身恢复成P而D变成D+,实现了电荷的分离。
7.1.2.2光合电子传递链(photosynthetic chain)如上述,在光合作用中水中的电子经过一系列的电子递体的传递,最后到达NADP+。
这些递体在类囊体膜上是有序的排列,互相衔接着,被称为电子传递链。
如果把这些物质按其氧化还原电位(E′)排列起来,其形状像英文中Z,所以又称为Z链,如图7-1所示。
从图上可以看出:图7-1高等植物光合作用电子传递链1 通过光对两个作用中心色素分子P680和P700的激发,提高了P*680和P*700的氧化还原电势,H2O中的电子逆电势传递到NADP+。
但在P*680→P700和P*700→NADP+之间是顺电势梯度的自发过程。
2 电子传递过程是电子递体之间的一系列氧化还原反应。
3 电子传递的结果是把光能变成电能,又变成了NADPH+H+中的活跃的化学能。
同时在电子传递过程中还偶联ATP的产生,这也是一个把光能转变成活跃化学能的过程。
光合作用中通过电子传递形成NADPH+H+和ATP,合称之为同化力,用于后文提到的卡尔文循环中CO2的固定和还原,从而形成有机化合物糖。
4 光合链电子的最终供体为H2O,这就导致水的光解,形成光合放氧。
Z链是按电子传递体的生物氧化还原电势排列的。
它并不反映这些物质在类囊体中的排列状况。
图7-2显示了电子传递体在类囊体中实际排列的电子传递链。
图7-2质子梯度的形成和ATP产生7.1.2.3光合磷酸化叶绿体利用光能使ADP+Pi生成ATP的反应,称之为光合磷酸化(photosynthetic phosphorylation)。
利用光能生成ATP的过程有两种。
一种是来自于水的电子经过PSⅡ,Cytb6/f和PSⅠ的传递到达NADP+,在传递过程中释放能量用于ADP磷酸化生成ATP,同时将NADP+还原成NADPH+H+。
此过程其电子传递是开放的,所以称之为非环式光合磷酸化。
另一种是PSⅠ的电子传给Fd后,再传给Cytb6/f,然后经PC又回到PSⅠ,形成一个环式电子流。
在电子流动过程中释放的能量使ADP+Pi形成ATP,因其电子传递路程是闭合的,所以称之为环式光合磷酸化。
形成ATP的机理可以用化学渗透学说来解释(参阅62)。
如在图7-3非环式电子传递过程中,当来自水的电子还原PQ成PQH2时,要从叶绿体基质中得到两个质子,而当PQH2将电子传给Cytb6/f时,要将两个质子释放到类囊体腔中,类囊体膜对质子是不可随便通过的。
因此使腔内的质子浓度大于基质的质子浓度。
即利用电子传递释放的能量建立了一个质子势。
当质子通过ATP合酶从腔中进入基质时,就利用这部分能量使ADP+Pi形成ATP。
这样,就很好地解释了非环式光合磷酸化中ATP的形成(图7-3)。
而对于环式光合磷酸化,因为电子传递过程没有完全搞清楚,因此没有一个满意的答案。
环式电子传递过程是PSⅠ→Fd→Cytb6/f→PC→PSⅠ时,就同样可以建立类囊体膜内外的质子势,用于ATP 的形成。
这一问题的解决还有待进一步研究。
7.1.3光合的碳素途径(卡尔文循环)基本的光合碳素途径——还原的戊糖途径是1946年M Calvin等科学家用单细胞绿藻作试验材料,应用14C示踪技术并结合纸上层析法,经过十年努力搞清楚的光合作用碳素同化途径。
因此也称作卡尔文循环(Calvin cycle)。
由于卡尔文在光合作用碳转化途径上作出了重大贡献,他于1961年获得诺贝尔奖。
卡尔文循环的最初产物为3-磷酸甘油酸,因此,此途径也称C3途径。
整个循环可分为三个阶段。
1 CO2的固定CO2与核酮糖-1,5-二磷酸(ribulose bisphosphate,RuBP)反应生成两分子三磷酸甘油酸(3-PGA)。
在此反应中,酶的作用使RuBP异构成烯醇式二磷酸核酮糖,后者羧化成中间产物2-羧基,3-酮基-1,5-二磷酸核糖,再加水分解生成2分子3-磷酸甘油酸(3-PGA)。
这一反应由核酮糖-1,5-二磷酸羧化酶(ribulose bisphosphate carboxylase oxygenase, Rubisco)催化,该酶位于叶绿体间质中,含量占叶片可溶性蛋白一半以上。
由八个大亚基和八个小亚基组成,大小亚基分别由叶绿体基因和核基因编码。
催化部位在大亚基上,而小亚基则具有调节作用。
该酶还具有加氧酶活性,加氧产物为3-PGA和磷酸乙醇酸,加氧和羧化作用发生在同一个活性中心,而且两种活性均可为CO2和Mg2+所活化。
2羧化产物的还原包括两步反应:3-PGA在激酶催化下磷酸化生成1,3-二磷酸甘油酸(1,3-diPGA);后者在脱氢酶催化下还原为3-磷酸甘油醛。
此反应所消耗的ATP和NADPH 称之为光合同化力,来自光合作用的光反应。
反应产物3-磷酸甘油醛是一个三碳糖。
3 RuBP的再生由一系列转酮酶、转醛酶和异构酶催化,经10步反应(表7-1,图7-4)使RuBP再生。
反应及酶类似于磷酸戊糖途径中分子重排阶段的逆过程。
表7-1光合作用碳还原循环的反应卡尔文循环的总反应式中的6-磷酸果糖可进一步转化为葡萄糖: 总反应式可写成:图7-4卡尔文循环(C3循环)表明每同化1分子CO2需3分子ATP和2分子NADPH。
7.1.4 C4途径M.D.Hatch和C。
R。
Slack发现,某些起源于热带的植物例如甘蔗、玉米等在光合作用中还存在一种辅助途径——C4途径。
它的作用是固定、转运和集中CO2到C3途径所在的维管束鞘细胞中,使其中CO2浓度升高,从而提高了光合速率。
C4途径开始于叶肉细胞中,在磷酸稀醇式丙酮酸羧化酶(PEP羧化酶)的作用下,CO2与磷酸烯醇式丙酮酸(PEP)缩合形成草酰乙酸。
后者是一种含四个碳原子的二羧酸,故将该反应途径叫C4途径。
在某些C4植物中草酰乙酸被转变成苹果酸,在另一些植物中它也可被转变成天冬氨酸,然后转入维管束鞘细胞中,经过脱羧作用分解成CO2和一个C3化合物(如PEP),C3化合物被转运回叶肉细胞中,进行下一次固定CO2的循环;CO2则进入卡尔文循环,形成糖。
简要过程见图7-5。
图7-5C4途径的简要过程在C4植物中,卡尔文循环只存在于维管束鞘细胞中,这些细胞中的O2浓度较低,而又由于C4途径的转运和集中,CO2浓度升高,因而提高了细胞中的CO2/O2之比,这有利于RuBisco的羧化作用而不利于其加氧作用,提高了光合作用的速率。
7.2糖异生作用糖异生(gluconeogenesis)作用是由非糖前体如丙酮酸、草酰乙酸等合成葡萄糖的过程。
可通过糖酵解的逆过程完成,但糖异生途径又非糖酵解的简单逆转。
在糖酵解中,由己糖激酶、磷酸果糖激酶和丙酮酸激酶催化的反应是不可逆的,若以另一些酶代替,这三步反应即可逆(图7-6)。
图7-6糖酸解和糖异生的比较7.2.1糖异生途径1.丙酮酸生成磷酸烯醇式丙酮酸通过两步反应: (1) 丙酮酸羧化酶催化丙酮酸羧化生成草酰乙酸:丙酮酸羧化酶是一个生物素蛋白,需乙酰CoA和Mg2+激活。
该酶定位于线粒体,丙酮酸需经运载系统进入线粒体后才能羧化成草酰乙酸,后者只有在转变为苹果酸后才能再进入细胞质。
苹果酸再经胞质中的苹果酸脱氢酶转变成草酰乙酸,才能进一步转变成PEP。
(2) 磷酸烯醇式丙酮酸羧激酶催化草酰乙酸形成PEP:PEP沿酵解途径逆向反应转变成1,6-二磷酸果糖。