多孔动物门主要特征
- 格式:doc
- 大小:73.50 KB
- 文档页数:6


动物学——多孔动物门(海绵动物门)
是最原始、最低等的多细胞动物;为多细胞动物进化中的一个侧支
一、主要特征
1.水中固着生活、体型多不规则
2.细胞水平的多细胞动物(有细胞分化,无组织分化)
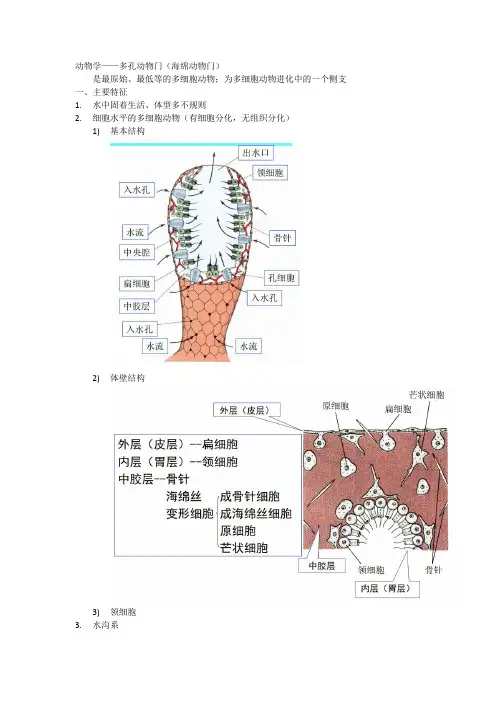
1)基本结构
2)体壁结构
3)领细胞
3.水沟系
水沟系是多空动物特有的结构,与其适应固着生活相关,多孔动物生物摄食、呼吸、排泄等生理活动都要借水的穿行来完成。
水沟系的类型:
4.生殖与胚胎发育
1)无性生殖:出芽生殖和形成芽球(芽球是多孔动物的繁殖方式,也是休眠体);
2)有性生殖:
I.雌雄同体或异体,异体受精;
II.卵和精子由原细胞发育来;
III.卵大,留在中胶层,同体的精子不能直接入卵,需随水流进入另一海绵体内,领细胞吞食精子后,鞭毛和领消失,成为变形虫状,将精子带入卵,与之受精。
3)胚层逆转
二、多孔动物的分类地位
1.最原始、最低等的多细胞动物
1)只有细胞分化、无组织分化;
2)无消化腔,只有细胞内消化(领细胞);
3)无神经系统;
4)细胞分化程度低,再生能力强。
机体所有细胞参与结构与机能的完全重新组织,形成一个新个体,称为体细胞胚胎发生。
2.侧生动物
胚胎发育中有胚层逆转现象,构造上有领细胞、水沟系、骨针等特殊结构——侧生动物,是很早由原始群体鞭毛虫发展来的一个侧支,不再演化为其他类群的多细胞动物。
但新的研究表明,多孔动物的滤食性摄食方式、原细胞的分化特征和细胞全能性、原始的神经细胞等,也说明其是处于原生动物和后生动物之间的中间类型。

多孔动物门知识点总结多孔动物门的形态特征多孔动物门的形态特征主要有以下几点:1. 体表无真皮、外胚层和中胚层2. 体内有细胞如白细胞活动,这些细胞代表了多孔动物门动物的基本形态,具有原始感觉细胞、运动细胞、营养细胞3. 水管系统:多孔动物门没有真正的组织器官,但它们有一套独特的水管系统,通过这个系统,多孔动物门可以在体内外换气和摄食。
水管系统由脂肪细胞、领细胞和囚细胞等细胞构成,这些细胞可以协同工作,完成海绵内部环境的维护。
4. 体内大多数细胞无特异性,极个别细胞可以分泌骨骼(trabecular)、颇似有生境,形成团体状多孔动物门的生活史多孔动物门的生活史主要有以下几个阶段:1. 孢子固着:多孔动物门的研究表明,多孔动物门有着比较特殊的生活史。
它们在幼年阶段的生活史大致可以分为孢子固着阶段和形态成熟阶段。
孢子固着是多孔动物门幼年期的第一阶段,它在该生命阶段时大多数动物表现为重要形态阶段。
2. 形态成熟:多孔动物门的幼年期生活史终结于孢囊被激活后,开始展现出成年状态。
这是多孔动物门生活史中的第二个阶段。
其主要特征是孩子和成人破裂,从而充分发挥自己的功能。
3. 个体生长:多孔动物门的个体生长没有固定的规律,海绵的生长速度极为缓慢。
有的海绵一生只长一公分,有的则能够长成一个有几米高的大型骷髅架。
它们的生长受到多种因素的影响,比如水温、光照、食物等。
多孔动物门的分类多孔动物门可以分为以下几个大类:1. 海绵状动物:海绵状动物类(Calcarea)是多孔动物门中最原始的类群,包括了大多数不定节数目的点钙棘、点钟海绵、玉通玛瑙、点钟海绵的钙棘等。
它们的身体呈不规则的形状,体表有许多细孔,这些细孔是水管系统的一部分。
2. 硅质海绵:硅质海绵类(Demospongiae)是多孔动物门中数量最多的类群,它们广泛分布在全球各大洋和海域。
硅质海绵类的特点是,它们的细胞由硅片构成,硅质海绵类的细胞结构较为松散,生长速度也较快。


第5章多孔动物门11生态学姓名:潘小微学号:11226010(一)名词解释1、芽球:芽球的形成是在中胶层中,由一些储存了丰富营养的原细胞聚集成堆,外包以几丁质膜和一层双盘头或短柱状的小骨针,形成球形芽球。
芽球可以在条件不适宜时生存下来,带条件适宜时才发育成新个体。
2、水沟系:水沟系是海绵动物所特有的结构,它对适应固着生活很有意义。
不同种的海绵其水沟系有很大差别,其基本类型包括单沟型、双沟型和复沟型3种。
水沟系的产生有利于海绵动物得到氧气和食物,同时不断排除废物,对海绵生命活动与适应环境都有利。
3、两囊蚴虫:就钙质海绵来说受精卵进行卵裂,形成囊胚,动物极的小细胞向囊胚腔内生出鞭毛,另一端的大细胞中间形成一个开口,后来囊胚的小细胞由开口倒翻出来,里面小细胞具鞭毛的一侧翻到囊胚表面。
这样,动物极的一端为具鞭毛的小细胞,植物极的一端为不具鞭毛的大细胞,此时称为两囊幼虫。
4、胚层逆转:幼虫从母体出水孔随水流逸出,然后具鞭毛的小细胞内陷,形成内层,而另一端大细胞留在外边形成外层细胞,着与其他多细胞动物原肠胚形成正相反,因此称为胚层逆转。
(二)判断与改错1、海绵动物主要生活于海水中,全部营漂游生活。
(F )(全部营固着生活)2、由于海绵动物体表有许多小孔,故又名多孔动物。
(T )3、海绵动物体表的一层细胞为领细胞,具保护作用。
(F )(消化食物)4、海绵动物体内为一层特有领细胞,能摄取食物进行细胞内消化。
(T )5、单沟型水沟系的水流途径是:进水小孔―――中央腔――――出水孔。
(T )6、复沟型水沟系的水流途径是:入水小孔―――流入管―――前幽门孔―――辐射管―――后幽门孔――――流出管―――中央腔―――出水孔。
(F )(辐射管改为鞭毛室)(三)填空题1、海绵动物体壁的结构包括扁细胞,中胶层,领细胞三层。
2、海绵动物中胶层由于含有骨针和海绵质纤维,因而能起到骨骼支持的作用。
3、海绵动物的三类水沟系反映了其进化过程,从单沟型到双沟型到复沟型,领细胞数目逐渐增多,使水流通过海绵体的速度和流量增加,从而使海绵动物得到更多的食物和氧气,有利其生命活动。


多孔动物门主要特征简介多孔动物门(Porifera)--海绵动物界的一门。
大约5,000种原始多细胞水生动物的统称。
除针海绵属(Spongilla)约20种为淡水产外,多孔动物门均分布在海洋的潮间带到8,500公尺(28,000余尺)深处,营固着生活。
由于海绵常呈分枝形,而且不会移动,从前被人们当作植物。
多孔动物为原始的多细胞动物,本动物门也称海绵动物门(Spongiatia),一般称之为海绵(Sponge)。
海绵是重要的造礁生物,少数属种也有一定的地层意义。
形态构造多孔动物的外形变化很大,除少数种类外,往往没有对称面,在许多方面与低等植物相似,常被描述为块状、垫状、球状、指状、树枝状、杯状或漏斗状等。
一般来说,深海种类的身体常趋于对称,具柄状体,固着在海底软泥上,由一个或成束的强大骨针形成柄或轴,下端深入泥中,上端将海绵体高高托起。
有的种类基部有须根状的骨针,将海绵固着在海底上。
多孔动物的基本构造,可用单沟型的毛壶为例说明(图1)。
毛壶体形如花瓶,辐射对称,体四周的薄壁围绕着中央的海绵腔(原腔),体壁内层覆以排列疏松的领细胞,外层为薄的扁平细胞。
体壁穿有无数小孔,称入水孔,顶端开口为出水孔。
基部的组织将毛壶附着于适合的物体上。
体壁内、外层之间为胶状基质,内有几种能游动的变形细胞。
构造较复杂的双沟型是由单沟型多孔动物的体壁呈管状向外辐射皱折而形成,原来成片的领细胞层分割成许多辐管,辐管内的领细胞和海绵腔被隔开。
复沟型是大多数多孔动物具有的水沟系,是双沟型具有领细胞的辐管被无数次地皱折、分割,最后领细胞被限制于小的鞭毛室内,海绵体可不断加厚,形成各种各样的体形(图2)。
组织细胞多孔动物的细胞已初步分化为几种不同功能的组织,但组织中细胞与细胞间并没有严密的关系。
扁平细胞相当于高等动物的表皮细胞,但它只有一层,覆盖着海绵体的表面和体内的水沟系表面,从正面看为多角形,中央较厚,有一大核;侧面看常呈“丁”字形,具有一定的伸缩性。
多孔动物门(海绵动物门)1.体型:体形大多不对称2.胚层:身体由两层细胞组成(皮层和胃层) ,之间为中胶层。
只有细胞分化,没有胚层和组织分化,没有明确的组织以及器官、系统.3.消化:具有特殊的水沟系统水沟系统从单沟型到双沟型到复沟型,领细胞数量不断增多,增加了水流和领细胞的接触面积,提高了食物和氧气的摄取效率4.生殖:胚胎发育过程中有逆转现象(动物极小细胞内陷形成内层,植物极大细胞形成外层)无性生殖–出芽: 体壁向外突起形成芽体,芽体与母体脱离形成新的个体–形成芽球: 原细胞聚集成堆,外保几丁质膜和骨针,形成芽球。
成体死亡后,芽球可度过不良环境有性生殖–雌雄同体或异体。
精子和卵都由原细胞发育而成。
卵在中胶层,领细胞吞食精子后失去鞭毛和领,成变形虫状,将精子带入卵中,使卵受精.5.再生能力强腔肠动物门1.体型:身体辐射对称:辐射对称的体形只有上下之分,没有前后左右之分。
适应于水中固着或漂浮生活。
2.胚层:身体由二个胚层组成,中间为中胶层腔肠动物第一次出现胚层分化,是真正的两胚层动物–外胚层:外层体壁(皮层),具保护、运动和感觉功能–内胚层:内层(胃层),具消化、营养功能–中胶层:内、外胚层细胞分泌的胶状物质。
具有支持的作用3.组织器官:有原始的组织分化–原始的上皮组织:上皮细胞含有肌原纤维,具有上皮和肌肉两种功能,称为上皮肌肉细胞(皮肌细胞)。
既是上皮细胞,又是原始的肌肉细胞–出现原始的神经组织:由各种类型的神经细胞构成弥散型的网状神经系统(原始性表现: 无神经中枢、传导无方向性、传导速度慢)4.消化:出现消化腔相当于高等动物的消化道,消化食物的场所。
相当于胚胎发育过程中的原肠腔–通过腺细胞分泌消化液,食物在消化腔内进行初步消化,是动物进化过程中最早出现细胞外消化(多孔动物:中央腔没有消化作用)–消化腔又具有循环的作用,可把消化后的营养物质输送到身体各部分,故也称为消化循环腔。
–消化腔只有一个对外开口,是原肠期的原口形成的,兼有口(摄食)和肛门(排遗)两种功能5.一般有水螅型和水母型两种基本形态水螅型:适应于固着生活水母型:适应于漂浮生活6.群体多态现象:群体有两种或两种以上具不同形态的体型,有不同的结构,完成不同的生理机能,使群体成为一个完整的整体。
多孔动物门主要特征简介多孔动物门(Porifera)--海绵动物界的一门。
大约5,000种原始多细胞水生动物的统称。
除针海绵属(Spongilla)约20种为淡水产外,多孔动物门均分布在海洋的潮间带到8,500公尺(28,000余尺)深处,营固着生活。
由于海绵常呈分枝形,而且不会移动,从前被人们当作植物。
多孔动物为原始的多细胞动物,本动物门也称海绵动物门(Spongiatia),一般称之为海绵(Sponge)。
海绵是重要的造礁生物,少数属种也有一定的地层意义。
形态构造多孔动物的外形变化很大,除少数种类外,往往没有对称面,在许多方面与低等植物相似,常被描述为块状、垫状、球状、指状、树枝状、杯状或漏斗状等。
一般来说,深海种类的身体常趋于对称,具柄状体,固着在海底软泥上,由一个或成束的强大骨针形成柄或轴,下端深入泥中,上端将海绵体高高托起。
有的种类基部有须根状的骨针,将海绵固着在海底上。
多孔动物的基本构造,可用单沟型的毛壶为例说明(图1)。
毛壶体形如花瓶,辐射对称,体四周的薄壁围绕着中央的海绵腔(原腔),体壁内层覆以排列疏松的领细胞,外层为薄的扁平细胞。
体壁穿有无数小孔,称入水孔,顶端开口为出水孔。
基部的组织将毛壶附着于适合的物体上。
体壁内、外层之间为胶状基质,内有几种能游动的变形细胞。
构造较复杂的双沟型是由单沟型多孔动物的体壁呈管状向外辐射皱折而形成,原来成片的领细胞层分割成许多辐管,辐管内的领细胞和海绵腔被隔开。
复沟型是大多数多孔动物具有的水沟系,是双沟型具有领细胞的辐管被无数次地皱折、分割,最后领细胞被限制于小的鞭毛室内,海绵体可不断加厚,形成各种各样的体形(图2)。
组织细胞多孔动物的细胞已初步分化为几种不同功能的组织,但组织中细胞与细胞间并没有严密的关系。
扁平细胞相当于高等动物的表皮细胞,但它只有一层,覆盖着海绵体的表面和体内的水沟系表面,从正面看为多角形,中央较厚,有一大核;侧面看常呈“丁”字形,具有一定的伸缩性。
孔细胞很特殊的海绵细胞。
分布于体壁上,圆柱形,中央有一孔,实际上形成管状的细胞,有一层薄的原生质膜覆盖孔的外端,孔细胞将海水引入体内,细胞有高度的伸缩性能调节水流。
领细胞圆或椭圆形细胞,整齐地覆盖着鞭毛室的壁,鞭毛和领不断运动,能使体内的水流动,从而自水中获得食物并进行其他生理过程。
变形细胞在扁平细胞和领细胞之间,充满胶状体,称中胶层。
有许多变形细胞能分化为各种具有特殊功能的细胞,如造骨细胞、领细胞等;未分化的变形细胞能贮存食物和色素等。
生殖细胞在生殖期内,领细胞能演变为生殖细胞。
造骨细胞有一类变形细胞专门分泌制造各种骨针。
小骨针常由一个变形细胞分泌制造,大骨针则由几个或许多变形细胞共同分泌制造。
根据骨针性质的不同,造骨细胞可分为钙质造骨细胞、硅质造骨细胞和海绵质造骨细胞等。
此外,还有能分泌粘液的腺细胞(gland cell),类似其他无脊椎动物肌肉细胞的肌原细胞(myocyte)等。
骨针和骨骼多孔动物的骨针有晶体骨针和丝或网状的纤维两种形态。
一般晶体骨针中心有一根有机质构成的轴丝,轴丝表面沉积着碳酸钙或氧化硅;钙质海绵只有钙质骨针,六放海绵只有硅质骨针;寻常海绵具有硅质骨针或海绵质纤维,或两者兼有;硬骨海绵兼有硅质和钙质构成的骨针以及海绵质纤维。
骨针的化学性质可作为多孔动物门各纲的分类依据。
骨针是根据其轴或辐来分类,一般可分为:单轴骨针、三辐骨针、四辐骨针、五辐和六辐骨针、多辐骨针和球状骨针等几种(图3)。
骨骼中的矿物质含量占海绵体重的大部分。
生理多孔动物没有特别分化的消化、排泄和呼吸等器官,其生理过程都是由各种细胞和水流接触时各自直接进行,它的排泄物直接排到水流中。
鞭毛室里千百万个领细胞的鞭毛作有规则的螺旋式运动,激动着水朝一个方向流动,使其从入水孔进入鞭毛室和其他管道,最后从出水口流出体外。
多孔动物是无选择过滤食性动物,食物是由流入体内的水携带而来。
在实验室观察到它们能摄取细菌、淀粉、奶粉、洋红等颗粒,并测得它能吞食的最大食物直径达50微米。
原生质构成的触手状领将食物颗粒附着,随后推入领细胞内进行消化,或由领细胞传给临近的变形细胞并在变形细胞内消化。
再生多孔动物具有很强的再生能力,同类多孔动物的身体紧密接触时,常出现彼此组织互相愈合的现象。
生殖多孔动物能进行有性和无性生殖,雌雄同体,雌性和雄性细胞往往不同时成熟,这样可以避免自体受精。
多孔动物没有特化的生殖腺,生殖细胞(即卵和精子)由领细胞演变而成,演变时领和鞭毛消失,细胞移入领细胞层内,作变形运动,由营养细胞供应所需的营养。
卵在体内受精并进行卵裂,各种多孔动物的生殖期尚不清楚。
胚胎发生受精卵有规律地分裂到32个细胞(囊胚)以后,卵裂就开始不规律地进行。
囊胚呈卵圆形,一端扁平,一端钝圆。
中央腔通过一端的开口与外界相通,称孔。
在周围的细胞和已经从其他细胞分化出来的细胞内,有许多颗粒内含物,另一部分细胞排列成半球形,有长鞭毛,颗粒细胞迅速增多,陷入中央腔内,同时胚胎以其鞭毛的活动移入最近的鞭毛室,经出水口离开母体。
碗状凹陷的胚胎吸收水分,很快膨大成卵圆形。
这时的多孔动物体称两囊幼体,其中一个囊的细胞具有发亮的色素和长的鞭毛,另一个囊的细胞为圆的、有颗粒的细胞。
所有多孔动物都具两囊幼体期,幼体以其纤毛自由游泳数日后,开始沉入海底,有纤毛的一端附着在适合的物体上,开始附着生活(图4)。
这时,多孔动物有两层细胞,原来具有颗粒的细胞在外层,具有鞭毛的细胞在内层。
不久,鞭毛细胞产生一圈原生质的触手状领,变成领细胞;进入两层之间的颗粒细胞演变成具有各种功能的变形细胞,这时多孔动物的幼体称为厚海绵,随后长大为成体多孔动物。
虽然各种多孔动物发生的具体过程略有不同,但都经过上述各阶段。
无性生殖常以出芽和芽球进行。
这在分布很广的荔枝海绵中常可见到。
芽球在淡水多孔动物中很普通。
芽球离开母体后,次年春天会在适合的环境中发育成新的多孔动物。
海绵动物组织原始,无真正消化腔和神经系统。
海绵动物的细胞虽有分工,但彼此合作甚微,如将海绵磨碎过筛,其中分离了的细胞仍能存活数天(相当于原生动物)。
但若彼此不再结合,就不能继续生存下去,海绵动物这种即独立又合作的特征,表明其有机体结构仍属细胞级,显示了原始多细胞动物的特点。
海绵动物多为群体,单体较少。
身体呈辐射对称或不对称。
群体的外形变化很大。
单体一般作角锥形、盘形、高脚杯形、球形等。
大小变化由数毫米到2m之间。
多数具有钙质、硅质或角质骨骼。
海绵动物的骨骼有骨针(海绵针)、海绵丝(骨丝)和非骨针型的矿物质三种。
骨针成分为钙质(方解石、文石)或硅质(蛋白石)。
骨针按大小可分为大骨针和小骨针。
海绵丝的成分是角质的有机化合物,呈丝状,分枝或交接在一起。
海绵丝易腐烂,不易形成化石。
海绵动物除了个别的科没有骨骼之外,其他所有的种类都是具有骨骼的,骨骼是海绵动物的一个典型特征,是用以分类的重要依据之一。
海绵动物的骨骼有骨针(spicule)及海绵丝(sponginfiber)两种类型,它们或散布在中胶层内,或突出到体表,或构成网架状。
骨骼具有支持及保护身体的功能。
骨针的成分或是由碳酸钙组成钙质骨针,或是由SiO2·Nh2O组成硅质骨针,其中还都可能包括微量的铜、镁、锌等离子。
骨针按其大小又可分为大骨针(megasclere),构成支持身体的骨架;及小骨针(microsclere),它散布在中胶层内,以支持体壁中的管道部分。
小骨针仅存在于硅质海绵中。
从形态上骨针可以分为多种,其中常见的有:(1)单轴骨针(monaxons),即沿一个轴生长形成的骨针,轴或直或弯,轴的两端或相似或不相似,末端或尖或具有其他改变;(2)四轴骨针(tetraxons),也称四放骨针(quadriradiate),这种骨针在一个平面上有四个放射端,但常因丢失一些放射端而变成三放、二放或一放型,三放骨针是钙质海绵纲动物中最普通的一种骨骼;(3)三轴骨针(triaxons),它的三个轴相互以直角愈合,因而呈六放型(hexactinal),这种也常减少末端而改变放数,其末端可以弯曲、分枝、或具钧、具结等变化而形成了多种形态;(4)多轴骨针(polyaxons),由中心向外伸出多射,形成星壮,这种类型多见于小骨针。
不同种的海绵,各种骨针或彼此分离,或按一定结构形成疏松的或坚实的网架以支持身体,因此可根据骨针的类型、数量及排列而作为海绵动物分类的依据。
海绵丝是一种纤维状骨骼,它是由硬蛋白(scleroprotein)组成,它们或单独的存在于海绵动物体壁内,或与硅质骨针同时存在。
许多小的硅质骨针埋在海绵丝中,形成有效地支持物。
许多大型群体海绵常同时存在着这两种骨骼。
海绵动物的骨针及海绵丝都是由中胶层中的变形细胞特化形成的造骨细胞所形成。
单轴的钙质骨针是由一个造骨细胞分泌形成,骨针形成时,造骨细胞核先分裂,并在双核细胞的中心出现一个有机质的细丝,然后围绕这一细丝沉积碳酸钙,随着骨针的逐渐增长,双核细胞也分成两个细胞,并分别加长骨针的两端,最后形成一个单轴骨针。
同样,三轴骨针是由三个造骨细胞聚集在一起,每个细胞也随着有机质细丝的形成而分裂一次,形成六个细胞,碳酸钙围绕有机质细丝沉积愈合的结果形成了一个三轴型骨针。
海绵丝是由许多造骨细胞联合形成,先是由少数细胞形成分离的小段,然后再愈合成长的海绵丝。
在寻常海绵纲动物中,这些海绵丝再相互联结形成网状骨架。
海绵的生殖有无性和有性两种。
现代海绵除普通海绵纲中少数类型属淡水海绵外,多数是海生动物,营底栖固着生活。
现代石海绵和钙质海绵多分布于浅海地带,但玻璃海绵可栖居在深达6000m的深海中。
化石海绵也大体要求相似的水深。
海绵在不同的地质时代常和层孔虫、苔藓虫和藻类在一起形成礁体。
分类依据骨骼成分及水沟系类型,可分为以下:·普通海绵纲(Demospongea)·玻璃海绵纲(Hyalospongea)·钙质海绵纲(Calcispongea)·硬海绵纲(Sclerospongea)普通海绵纲(Demospongea)普通海绵纲本纲骨骼为硅质骨针或海绵丝,或两者都有。
骨针通常是互成60o角或120o角度,四轴针、单轴针或网结针,没有六射三轴针。
水沟系为紧密的复沟型。
寒武纪至现代。
Archaeoscyphia Hinde(古钵海绵)玻璃海绵纲玻璃海绵纲(Hyalospongea)又称六射海绵纲(Hexactinellida),形体大,骨骼全由硅质骨针组成,无海绵丝。
典型的大骨针多为六射三轴针,互成直角,故又称六射海绵纲。
此外还有四射双轴针等。
骨针往往连接形成立体格架。
小骨针呈六射星状或双盘状。
简单复沟型,鞭毛室大;某些古生代种类还有单沟型。