Effect of planting geometry and weed management practices on crop growth and yield
- 格式:pdf
- 大小:6.23 MB
- 文档页数:6

类词汇|新托福分类词汇下载新托福分类词汇|新托福分类词汇下载part 1.天文类1 astronom y 天文astronom y 天文学astronom ical 天文的astronom ical observat ory 天文台astronom er 天文学家astrophy sics 天文物理学astrolog y 占星学pseudosc ience 伪科学cosmos(u niverse)宇宙cosmolog y 宇宙学infinite 无限的cosmic 宇宙的radiatio n 宇宙辐射cosmic rays 宇宙射线celestia l 天的celestia l body (heavenl y body)天体celestia l map (sky atlas)天体图celestia l sphere 天球dwarf (dwarf star) 矮星quasar 类星体,类星射电源constell ation 星座galaxy (Milky Way) 新河系cluster 星团solar system 太阳系solar corona 日冕eclipse 日食solar radiatio n 太阳辐射planet 行星planetoi d (asteroi d) 小行星revolve 旋转,绕转twinkle 闪烁naked eye 肉眼Mercury 水星Venus 金星Earth 地球Mars 火星Jupiter 木星Saturn 土星Uranus 天王星Neptune 海王星Pluto 冥王星orbit 轨道spin 旋转satellit e 卫星lunar 月球的流星meteor shower 流星雨meteoroi d 流星体meteorit e 陨石comet 彗星space (outer space)太空,外层空间spacecra ft (spacesh ip) 宇宙飞船spaceman ( astronau t) 宇航员,航天员space suit 宇航服,航天服space shuttle 航天飞机space telescop e 空间望远镜astronau t 宇航员star 恒星stellar 恒星的intergal actic 星系间的llar 恒星间的interpla netary 行星间的asteroid 小行星nebula 星云space debris 太空垃圾ammonia 氨corona 日冕chromosp here 色球photosph ere 色球层convecti on zone 对流层vacuum 真空infrared ray 红外线chondrit e 球粒absolute magnitud e 绝对量级radiatio n 辐射emission 发射,散发high-resoluti on 高清晰度ometer 干扰仪新托福分类词汇|新托福分类词汇下载2 geograph y 地理geograph y 地理geograph er 地理学家hemisphe re 半球meridian 子午线,经线parallel 平行圈,纬线latitude 经度longitud e 精度elevatio n 海拔altitude 高度temperat e latitude s 温带地区horizon 地平线equator 赤道tropics 热带地区Arctic 北极c(Antarc tica) 南极expediti on 探险time zone 时区topograp hy 地形,地形学plain 平原plateau (highlan d) 高地lowland 低地basin 盆地cavern (cave)洞穴terrain 地域subterra nean ( undergro und) 地底下coastlan d 沿海地区island 岛屿continen tal island 大陆岛volcanic island 火山岛coral island 珊瑚岛岛peninsul ar 半岛continen t 大陆continen talshelf 大陆架ranges 山脉valley 峡谷canyon 峡谷channel (strait)海峡remote-sensing 遥感的terrestr ial 地球的,陆地的terrestr ial heat (geother mal) 地热terrestr ial magnetis m (geomagn etism)地磁continen taldrift 大陆漂移学说sea-floor spreadin g 海床扩展evaporat ion 蒸发salinity 含盐度ocean bottom 海床sediment 沉淀物,沉积物tropical 热带的temperat e 温带的frigid 寒带的formatio n 形成frost heaving 冻胀现象fieldsto ne 卵石新托福分类词汇|新托福分类词汇下载3 geology 地质学geologis t 地质学家crust 地壳mantle 地幔core 地核continen tal crust 大陆地壳oceanic crust 海洋地壳layer (stratum , 复数strata)地层plate 板块fault 断层fault plane 断层面fault zone 断层带rift (crack; split)断裂disinteg ration (decompo sition)分解erosion 腐蚀fossil 化石igneous rock 火成岩sediment ary rock 沉积岩metamorp hic rock 变质岩quartz 石英e 石灰岩marble 大理石granite 花岗岩lithogen ous 岩成的lithosph ere 岩石圈magma 岩浆mineral 矿物ore 矿石deposit 矿床platinum 白金,铂silver 银copper 黄铜aluminum 铝tin 锡lead 铅zinc 锌nickel 镍mercury 汞,水银sodium 钠gem 宝石diamond 钻石emerald 绿宝石ruby 红宝石glacier 冰川glacial 冰川的epoch ( age, period)冰川期glacial drift 冰渍iceberg 冰山volcano 火山active volcano 活火山extinct volcano 死火山dormant volcano 休眠火山eruption 火山喷发crater 火山口lava 火山岩浆volcanic 火山的volcanic dust 火山尘volcanic ash 火山灰earthqua ke ( quake; tremor; seism)地震seismic 地震的seismolo gy 地震学e 震级seismic wave 地震波cataclys m 灾变新托福分类词汇|新托福分类词汇下载4 anthropo logist 人类学家paleoant hropolog ist 古人类学家ecologic al anthropo logist 生态人类学家psycholo gical anthropo logist 心理人类学家origin 起源originat e 起源于ancestor 祖先hominid 人(科)homogene ous 同一种族(种类)的tribe 部落族archeolo gist 考古学家excavati on 挖掘excavate (unearth ) 挖掘ruins 遗迹,废墟remains 遗迹,遗骸artifact 手工艺品relic 遗物,文物antique 古物,古董antiquit y 古代,古老Stone Age 石器时代Bronze Age (青)铜器时代Iron Age 铁器时代Paleolit hic 旧石器时代的Mesolith ic 中石器时代的Neolithi c 新石器时代的archaeol ogy 考古学logy 人类学morpholo gy 形态学skull 颅骨cranial 颅骨的新托福分类词汇|新托福分类词汇下载5 ecology 生态学ecologis t 生态学家ecosyste m 生态系统balance (of nature)自然界生态平衡fauna 动物群flora 植物群rain forest 雨林food chain 食物链acidrain 酸雨greenhou seeffect 温室效应radiatio n 红外线辐射ozone layer (ozonosp here) 臭氧层ultravio let radiatio n 紫外辐射pollutio n 污染pollutio ncontrol 污染控制air pollutio n 空气污染water pollutio n 水污染noise pollutio n 噪音污染soil pollutio n 土壤污染pollutio n-free 无污染pollutan t 污染物noxious (toxic)有毒的fumes (有毒)废气物solid waste 固体垃圾sewage (wastewa ter) 污水sewage purifica tion 污水净化sewage disposal 污水处理decibel (噪音)分贝新托福分类词汇|新托福分类词汇下载6 meteorol ogy 气象meteorol ogy 气象meteorol ogist 气象学家forecast (predict ) 预报climate 气候atmosphe re 大气层troposph ere 对流层current (气)流汽evaporat e 蒸发damp (wet; moist; humid)潮湿的humidity 湿度moisture 潮湿;水分saturate 饱和dew 露frost 霜fog (mist)雾smog 烟雾droplet 小水珠condense 浓缩crystal 水晶体downpour (torrent ial rain) 大雨tempest (storm)暴风雨drizzle 细雨shower 阵雨hail 冰雹blizzard (snowsto rm) 暴风雪e (snowsli de) 雪崩precipit ation (雨、露、雪等)降水breeze 微风gale 大风whirlwin d 旋风typhoon 台风hurrican e 飓风tornado (twister , cyclone)龙卷风funnel 漏斗,漏斗云disaster (calamit y, catastro phe) 灾难devastat ion 破坏submerge 淹没drought 旱灾新托福分类词汇|新托福分类词汇下载7 chemistr y 化学chemical property 化学特性,化学性质chemical composit ion ( makeup )化学成分chemical agent 化学试剂chemical reaction 化学反应chemical change 化学变化chemical bond 化学键chemical apparatu s 化学器械substanc e (matter; material ) 物质element 元素periodic table 周期表hydrogen 氢oxygen 氧nitrogen 氮helium 氦碳calcium 钙silicon 硅sulfur 硫iodine 碘compound 化合物hydrocar bon 碳氢化合物derivati ve 衍生物alchemy 炼金术petroleu m (oil)石油petroleu m products 石油产品crudeoil 原油refine 提炼,精炼gasoline 汽油methane 甲烷solution 溶液dissolve 溶解solvent 溶剂solubili ty 可溶性cohesive粘合力atom 原子nucleus 原子核electron 电子neutron 中子proton 质子molecule 分子particle 粒子ion 离子particle accelera tor 粒子加速器catalysi s (复数catalyse s ) 催化作用catalyst 催化剂artifici al 人造的syntheti c 合成的syntheti c fiber 人造纤维polymer 聚合物polymeri zation 聚合作用plastic 塑料dye 染料cosmetic s 化妆品类词汇|新托福分类词汇下载8 .physics 物理physics 物理mechanic s 力学thermody namics 热力学acoustic s 声学electrom agnetism 电磁学optics 光学dynamics 动力学force 力velocity 速度accelera tion 加速度equilibr ium 平衡statics 静力学motion 运动inertia 惯性gravitat ion 引力relativi ty 相对gravity 地心引力vibratio n 震动(media)媒质frequenc y 频率waveleng th 波长pitch 音高intensit y 强度echo 回声resonanc e 回声,洪亮sonar 声纳ultrason ics 超声学electric ity 电static electric ity 静电magnetis m 磁性,磁力magnet 磁体electrom agnet 电磁magnetic field 磁场electric current 电流direct current (DC) 直流电ing current (AC) 交流电electric circuit 电路electric charge 电荷electric voltage 电压electric shock 触电electric applianc e 电器conducto r 导体insulato r 绝缘体semicond uctor 半导体battery (cell)电池dry battery 干电池storage battery 蓄电池electron ics 电子学electron ic 电子的electron ic componen t (part)edcircuit 集成电路chip 集成电器片,集成块electron tube 电子管vacuum tube 真空管transist or 晶体管amplific ation (名词 )放大amplify (动词)放大amplifie r 放大器,扬声器oscillat ion 震荡optical 光(学)的optical fiber 光学纤维lens 透镜,镜片convex 凸透镜concave 凹透镜microsco pe 显微镜telescopr 放大镜spectrum 光谱ultravio let 紫外线X rays X 射线Gamma rays γ射线infrared rays 红外线microwav es 微波dispersi on 色散transpar ent 透明transluc ent 半透明opaque 不透明的新托福分类词汇|新托福分类词汇下载9 mathemat ics mathemat ics 数学mathemat ician 数学家arithmet ic 算术calculat ion 计算calculat or 计算算盘numeral 数字sum 和total (sum total)总和,总数aggregat e 总计fraction 分数,小数decimal 十进位,(十进)小数digit 数字,数位percenta ge 百分比addition 加法plus 加上subtract ion 减法minus 减去multipli cation 乘法multipli cation table 乘法表multiply 乘以times 乘以power 乘方square除法divide 除以algebra 代数quantity 表示量的数(或符号)equation 方程式,等式formula 公式root 根geometry 几何geometri c 几何的dimensio n 维parallel adj. 平行的 n.平行线quadrila teral 四边形square 正方形area 面积rectangl e 长方形,矩形polygon 多边形cube 立方形volume 体积sphere 球形circle 圆形(oval)椭圆形cone 圆锥形circumfe rence 圆周diameter 直径radius 半径triangle 三角形angle 角degree (角 ) 度calculus 微积分differen tial calculus 微分学integral calculus 积分学function 函数probabil ity 概率statisti cs 统计quantita tive 量的,数量的qualitat ive 质的,质量的deductio n (inferen ce) 推论,演绎inductio新托福分类词汇|新托福分类词汇下载10 agricult ure 农业agricult ure (farming ) 农业agricult ural 农业的land (soil)土壤;土地soil conserva tion 土壤保护soil erosion 泥土流失silt 粉砂,泥沙clay 黏土,湿土clod 土块agrarian 土地的,农业的pilot (experim ental )试验性的harrow( rake)耙,耙土ridge 田埂furrowpatch )小块地ranch 农场,牧场plantati on 种植园orchard 果园nursery 苗圃seedbed 苗床sickle 镰刀spade 铲,锹shovel (平头 )铲pick 稿tractor 拖拉机cultivat e (till)耕作sow(seed ) 播种harvest 收割weed 除草irrigate 灌溉manure(f ertilize r) 肥料spray 喷洒(农药)insectic ide(pest icide)杀虫剂pest 害病grain (cereal)谷物,谷粒granary( grain store)粮仓mill 碾,磨wheat 小麦corn 玉米rice 大米barley 大麦sorghum 高粱oats 燕麦rye 黑麦millet 粟,小米vegetabl e 蔬菜horticul ture 园艺学hydropon ics 水栽法,营养液栽培法greenhou se(glass house, hotbed)温室cabbage 洋白菜lettuce 生菜mustard菠菜broccoli 花椰菜cucumber 黄瓜eggplant 茄子pepper 辣椒pumpkin 南瓜tomato 西红柿beet 甜菜carrot 胡萝卜radish 小红萝卜pea 豌豆soybean 大豆celery 芹菜garlic 大蒜leek 韭菜onion 洋葱头potato 土豆peanut 花生sesame 芝麻cotton 棉花husbandr y (animal husbandr y) 畜牧业pasturek 家畜fowl (poultry ) 家禽cattle 牛,家畜buffalo 野牛dairy (dairy cattle)奶牛dairy farm 乳牛场hay (作饲料用)干草haystack 干草堆fodder (feed)饲料trough 饲料槽barn(she d) 牲口槽stable 厩,马厩cowshed 牛棚pigpen (hog pen, pigsty)猪圈sheepfol d (sheep pen) 羊栏roost (hen house)鸡舍fish farm 养鱼场aquacult ure 水产养殖新托福分类词汇|新托福分类词汇下载11 animal 动物Darvinis m 达尔文学说natural selectio n 自然选择class 纲order 目family 科genus 属suborder 亚目species 种inverteb rate 无脊椎动物vertebra te 脊椎动物aquatic( life) 水生动物reptile 爬行动物n (amphibi ous animal)两栖动物dinosaur 恐龙bird 鸟类extincti on 灭绝mammal 哺乳动物primates 灵长目动物insect 昆虫antenna ( 复数antennae ) 触须larva 幼虫,幼体camoufla ge 伪装pest 害虫worm 虫,蠕虫hibernat e 冬眠lizard 蜥蜴chameleo n 变色蜥蜴regenera tion 再生turtle 龟beast 野兽domesticy (carnivo rous )食肉的predator 捕食者prey (动词)捕食(名词)被捕食的动物scavenge r 食腐动物migrate 迁移wing 翅膀,翼bill (鸟)嘴beak (鹰等的)嘴nest 筑巢dolphin 海豚whale 鲸鱼bat 蝙蝠gorilla 大猩猩chimpanz ee 黑猩猩habitat 栖息地rodent 啮齿动物(如松鼠)primate 灵长动物plankton 浮游生物软体动物coelente rate 腔肠动物(如珊瑚)shrimp 小虾prawn 对虾lobster 对虾crab 螃蟹clam 蛤蜊sponge 海绵coral 珊瑚starfish 海星canary 金丝雀chirp (鸟,虫的叫声 )唧唧squeak (老鼠等)吱吱porpoise 海豚sloth 树懒slothful 懒惰的hygiene 卫生sanitati on 卫生parasite 寄生虫moth 蛾caterpil昆虫等 )群swarms (昆虫等 )群flock (鸟,羊等)群communit y 动物的群落或人的部落herd 兽群bunch (花等)束,捧beaver 海狸monogamo us 一夫一妻的,一雄一雌的polygamo us 一夫多妻的,一雄多雌的polyandr ous 一妻多夫的,一雌多雄的trapper 诱捕动物者rhinocer os 犀牛niche 小生态环境vestige 退化器官,遗迹oysterer 使 …受精larvae 幼虫tentacle 触角homother m 恒温动物poikilot herm 变温动物metaboli sm 新陈代谢baboon 狒狒breed (名词 )品种,(动词)繁殖multiply (reproduc e )繁殖hatch 孵spawn (鱼、虾、蛙等 )孵offsprin g (young)解剖学anatomy 解剖学appetite 食欲creature 生物microbe 微生物herbivor ous 食草的ous 食肉的omnivoro us 杂事的新托福分类词汇|新托福分类词汇下载12 plant 植物botany 植物学botanist 植物学家botanica l (botanic ) 植物的plant 植物aquatic plant 水生植物parasite plant 寄生植物root 根canopy 树冠层,顶棚foliage (leaf)叶leaflet 小叶rosette ( 叶的 )丛生stem 茎stalk 杆leafstal k 叶柄shoot (sprout ) 嫩芽,抽枝flower 花bud 花蕾petal 花瓣peel(ski n) 果皮shell (硬 ) 果壳husk (干)果壳;(玉米)苞叶trunk 树干branch 树枝bough 大或者粗的树枝twig 小树枝bark 树皮jungle 丛林lawn 草坪meadow 草地,牧场prairie 大草原shrub (bush)灌木cluster 一簇(灌木)fern 蕨类植物herb 草thesis 光合作用symbiosi s 共生wither (shrivel , fade)凋谢pollen 花粉pollinat e 传授花粉pollinat ion 授粉cell 细胞tissue 组织organ 器官system 系统seeds 种子everlast ing 永久的orchid 兰花pollinat e 授粉crossbre ed 杂交shrub 灌木sequoia 红杉root pressure 根压cohesion -tension 凝聚压力bore 腔,肠花柱symbioti c 共生的immunity 免疫organism 生物,有机物necrosis 坏死barren 贫瘠futile 无用的carbohyd rate (starch)碳水化合物(淀粉),糖类glucose 葡萄糖starch 淀粉fat 脂肪protein 蛋白质vitamin 维他命calorie 卡路里(热量单位)malnouri shed 营养不良的nutritio n 营养新托福分类词汇|新托福分类词汇下载13 politics 政治constitu tion 宪法legislat ion 立法Democrat s 民主党Republic ans 共和党Amendmen t 修正案Congress 美国国会Senate 参议院House of Represen tatives 众议院monarchy 君主制anarchis m 无政府主义federal system 联邦制presiden tial system 总统制municipa l 市的,市政的metropol itan 大都市的centrali zed 中央集权的ideology 意识形态擦略tariff 关税immigran t 移民institut ionalize 机构化election 选举vote 投票officeho lding 任职veto 否决American Revoluti on 美国革命 /独立战争Independ ence War 美国独立战争American Civil War 美国内战radical 激进的 ,根本的overturn ing 颠覆性的independ ent 独立的breathta king 激动人心的military 军事的拓殖,殖民per capita 人均treason 叛国conspira cy 阴谋imprison ment 监禁release 释放pardon 特赦accusati on 谴责,指控welfare 福利patronag e 资助,赞助unioniza tion 联合,结合新托福分类词汇|新托福分类词汇下载14 educatio n 教育educator 教育家higher educatio n 高等教育moral characte r 道德品质领域faculty 全体教员alumni 校友treasure r 生活委员,财务大臣elective system 选课制度curricul um 课程(总称)discipli ne 学科,管理learn by rote 死记硬背15 music 音乐musical instrume nt 乐器orchestr a 乐队string 弦乐wind 管乐shook rattle 摇拨浪鼓pound drum 击鼓footbeat 点脚,跺脚percussi on 震荡,打击乐器符score 乐谱movement 乐章fanatica l 狂热的hillbill y music 乡村音乐folk music 民间音乐pop music 流行音乐classica l music 古典音乐Jazz 爵士乐16 psycholo gy 心理mental 心理的physical 身体的,物质的,物理的spiritua l 心灵的conformi ty 从众majority 多数人minority 少数人threshol d judgment (心理学)初始性判断。
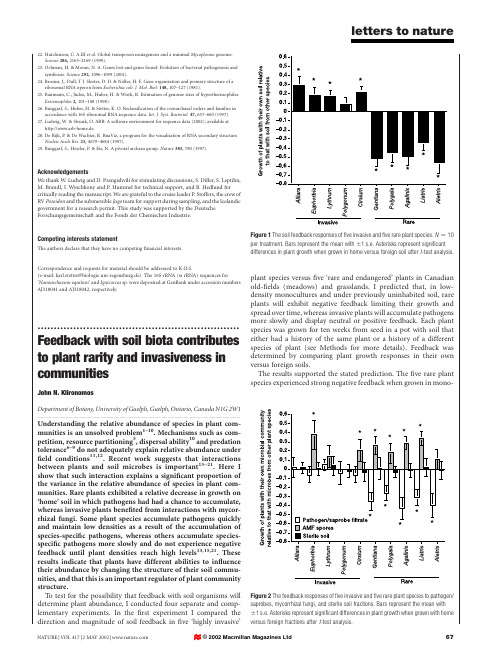
22.Hutchinson,C.A.III et al.Global transposon mutagenesis and a minimal Mycoplasma genome.Science286,2165–2169(1999).23.Ochman,H.&Moran,N.A.Genes lost and genes found:Evolution of bacterial pathogenesis andsymbiosis.Science292,1096–1099(2001).24.Brosius,J.,Dull,T J.Sleeter,D.D.&Noller,H.F.Gene organization and primary structure of aribosomal RNA operon from Escherichia coli.J.Mol.Biol.148,107–127(1981).25.Baumann,C.,Judex,M.,Huber,H.&Wirth,R.Estimation of genome sizes of hyperthermophiles.Extremophiles2,101–108(1998).26.Burggraf,S.,Huber,H.&Stetter,K.O.Reclassification of the crenarchaeal orders and families inaccordance with16S ribosomal RNA sequence data.Int.J.Syst.Bacteriol.47,657–660(1997). 27.Ludwig,W.&Strunk,O.ARB:A software environment for sequence data(2002);available athttp://www.arb-home.de.28.De Rijk,P.&De Wachter,R.RnaViz,a program for the visualisation of RNA secondary structure.Nucleic Acids Res.25,4679–4684(1997).29.Burggraf,S.,Heyder,P.&Eis,N.A pivotal archaea group.Nature383,780(1997).AcknowledgementsWe thank W.Ludwig and D.Prangishvili for stimulating discussions,S.Diller,S.Leptihn, M.Brandl,I.Wyschkony and P.Hummel for technical support,and B.Hedlund for critically reading the manuscript.We are grateful to the cruise leader P.Stoffers,the crew of RV Poseidon and the submersible Jago team for support during sampling,and the Icelandic government for a research permit.This study was supported by the Deutsche Forschungsgemeinschaft and the Fonds der Chemischen Industrie.Competing interests statementThe authors declare that they have no competingfinancial interests.Correspondence and requests for material should be addressed to K.O.S.(e-mail:karl.stetter@biologie.uni-regensburg.de).The16S rRNA(ss rRNA)sequences for‘Nanoarchaeum equitans’and Ignicoccus sp.were deposited at GenBank under accession numbers AJ318041and AJ318042,respectively. .............................................................. Feedback with soil biota contributes to plant rarity and invasiveness in communitiesJohn N.KlironomosDepartment of Botany,University of Guelph,Guelph,Ontario,Canada N1G2W1 ............................................................................................................................................................................. Understanding the relative abundance of species in plant com-munities is an unsolved problem1–10.Mechanisms such as com-petition,resource partitioning5,dispersal ability10and predation tolerance6–9do not adequately explain relative abundance under field conditions11,12.Recent work suggests that interactions between plants and soil microbes is important13–21.Here I show that such interaction explains a significant proportion of the variance in the relative abundance of species in plant com-munities.Rare plants exhibited a relative decrease in growth on ‘home’soil in which pathogens had had a chance to accumulate, whereas invasive plants benefited from interactions with mycor-rhizal fungi.Some plant species accumulate pathogens quickly and maintain low densities as a result of the accumulation of species-specific pathogens,whereas others accumulate species-specific pathogens more slowly and do not experience negative feedback until plant densities reach high levels13,15,21.These results indicate that plants have different abilities to influence their abundance by changing the structure of their soil commu-nities,and that this is an important regulator of plant community structure.To test for the possibility that feedback with soil organisms will determine plant abundance,I conducted four separate and comp-lementary experiments.In thefirst experiment I compared the direction and magnitude of soil feedback infive‘highly invasive’plant species versusfive‘rare and endangered’plants in Canadian old-fields(meadows)and grasslands.I predicted that,in low-density monocultures and under previously uninhabited soil,rare plants will exhibit negative feedback limiting their growth and spread over time,whereas invasive plants will accumulate pathogens more slowly and display neutral or positive feedback.Each plant species was grown for ten weeks from seed in a pot with soil that either had a history of the same plant or a history of a different species of plant(see Methods for more details).Feedback was determined by comparing plant growth responses in their own versus foreign soils.The results supported the stated prediction.Thefive rare plant species experienced strong negative feedback when grown inmono-Figure2The feedback responses offive invasive andfive rare plant species to pathogen/ saprobes,mycorrhizal fungi,and sterile soil fractions.Bars represent the mean with^1s.e.Asterisks represent significant differences in plant growth when grown with home versus foreign fractions after t-testanalysis.Figure1The soil feedback responses offive invasive andfive rare plant species.N¼10 per treatment.Bars represent the mean with^1s.e.Asterisks represent significant differences in plant growth when grown in home versus foreign soil after t-test analysis.culture (reduced growth when growing in their own soil relative to that with soil from other plant species)whereas four of the five invasive plant species showed net positive feedback (increased growth in their own soil,relative to soil of other plant species)(Fig.1).Direct observations were done on plant roots in search for the accumulation of species-specific fungi,but this could not be done with confidence,as it was difficult to distinguish among fungal taxa.Two more experiments were set up to test for the possibility that pathogens and mycorrhizal fungi were the agents responsible for the observed feedback responses in the first experiment.In experiment 2,instead of using the entire soil,only specific microbial fractions were used to re-run the original experiment.The fractions were:(1)spores of arbuscular mycorrhizal fungi (AMF);(2)pathogenic/saprobic microbial filtrate;and (3)sterile soil.Once again,the same ten plant species were grown for ten weeks from seed with one of the soil fractions.The fractions originated from soil containing either a similar or a different plant species (as in experiment 1).Overall,rare plants showed strong negative feedback for the pathogen/saprobe filtrate when grown in soil with a history of the same plant species (Fig.2).In contrast,the origin of the pathogen/saprobe filtrate did not significantly affect growth of the invasive plant species.Effects of AMF,however,did not differ between invasive and rare plants.For both groups of plants,AMF isolated from soil with a history of the same plant species had a more positive effect on plant growth than fungi isolated from a different plant host.There was no significant feedback effect using the sterile soil,suggesting that the effects are biological rather than chemical.A third experiment was conducted to confirm that pathogenic or parasitic organisms were responsible for the negative feedback responses.The three mostcommonlyFigure 3The relationship between relative plant abundance in an old-field site and soil feedback response.Squares represent means.y ¼114.529x 2þ156.652x þ44.013,r 2¼0.575,P ¼0:0001.Numbers represent different plant species.1,Carex garberi ;2,Carex aurea ;3,Carex granularis ;4,Daucus carota ;5,Agrostis gigantea ;6,Solidago graminifolia ;7,Solidago nemoralis ;8,Aster simplex ;9,Aster vimineus ;10,Aster novae-angliae ;11,Circium vulgare ;12,Chenopodium ambrosioides ;13,Oenothera biennis ;14,Carex flava ;15,Juncus dudleyi ;16,Solidago canadensis ;17,Linaria vulgaris ;18,Cichorium intybus ;19,Circium arvense ;20,Solidago rugosa ;21,Geum aleppicum ;22,Satureja vulgaris ;23,Potentilla recta ;24,Coronilla varia ;25,Asclepias syriaca ;26,Achillea millefolium ;27,Apocynum cannabinum ;28,Hypericum perforatum ;29,Agrostis scabra ;30,Phleum pratense ;31,Poa compressa ;32,Echium vulgare ;33,Centaurea jacea ;34,Rudbeckia serotina ;35,Poa pratensis ;36,Dactylis glomerata ;37,Cerastium vulgatum ;38,Galium palustre ;39,Oenothera perennis ;40,Prunella vulgaris ;41,Trifolium pratense ;42,Convolvulus arvensis ;43,Silene cucubalus ;44,Erigeron strigosus ;45,Asparagus officinalis ;46,Hieracium auranticum ;47,Erigeron philadelphicus ;48,Veronica officinalis ;49,Plantago lanceolata ;50,Galium mollugo ;51,Hieracium pilosella ;52,Vicia cracca ;53,Hieracium pratense ;54,Medicago lupulina ;55,Ranunculus acris ;56,Taraxacum officinale ;57,Fragaria virginiana ;58,Chrysanthemum leucanthemum ;59,Tragopogon pratensis ;60,Bromus inermis ;61,Panicum lanuginosum .Table 1Fungi isolated from the following invasive or rare and endangered plant speciesInvasiveRare and endangeredPlant species tested ApEeLsaPcCaGaPiAgLspAf...................................................................................................................................................................................................................................................................................................................................................................Ap 7323213251152922Ee 7282109914510264Lsa 282626616213218*214528Pc 13521522104762410Ca 169171272152822924Ga 11262316218254*571027Pi 2142241275228*1227211Ag 928272219*325246*11216Lsp 321121573121022251*9Al2151112224*245*232517263*...................................................................................................................................................................................................................................................................................................................................................................The influence of the three most commonly isolated non-mycorrhizal fungi from the rhizosphere of each plant species on the growth of each plant species.Values represent percentage difference in growth compared to non-inoculated control plants.Asterisks represent significant differences (P ,0:05)after t -test.Invasive plant species:Ap,Alliara ;Ee,Euphorbia ;Lsa,Lythrum ;Pc,Polygonum ,Ca,Circium .Rare and endangered plant species:Ga,Gentiana ;Pi,Polygala ;Ag,Agalinis ;Lsp,Liatris ;Af,Aletris .isolated non-AMF fungi from the rhizosphere of each plant species were tested for their effects on the growth of each plant species (Table1).With allfive rare plants,significant reductions in growth were observed when they were inoculated with fungi isolated from their own root systems.The most commonly isolated fungi from these plants were species of Verticillium,Fusarium and Cylindro-carpon and,in all cases,they formed systemic infections in the roots of these plants when re-inoculated.No such growth reductions were observed for any of the invasive plants tested.With both groups of plants,inoculations using fungi from foreign plants rarely resulted in significant growth depressions(Table1).Overall,these combined results support the hypothesis that feedback between plants and soil communities may strongly determine the ability of a plant to establish,invade and persist in a local habitat.Most plants can potentially experience positive feedback with AMF communities, but this is not realized unless negative feedback with pathogens is lacking.Thefirst three experiments indicate that soil feedback could be an important mechanism for coexistence and the regulation of plant biodiversity in communities.In the fourth experiment I tested the hypothesis that the direction and degree of soil feedback can account for a large proportion of the variation in relative abundance among species that coexist within a plant community.To test for this I measured soil feedback and relative abundance of each of61 coexisting plant species within an old-field meadow community in southern Ontario,Canada.Relative plant abundance in thefield ranged from1–81%.Also,there was a wide range of feedback responses,from20.5to0.2.The majority of plants were found to experience a net negative feedback,whereas net positive feedback was found in only18%of plant species.As predicted,there was a strong positive relationship between soil feedback and relative abundance in thefield(Fig.3).All plant species that were found to have strong negative feedback were also found to have low relative abundance.Conversely,plants with high abundance had either low negative feedback or positive feedback.Variation in degree of feedback accounted for a large proportion(57.5%)of the variation in relative abundance.The results indicate that soil organisms and their feedback effects on plants can strongly influence the relative abundance of plant species within a community.When introduced to previously uncultured soil at low densities,plants that ultimately achieve high abundance do not seem to accumulate species-specific patho-gens at the same rate as plants that remain in low abundance. Negative feedback responses,particularly reductions in plant growth and spread,are pathogen-density dependent13,15,16,22,so plants that accumulate pathogens at a slower rate can potentially reach higher densities in nature.Plants may accumulate pathogens at a slower rate either because of inherent plant traits22,or because they have escaped their harmful pathogens by invading foreign territory23.Both are likely explanations for the lack of negative feedback in alien invasive plants in this study.Ultimately,even abundant plants are unlikely to expand indefinitely.Plant-specific pathogen loads are maximized under high plant densities,especially under dense monocultures,eventually incurring negative feedback on abundant plants13,15,21,22.Other above-and belowground organ-isms across trophic groups can further interact with feedback responses24,and probably explain a large proportion of the remain-ing variance in relative plant abundance.Understanding the mech-anisms responsible for the abundance of organisms may lead to new approaches for the management of ecosystems,in particular, conservation of rare and endangered species,and protection of ecosystems from species invasions.A MethodsExperiment1Soil feedback in rare versus invasive plant species(Fig.1)was examined as follows:seeds of rare and invasive plant species were collected from variousfield sites across southern Ontario and Quebec,Canada.The rare species were:Agalinis gattingeri(Small)Small (Gattinger’s agalinis),Aletris farinosa L.(colicroot),Gentiana alba Muhl.(white prairie gentian),Liatris spicata(L.)Willd.(dense blazing star)and Polygala incarnata L.(pink milkwort).The invasive species were:Alliara petiolata(Bieb.)Cavara and Grande(garlic mustard),Cirsium arvense(L.)Scop.(Canada thistle),Euphorbia esula L.(leafy spurge), Lythrum salicaria L.(purple loosestrife)and Polygonum cuspidatum Sieb.&Zucc. (Japanese knotweed).Thefive rare species are all native to North America.They are considered to be rare within prairie communities,and they are further threatened because of the loss of prairie habitat due to agricultural expansion and residential development in southern Canada.Thefive invasive plants were all introduced to North America over a century ago from Eurasia.Soil was collected from the Long-Term Mycorrhizal Research Site(LTMRS),located within the University of Guelph Arboretum,Canada.A section of the LTMRS was used that did not contain any of the plant species tested.The experiment had three stages,beginning with200pots,growing the same plant species twice in succession(stages1and2)and then dividing among‘home’and‘foreign’treatments (stage3).In‘home’treatments thefive invasive andfive rare plants were grown in pots with their own respective histories(10replicates each).In‘foreign’treatments they were grown in pots with histories of the other plants(10replicates).The ten foreign replicates were comprised of one replicate with a history of each of the remaining nine plant species tested,as well as one additional species(Taraxacum officinale Weber ex Wiggers).Each experimental unit consisted of a single seedling growing in an eight-inch pot containing field-collected loam soil(total N¼83:1mmol kgÿ1,total P¼6:2mmol kgÿ1,percentage organic matter was6.1),and placed in a random position on a greenhouse bench.Plants were watered on a weekly basis but did not receive additional nutrients.Each plant was allowed to grow for10weeks after which the entire plant was harvested.In the second stage,soil from each pot from thefirst stage was placed into new pots,a new seedling of the same species was grown for another10weeks,and then harvested.In the third stage,soil from each pot from the second stage was placed into new pots,a new seedling was grown for another10weeks(either the same or a different plant species),and then harvested.At harvest,plant weight was determined after drying at608C for48hours.Only plants from the third stage were used to determine feedback with the following equation(for plant A): biomass of plant A(grown in soil with a history of plant A)-biomass of plant A(grown in soil with a history of other plants).For each plant species,comparisons of plant growth in home versus foreign soil were done with Student t-tests.Experiment2Soil feedback in rare versus invasive plant species was determined using AMF,pathogen/ saprobe and chemical fractions(Fig.2).After the second round of harvests from the previous experiment(second stage),two10-g soil subsamples were removed from each pot.Thefirst soil sample was used to extract(1)AMF communities;and(2)soil pathogen/ saprobe communities.AMF spores were collected on a45-m m mesh after wet-sieving25, and surface sterilized with10%sodium hypochlorite.Pathogens and other non-mycorrhizal microbes were collected from thefiltrate that passed through a20-m m sieve. The second soil sample was sterilized by g-irradiation,and used as a control for abiotic effects.To determine AMF,pathogen/saprobe,and abiotic feedback,seedlings of each plant was grown in pots containing sterilized soil and one of the three fractions(AMF, pathogen/saprobes,sterilized soil).Plants were grown for10weeks as in experiment1,and then harvested.After harvest,plant weight was determined after drying at608C for48 hours.There were10replicates for each treatment.Feedback was calculated using the following equations:(1)AMF feedback for plant A¼biomass of plant A(grown in soil with AMF of plant A)2biomass of plant A(grown in soil with AMF of other plants);(2) pathogen/saprobe feedback for plant A¼biomass of plant A(grown in soil with pathogens/saprobes of plant A)2biomass of plant A(grown in soil with pathogens/ saprobes of other plants);(3)Sterile soil feedback for plant A¼biomass of plant A(grown in soil with sterile soil of plant A)2biomass of plant A(grown in soil with sterile soil of other plants).For each plant species,comparisons of plant growth in home versus foreign soil fraction were done with Student t-tests.Experiment3Growth responses of plants to fungi cultured from home versus foreign rhizospheres (Table1)were determined as follows.Atfinal harvest of experiment1,twenty0.5-cm root fragments were plated on a2%malt-extract agar medium containing Rose Bengal26.Root fragments were obtained from each experimental unit in the‘home’treatment and were surface sterilized with0.1%mercuric chloride before plating.Plates were incubated for three weeks at208C,and then all resulting fungal colonies were subcultured andidentified27.A new experiment was set up to test the influence of fungi isolated from each of10plant species on the growth of each of10plant species.Each experimental unit consisted of a single plant seedling growing in an eight-inch pot containing sterilized silica sand and fungal inoculum originating from one of the ten plant species.There were10 replicates per100plant/fungal combinations,as well as ten non-inoculated controls for each plant species,resulting in1,100experimental units.Plant seeds were originally germinated on moist sterilefilter paper.Before being planted in the pots,the roots of one-week-old seedlings were dipped in spore suspensions of the cultured fungi.Spore suspensions were comprised of1£106spores of each of the three most common fungal species isolated from the respective plant species.Experimental units were placed in a random position on a greenhouse bench.Plants were watered as needed,and fertilized on a weekly basis with half-strength Hoagland’s solution.Plants were grown for10weeks as in experiment1,and then harvested.At harvest,plant weight was determined after drying at 608C for48hours.Subsamples of roots were stained with Chlorazol Black E,and inspected with differential-interference contrast and incandescent light microscopy forconfirmation of fungal infection.Plant biomass in inoculated versus non-inoculated plants were compared using Student t-tests.Experiment4Soil feedback and relative abundance of plants in thefield(Fig.3)was measured.The abundance of each plant species was measured at100different locations within the LTMRS.Locations were randomly chosen,and the presence of all plant species within a 1-m2quadrat was recorded at each location.Relative abundance for each species was calculated as the percentage of locations containing that species.This was performed in the summers of1998and2000,and results were pooled.Seeds were collected from each of61 plant species,and feedback response was determined using similar methods as described above in experiment1.Regression analysis was used to determine the relationship between plant abundance and soil feedback.Received6November2001;accepted14February2002.1.Pysek,P.,Prach,K.,Rejmanek,M.&Wade,M.Plant Invasions:General Aspects and Special Problems(SPB Academic,Amsterdam,1995).2.Huston,M.A.Biological Diversity.The Coexistence of Species on Changing Landscapes(CambridgeUniv.Press,Cambridge,1994).3.Ricklefs,R.E.&Schluter,D.Species Diversity in Ecological Communities(Univ.Chicago Press,Chicago,1993).4.Gaston,K.J.Rarity(Chapman&Hall,London,1994).5.Grace,J.B.&Tilman,D.Perspectives on Plant Competition(Academic,San Diego,1990).6.Belsky,A.J.Effects of grazing,competition,disturbance andfire on species composition and diversityin grassland communities.J.Veget.Sci.3,187–200(1992).ton,W.The effect of manuring,grazing and cutting on the yield,botanical and chemicalcomposition of natural hill pastures.J.Ecol.28,326–356(1940).8.Crawley,M.J.in Insect–Plant Interactions(ed.Bernays,E.A.)Vol.1,45–71(CRC Press,Boca Raton,1989).9.Reader,R.J.Relationship between species relative abundance and plant traits for an infertile habitat.Plant Ecol.134,43–51(1998).10.Rabinowitz,D.,Rapp,J.K.,Cairns,S.&Mayer,M.The persistence of rare prairie grasses in Missouri:environmental variation buffered by reproductive output of sparse species.Am.Nat.134,525–544 (1989).11.Tokeshi,M.Species Coexistence:Ecological and Evolutionary Perspectives(Blackwell Science,Oxford,1999).12.Tilman,D.in Biodiversity and Ecosystem Function(eds Schulze,E.-D.&Mooney,H.A.)327–344(Springer,Berlin,1994).13.Van der Putten,W.H.,Van Dijk,C.&Peters,B.A.M.Plant-specific soil-borne diseases contribute tosuccession in foredune vegetation.Nature362,53–56(1993).14.Grime,J.P.,Mackey,J.M.L.,Hillier,S.H.&Read,D.J.Floristic diversity in a model system usingexperimental microcosms.Nature328,420–422(1987).15.Bever,J.D.Feedback between plants and their soil communities in an oldfield community.Ecology75,1965–1977(1994).16.Bever,J.D.,Westover,K.M.&Antonovics,J.Incorporating the soil community into plant populationdynamics:the utility of the feedback approach.J.Ecol.85,561–573(1997).17.Van der Heijden,M.G.A.et al.Mycorrhizal fungal diversity determines plant biodiversity,ecosystemvariability and productivity.Nature396,69–72(1998).18.Packer,A.&Clay,K.Soil pathogens and spatial patterns of seedling mortality in a temperate tree.Nature404,278–281(2000).ls,K.E.&Bever,J.D.Maintenance of diversity within plant communities:soil pathogens as agentsof negative feedback.Ecology79,1595–1601(1998).20.Callaway,R.M.&Ascheloug,E.T.Invasive plants versus their new and old neighbors:a mechanismfor exotic invasion.Science290,521–523(2000).21.Olff,H.,Hoorens,B.,de Goede,R.G.M.,van der Putten,W.H.&Gleichman,J.M.Small-scaleshifting mosaics of two dominant grassland species:the possible role of soil-borne pathogens.Oecologia125,45–54(2000).22.Burdon,J.J.Diseases and Plant Population Biology(Cambridge Studies in Ecology,Cambridge Univ.Press,Cambridge,1987).23.van der Putten,W.H.Pathogen-driven forest diversity.Nature404,232–233(2000).24.Blomqvist,M.M.,Olff,H.,Blaauw,M.B.,Bongers,T.&van der Putten,W.H.Interactions betweenabove-and belowground biota:importance for small-scale vegetation mosaics in a grasslandecosystem.Oikos90,582–598(2000).25.Klironomos,J.N.,Moutoglis,P.,Kendrick,B.&Widden,P.A comparison of spatial heterogeneityof vesicular-arbuscular mycorrhizal fungi in two maple-forest soils.Can.J.Botany71,1472–1480 (1993).26.Bragulat,M.R.,Abarca,M.L.,Bruguera,M.T.&Cabanes,F.J.Dyes as fungal inhibitors:effect oncolony diameter.Appl.Environ.Microbiol.57,2777–2780(1991).27.Domsch,K.H.,Gams,W.&Anderson,pendium of Soil Fungi Vol.1(Academic,London,1980).AcknowledgementsI thank J.Bever,M.Hart,B.Husband and R.Reader for comments on the manuscript.I also thank A.Jones,Pierre,G.Lewis,D.MacIntosh,J.Martin,J.Mosquin,P.Moutoglis and A.Shaw for laboratory andfield assistance.This work was supported by a grant from the Natural Sciences and Engineering Research Council of Canada.Competing interests statementThe author declares that he has no competingfinancial interests.Correspondence and requests for materials should be addressed to J.N.K.(e-mail:jklirono@uoguelph.ca)............................................................... Effects of size and temperature on developmental timeJames.F.Gillooly*,Eric L.Charnov*,Geoffrey B.West†‡,Van M.Savage†‡§&James H.Brown*†*Department of Biology,The University of New Mexico,Albuquerque,New Mexico87131,USA†Santa Fe Institute,1399Hyde Park Road,Santa Fe,New Mexico87501,USA ‡Theoretical Division,MS B285,Los Alamos National Laboratory,Los Alamos, New Mexico87545,USA§Department of Physics,Washington University,St Louis,Missouri63130,USA ............................................................................................................................................................................. Body size and temperature are the two most important variables affecting nearly all biological rates and times1–7.The relationship of size and temperature to development is of particular interest, because during ontogeny size changes and temperature often varies8–12.Here we derive a general model,based onfirst principles of allometry and biochemical kinetics,that predicts the time of ontogenetic development as a function of body mass and temperature.The modelfits embryonic development times spanning a wide range of egg sizes and incubation temperatures for birds and aquatic ectotherms(fish,amphibians,aquatic insects and zooplankton).The model also describes nearly75% of the variation in post-embryonic development among a diverse sample of zooplankton.The remaining variation is partially explained by stoichiometry,specifically the whole-body carbon to phosphorus ratio.Development in other animals at other life stages is also described by this model.These results suggest a general definition of biological time that is approximately invariant and common to all organisms.The effects of body size and temperature on biological rates and times,including development time,have traditionally been studied separately.There is a rich literature on biological allometry that spans nearly a century1–3.The relationships of various attributes of organisms such as metabolic rate,development time and lifespan,to body mass,m,are well approximated by power laws.In endother-mic birds and mammals,where body temperature is nearly con-stant,biological rates and times(t)vary with body size as t/m1=4 (refs4and5).An equally rich literature on physiology relates many。

植树带来的效应英语作文Title: The Effects of Tree Planting。
Tree planting is a practice that holds immense significance in various aspects of our lives and the environment. The act of planting trees not only contributes to the beautification of our surroundings but also brings about a plethora of positive effects that extend far beyond aesthetic appeal. In this essay, we will explore the multifaceted effects of tree planting and its profound impact on the environment, economy, and society.First and foremost, tree planting plays a crucial role in mitigating the adverse effects of climate change. Trees act as natural carbon sinks, absorbing carbon dioxide from the atmosphere during photosynthesis and storing it intheir biomass. By reducing the concentration of greenhouse gases in the atmosphere, trees help alleviate global warming and its associated consequences, such as rising temperatures, erratic weather patterns, and sea-level rise.Therefore, widespread tree planting initiatives are essential in combating climate change and preserving the habitability of our planet for future generations.Moreover, trees play a vital role in maintaining ecological balance and biodiversity. Forests serve as habitats for numerous plant and animal species, providing them with food, shelter, and breeding grounds. The preservation of natural habitats through tree planting efforts is crucial for the conservation of endangered species and the overall health of ecosystems. Additionally, trees help prevent soil erosion by stabilizing the soil with their roots, thereby preserving fertile land for agriculture and reducing the risk of landslides in hilly terrain.Furthermore, tree planting has significant economic benefits for communities and nations. Forests contribute to the economy through the provision of timber, fuelwood, and non-timber forest products such as fruits, nuts, and medicinal plants. The forestry sector also generates employment opportunities in various activities such as treeplanting, forest management, and wood processing industries. Additionally, trees enhance property values and attract tourists, thereby stimulating local economies andsupporting livelihoods.In addition to its environmental and economic benefits, tree planting has positive social impacts as well. Green spaces and urban forests created through tree planting initiatives improve the quality of life in urban areas by providing recreational opportunities, promoting physicaland mental well-being, and reducing air and noise pollution. Trees also have aesthetic value, enhancing the visualappeal of neighborhoods, parks, and streetscapes. Moreover, tree planting projects often foster community engagementand social cohesion as people come together to participatein tree planting events and environmental conservation efforts.In conclusion, tree planting is a powerful tool for addressing a wide range of environmental, economic, and social challenges facing our planet. From mitigatingclimate change and preserving biodiversity to supportingsustainable development and enhancing quality of life, the benefits of tree planting are manifold and far-reaching. Therefore, it is imperative that we continue to prioritize and invest in tree planting initiatives as part of our collective efforts to build a greener, healthier, and more sustainable future for all.。

the effects of light on plant 雅思阅读摘要:I.引言- 光对植物生长的重要性- 光对植物的影响II.光合作用- 光合作用的定义- 光合作用的过程- 光合作用对植物生长的影响III.光的形态建成- 形态建成的定义- 形态建成的过程- 形态建成对植物生长的影响IV.光周期现象- 光周期现象的定义- 光周期现象的过程- 光周期现象对植物生长的影响V.光对植物生长的影响- 光的强度对植物生长的影响- 光的方向对植物生长的影响- 光周期对植物生长的影响VI.结论- 光对植物生长的重要性- 光对植物的影响正文:I.引言光对植物生长具有重要意义,植物通过光合作用将阳光能转化为化学能,为自身生长提供能量。
此外,植物的生长和发育过程也受到光的调控。
本文将详细探讨光对植物的影响,包括光合作用、形态建成和光周期现象等方面。
II.光合作用光合作用是植物通过吸收阳光能将水和二氧化碳转化为葡萄糖和氧气的过程。
在这个过程中,植物利用阳光能进行光激发,将电子从基态跃迁到激发态,最终传递给辅酶NADP+,形成NADPH。
NADPH 和ATP 为碳的固定提供能量,使二氧化碳转化为葡萄糖。
光合作用对植物生长具有重要意义,它为植物提供了生长所需的能量和物质。
III.形态建成形态建成是植物对环境因素(如光、温度等)的响应,通过调节细胞分裂、细胞伸长和细胞分化等过程,使植物能够适应不同的生长环境。
在光的影响下,植物会产生不同的激素,如生长素、赤霉素和细胞分裂素等,这些激素会影响植物的形态建成。
形态建成对植物生长具有重要影响,它使植物能够更好地适应环境,提高生长效率。
IV.光周期现象光周期现象是指植物对日照长度变化的反应。
不同的植物对光周期的反应不同,有的植物喜欢长日照,有的植物喜欢短日照。
植物通过光周期现象调控自身的生长和发育过程,如开花、结果等。
光周期现象对植物生长具有重要影响,它决定了植物的生长周期和生物钟。

Geometric ModelingGeometric modeling is a fundamental concept in the field of computer graphics and design. It involves creating digital representations of physical objects using mathematical equations and algorithms. Geometric modeling plays a crucial role in various applications such as animation, virtual reality, architectural design, and manufacturing. This technology allows designers and engineers to visualize and manipulate complex shapes and structures with precision and accuracy. One of the key perspectives to consider when discussing geometric modeling is its importance in the field of computer-aided design (CAD). CAD software relies heavily on geometric modeling to create 2D and 3D models of products and buildings. These models serve as the basis for the design and development of various products, ranging from consumer goods to industrial machinery. Geometric modeling enables designers to simulate the behavior of their designs under different conditions, leading to better and more efficient products. Another important perspective to explore is the role of geometric modeling in the entertainment industry. In the realm of animation and visual effects, geometric modeling is used to createlifelike characters, environments, and special effects. Whether it's a blockbuster movie or a video game, geometric modeling allows artists and animators to bring their imagination to life in a virtual space. The level of detail and realism that can be achieved through geometric modeling has revolutionized the entertainment industry and has set new standards for visual storytelling. From a scientific and engineering standpoint, geometric modeling is essential for simulating and analyzing complex systems and phenomena. Whether it's studying the behavior of fluid dynamics, analyzing the structural integrity of a building, or simulating the movement of a robotic arm, geometric modeling provides a powerful tool for understanding and predicting real-world scenarios. By creating accurate digital representations of physical objects and environments, scientists and engineers can conduct experiments and tests in a virtual space, saving time and resources while gaining valuable insights. In addition to its practical applications, geometric modeling also has a profound impact on artistic expression and creativity. Artists and designers use geometric modeling tools to explore new forms, shapes, and textures, pushing the boundaries of what is visually possible. From avant-gardesculptures to futuristic architectural designs, geometric modeling has opened up new avenues for artistic exploration and self-expression. The ability to manipulate and transform geometric shapes in a digital environment has empowered artists to create bold and innovative works that challenge traditional notions of art and design. Moreover, geometric modeling has also played a significant role in the advancement of medical technology. From the development of prosthetic limbs to the design of medical devices, geometric modeling has enabled breakthroughs in the field of healthcare. By creating precise digital models of the human body and its various systems, medical professionals and researchers can better understand and address complex medical conditions. This has led to the development of personalized medical treatments and improved patient care, ultimately saving lives and improving the quality of life for countless individuals. In conclusion, geometric modeling is a versatile and powerful tool that has revolutionized various industries and fields. Its impact can be seen in the way we design products, create entertainment experiences, conduct scientific research, express artistic vision, and improve healthcare. As technology continues to advance, the role of geometric modeling will only become more significant, shaping the way we interact with the world around us and pushing the boundaries of what is possible.。

塑料作为一种被广泛应用的有机合成聚合物材料,在为我们生活提供便利的同时,也带来了后续的环境问题。
据估算,到2050年,将有大约12000万t 塑料垃圾被埋入垃圾填埋场或自然环境中[1],塑料垃圾进入到环境后会逐渐破碎变成微塑料(<5mm ),导致其在土壤和水体中的丰度逐年递增[2-3]。
2015年的第二届联合国环境大会已将微塑料污染列为环境与生态领域的第二大科学问题[3]。
目前,人们对水体中微塑料的认识已较为系统[4-6],土壤微塑料逐步成为新的研究热点[7-8]。
2012年Rillig [9]首次提出微塑料会影响土壤理化性质,这引起了人们对土壤微塑料的关注,后续研究表明微塑料可被植物吸收并积累[10-11],最终通过食物链进入人体。
厘清微塑料对植物生长的影响及其机制,有助于系统掌握其在土壤-植物体陈欣,郭薇,李济之,等.土壤微塑料影响植物生长的因素与机制研究进展[J].农业环境科学学报,2024,43(3):488-495.CHEN X,GUO W,LI J Z,et al.Research progress on the influencing factors and mechanisms of soil microplastics on plant growth[J].Journal of Agro-Environment Science ,2024,43(3):488-495.土壤微塑料影响植物生长的因素与机制研究进展陈欣1,郭薇1,2,李济之1,2,迟光宇1*(1.中国科学院沈阳应用生态研究所,污染生态与环境工程重点实验室,沈阳110016;2.中国科学院大学,北京100049)Research progress on the influencing factors and mechanisms of soil microplastics on plant growthCHEN Xin 1,GUO Wei 1,2,LI Jizhi 1,2,CHI Guangyu 1*(1.Key Laboratory of Pollution Ecology and Environmental Engineering,Institute of Applied Ecology,Chinese Academy of Sciences,Shenyang 110016,China ;2.University of Chinese Academy of Sciences,Beijing 100049,China )Abstract :Microplastics in soil can affect plant growth in a variety of ways,accumulate in plants,and eventually enter the human body via the food chain.Clarifying the mechanisms and main factors whereby microplastics influence plant growth can contribute to a systematic understanding of their environmental behavior in soil-plant systems.Both the occurrence state and physicochemical characteristics of microplastics can influence their effects on plants.In this paper,from the perspectives of particle size,shape,concentration,and type,plastic additives,and aging degree of microplastics,we review the main factors and mechanisms underlying the effects of soil microplastics on plant growth.The key direction of future research is proposed,which will provide a reference for further clarifying the impact of microplastics on soil ecosystems.Keywords :soil;microplastics;plant;influencing factor;mechanism of action收稿日期:2023-04-13录用日期:2023-06-19作者简介:陈欣(1968—),男,辽宁沈阳人,博士,研究员,研究方向为农业生态。
Effect of tillage systems, mulches and nitrogen fertilization onspring barley (Hordeum vulgare)I. Małecka1 and A. Blecharczyk21 Poznan University of Life Sciences, Plant and Soil Cultivation Department, Mazowiecka45/46, 60-623 Poznan, Poland; e-mail: malecka@up.poznan.pl2 Poznan University of Life Sciences, Plant and Soil Cultivation Department, Mazowiecka45/46, 60-623 Poznan, Poland; e-mail: blechar@up.poznan.plAbstract. Yield, N uptake, weeds and diseases of spring barley were examined under five mulching practices (white mustard, phacelia, oat-pea mixture, straw mulch, and no mulch), three tillage systems (conventional, reduced and no-tillage) and three doses of nitrogen fertilization (0, 50 and 100 kg N ha-1). In general the grain yield of spring barley for cover crops was 10-31% higher compared with the no-mulch treatment. A mulch of straw provided a smaller barley grain yield than the no-mulch treatment. Compared to conventional tillage, grain yield under reduced tillage and no-tillage were 7 and 12% less, respectively. Spring barley sowing after a mixture of oat-pea led to decreased a negative response of reduced and no-tillage. Grain yield after treatment with legume cover crops and without N fertilization was similar compared as the rates 50 kg N ha-1 after white mustard or phacelia and as the rate 100 kg N ha-1 without mulches. There was no evidence of tillage x N fertilization interaction on grain yield, dry matter production and plant-N uptake. Cover crops and straw mulch significantly decreased total weed populations compared with the treatment without mulch. Total weed density increased from 108 plants per m2 in the no-tillage to 322 plants per m2 for reduced tillage, and to 416 plants per m2 for the conventional tillage over mulch. Higher infestation of spring barley with stem base and root diseases was observed in reduced and no-tillage in comparison with the conventional soil tillage and after straw mulch and no-mulch than after cover crops.Key words: spring barley, cover crops, tillage systems, nitrogen fertilization, grain yield, N uptake, weed and diseases infestationINTRODUCTIONCover crops and conservation tillage systems provide many benefits to crops, including erosion control, water conservation, reduced temperature fluctuations, improved soil structure, and suppression of weeds (Richards et al., 1996; Deryło, 1997; Soane & Ball, 1998; Swanton et al., 1999). Long-term benefits are derived from the buildup of organic matter resulting in increased soil health, production of healthy crops and bountiful yields. Cover crops sown after the main crop and allowed to grow through the autumn and into mind/late winter could take up nitrate-N, thus reducing the amount at risk of leaching. Therefore as much as possible of the retained N should be recovered in the next crop, enabling a reduction of fertilizer N (Richards et al., 1996; Garwood et al., 1999; Justes et al., 1999).517The effects of tillage systems on cereals grain yield have been studied previously but results are unclear. Reduced and no tillage may give more yield than conventional tillage, especially in dry years. The no tillage system led to occasionally diminished yield through decreased N availability, which suggests that cereals under reduced tillage and no tillage may require additional N fertilizer to reach production levels similar to those of conventional tillage. Potential causes of this yield depression included slow mineralization, increased N immobilization, denitrification, leaching, and surface runoff losses, but the effect of tillage systems depends heavily on the previous crop (Arshad & Gill, 1997; Canterro-Martinez et al., 2003; Angas et al., 2006; Martin-Rueda et al., 2007).Cover crops may suppress weed populations through physical suppression, competition for water, light, and nutrients, or allelopathy, however this influence is often inconsistent (Swanton et al., 1999). Research has shown that tillage systems influenced weed density and species composition (Stevenson et al., 1997). Several studies have documented that conservation tillage increased the density of perennial weeds, some annual grasses, and volunteer crops (Buhler et al., 1994; Tørresen et al., 2003). However, more research is needed on the direct effect of tillage on weed population changes over several seasons, because the weed seed shed during the first year of the experiment would remain on the soil surface and lead to a large increase in density the following year. In no-till systems, the post-harvest residues left on the field surface become a direct source of fungal diseases, especially those which infect roots (Rothrock, 1992; Bailey, 1996; Bailey & Lazarovits, 2003). The effect of tillage systems on subsequent disease severity depends heavily on the previous crop. Management of the interaction between the pathogen and the residue may be achieved through modifications in the local environment (Bailey, 1996; Bailey & Lazarovits, 2003). Alternative crops such as cover crops used to diversify cereal grain cropping systems play a valuable role in breaking the life cycle of fungal diseases (Janzen & Schaalje, 1992; Jenkyn et al., 2001; Krupinsky et al., 2002).The aim of this work was to study the effect of cover crops, tillage systems and N fertilizer rate on grain yield, N uptake, weed- and disease-infestation of spring barley.MATERIALS AND METHODSField studies were conducted from 2001–2003 at Research Station Brody of the Agricultural University of Poznan, Poland, on a sandy loam soil (Albic Luvisols, 1.4% organic matter, and pH 6.5). The experimental design was as randomized complete blocks with a three-way factorial and four replications. Each year spring barley (Hordeum vulgare var. Atol) was sowed on land that had been previously cropped to winter wheat. The sowing rate was 400 seeds per 1 m2. Treatments included five mulching practices with three tillage systems and three doses of nitrogen fertilization. The tillage systems were conventional tillage with mouldboard ploughing, reduced tillage with a shallow stubble cultivator and no–tillage. The mulches were three cover crops (Sinapis alba, Phacelia tanacetifolia and Avena sativa-Pisum sativum mixture), winter wheat straw mulch, and no mulch. Cover crops were sowed after harvest of winter wheat (Triticum aestivum ssp. vulgare) in early August. In the spring, cover crops and straw mulch were incorporated into the soil by ploughing in conventional 518tillage or by shallow stubble cultivator in a reduced tillage system. Two N treatments at rates 50 and 100 kg N ha-1 were used and included a zero N content.The fertilizer rates were applied at 26 kg P ha-1, 81 kg K ha-1 in the autumn. The herbicide program on tillage systems used preplanting and postemergence applications. In early spring before planting, 3 dm3 ha-1 glyphosate herbicide was applied on all plots with reduced tillage and direct drilling to control perennial weed and volunteers from the previous crop. For weed control during the growing season postemergence application of Stork 50 WG (tifensulfuron–methyl 25.0% and carfentrazone–ethyl 25.0%) at the rate of 60 g ha-1 was applied to spring barley. The seeds were dressed with Raxil 060 FS fungicide (60 ml per 100 kg seeds) containing tebuconazole (6.0%). For disease control Sportak Alpha 380 EC fungicide (prochloraz 30.0% and carbendazim 8.0%) at the rate 1.5 l ha-1 was applied on all plots at GS 31–32 growth stage.In the spring after crop emergence and prior to herbicide application, weed species density and aboveground fresh weights were determined in 1 m2 areas of each plot. The level of infection of spring barley with the stem base and root diseases was quantified by observing 30 randomly removed plants per each plot. The assessment of the infestation was carried out at GS 73 growth stage (Zadoks et al., 1974). Crop infestation with the stem base and root diseases was evaluated visually on the basis of the occurrence at the bottom portions of the culms and roots of characteristic spots, discolorations and necroses. For each of the diseases, the percentage of plants with symptoms (irrespective of their intensity) as well as the infestation index was calculated.The degree of infestation with the take all and the brown foot diseases was assessed according to the three-point scale (Windels & Wiersma, 1992). Grain and straw yields were estimated from a harvested area of 15 m2 on each plot using a plot combine harvester. A subsample of grain and straw was taken from each plot to determine moisture, grain weight and N content. Nitrogen in the grain and straw was determined by near infra-red spectrophotometry using InfraAlyzer 500 (Bran+Luebbe) that was calibrated with the standard Kjeldahl method. Number of ears was counted from 0.5 m2 on each plot before harvest. After recording a thousand grain weight the number of grain per ear was calculated.The harvest index was calculated as the ratio of grain dry matter yield to the total dry matter yield (grain+straw). N uptake was estimated by the product of dry matter of grain or straw and N concentration of grain or straw. The nitrogen harvest index was calculated as the ratio of grain N accumulation to the total plant N accumulation (grain+straw).Standard analysis of variance (ANOVA) was performed to determine the main factors and the interactions. Mean separations were made for significant effects with LSD and Tukey tests at probability P≤ 0.05.The weather conditions varied markedly during the experimental period (Table 1).519Table 1. Weather characteristics.Average monthly temperatures (°C)YearMarch April May June July mean2001 2002 2003 1961–1999 2.65.13.42.78.18.88.27.514.816.716.012.815.318.319.816.220.320.419.617.712.213.913.411.4Monthly precipitation (mm)YearMarch April May June July sum2001 2002 2003 1961–1999 70.858.119.936.137.333.221.139.134.748.920.156.675.652.635.067.153.440.696.778.3271.8233.4192.8277.2During the vegetation period of spring barley, the mean air temperature was above the long-term mean, with the exception of June 2001. Growing season precipitation from 2001–2003 (from sowing to harvest) was 201, 175 and 173 mm, respectively. Greater variations of precipitation occurred during the period April–June. In this period the higher precipitation was in the years 2001 (148 mm) and 2002 (135 mm) in comparison with the last year (76 mm). In conclusion, weather conditions were more favorable to the development of spring barley in 2001 and 2002 than in 2003.RESULTS AND DISCUSSIONGrain yield and yield componentsSpring barley grain yields were significantly affected by mulches, tillage systems and N fertilization (Table 2).The results of mulch establishment varied among mulch types. Over all years the oat-pea mixture gave the highest recorded grain yield (4.94 t ha-1 mean of years). Grain yield for cover crops was 10-31% higher compared with no-mulch treatment. Mulch of straw provided lower barley grain yield than no-mulch treatment, especially in the 2002 season (14.5%). The yield of spring barley varied significantly with tillage systems, both for the individual years and for the 3-year period as a whole. Compared to conventional tillage, grain yield under reduced tillage and no-tillage were 7 and 12% less, respectively. The analysis of variance showed interaction affects mulches x tillage for grain yield of spring barley (Table 3). The effect of the tillage systems depended upon the mulches. Spring barley sowing after the mixture of oat-pea led to a decreased negative response of reduced and no-tillage.There were no significant differences between tillage systems on the plots with a cover crop of oat-pea mixture. A negative response for no-tillage compared to reduced tillage was observed in no-mulch treatments only. Most studies on cereal production comparing conventional and ploughless tillage have given controversial results, apparently depending on soil type, crop rotation and local climatic conditions (Nyborg et al., 1995; Arshad & Gill, 1997; Halvorson et al., 2000; Małecka & Blecharczyk, 2002; Cantero-Martinez et al., 2003; Angas et al., 2006; Martin-Rueda et al., 2007).520Table 2. Grain yields of spring barley for mulches, tillage systems and nitrogen fertilization, t ha-1.YearTreatments2001 2002 2003 2001–2003NoneStrawWhite mustard Oats+pea Phacelia 4.324.164.755.644.633.462.964.104.694.093.433.353.954.503.873.773.494.274.944.16LSD0.05 0.30 0.51 0.43 0.23Conventional ReducedNo-till 5.164.624.323.993.843.754.063.833.574.404.103.88LSD0.05 0.280.140.190.21 N rate, kg ha-1:0 50 100 3.824.795.502.873.944.763.303.884.273.334.204.84LSD0.05 0.220.230.240.21Table 3. Effect of mulches and tillage systems on grain yield of spring barley, t ha-1 (mean 2001–2003).Tillage systemsMulchconventional reduced no-tillStrawWhite mustard Oats+pea Phacelia 3.844.545.054.473.454.244.894.093.174.034.883.92LSD0.050.29 Table 4. Effect of mulches and nitrogen fertilization on grain yield of spring barley, t ha-1(mean 2001–2003).Nitrogen rate (kg ha-1)Mulch0 50 100NoneStrawWhite mustard Oats+pea Phacelia 2.872.683.384.383.323.733.564.425.084.224.704.225.005.364.94LSD0.05 0.33 Reduced mineralization of plant residues in addition to mechanical difficulties affecting seed placement have often been considered responsible for the low yields obtained with reduced and no-tillage systems (Lopez-Fando & Almendros, 1995).Average grain yield increased significantly with increasing N rates (Table 2). The greatest fertilizer rate (100 kg ha-1) tended to increase grain yield 15% as comparedwith rate 50 kg ha-1. No significant difference was observed between the rates 50 and521100 kg N ha-1 on the plots with mixture of oat-pea (Table 4). Grain yield after treatment with legume cover crops and without N fertilization was similar as compared to the rate of 50 kg N ha-1 after white mustard or phacelia and to the rate 100 kg N ha-1 without mulches.Legume cover crops offer an effective method of incorporating atmospheric N into the soil and improving nitrogen fertility (Janzen & Schaalje, 1992; Richards et al., 1996; Garwood et al., 1999). It provides a way to address significant management challenges to producers interested in reducing fertilizer N inputs or producing crops without fertilizer N. The positive effect of other cover crops on barley grain yield decreased with an increased N fertilizer rate. With straw mulch treatment the average grain yield response to N applications was less than in the other treatments which removed straw. These results were generally similar to earlier results (Nyborg et al., 1995; Procházková et al., 2002). The problems emerged from a probably increased N immobilization caused by straw addition during the first years of the experiment, but disappeared thereafter, and from a possible inhibitory effect on germination, emergence, growth and yields of following crops. The transition from intensive to conservative tillage systems will require the modification of N dynamics in the soil-plant system, on crop productivity, and on efficiency of N fertilizer use. During the first few years of a change in tillage systems, farmers tend to increase N rates to secure high yields (Wienhold & Halvorson, 1999). However, in some cases this increase is not necessary, because the availability of N improves in a few years (Cantero-Martinez et al., 2003; Angas et al., 2006). This was in agreement with our results in which no significant interaction between N rates and tillage systems was observed.In the present paper, yield components such as ears per 1 m2, grains per ear and grain weight were also quantified (Table 5).Table 5. Yield components of spring barley for mulches, tillage systems and nitrogen fertilization (mean 2001–2003).Treatment Ears per 1 m2 Grains per ear(g)Mulch:NoneStrawWhite mustard Oats+pea Phacelia 50446755361854716.716.817.217.516.745.745.245.646.346.3LSD0.05 36.50.390.58 Tillage systems:Conventional ReducedNo-till 59052849517.116.917.044.346.446.7LSD0.05 18.5ns0.48 N rate, kg ha-1:0 50 100 45454461615.917.117.946.146.145.2LSD0.05 14.80.260.38522Lower yields resulted first of all from fewer ears per m2. The mean of years with higher ears per m2 were on the plots with cover crops rather than with straw mulch and none. The effect of mulches on grains per ear and grain weight varied by year. Number of ears per m2 and grain weight were significantly influenced by the main effect of tillage systems. The number of ears was higher under conventional tillage than under other tillage systems, however, the opposite result was observed for grain weight. The number of ears per m2 and grains per ear increased with increasing N rate both for the individual years and for the studied period as a whole. The effect of N fertilizer on grain weight differed by individual year. In 2001 grain weight decreased with increasing N rate; in 2002 grain weight was maximum with the medium N rate. In 2003 grain weight increased with increasing N rate, with no difference between the rates 50 and 100 kg N ha-1. For the 3-years as a whole, the greatest fertilizer rate (100 kg N ha-1) had an adverse effect on grain weight. Our results are comparable with results of other research (Campbell et al., 1993; Cantero-Martinez et al., 2003).Total dry matter and nitrogen accumulation at maturityTotal dry matter production and N accumulation at maturity were affected more by N fertilization and mulches than by tillage systems (Table 6).Table 6. Mean total aboveground dry matter, N accumulation and harvest index of spring barley at maturity as affected by mulches, tillage systems and nitrogen fertilization (mean 2001–2003).TreatmentsTotaldrymatter(t ha-1)Graindrymatter(t ha-1)HarvestindexTotal Naccumulation(kg ha-1)Grain Naccumulation(kg ha-1)NitrogenharvestindexMulch:NoneStrawWhite mustard Oats+pea Phacelia 5.965.496.937.946.683.202.973.634.203.540.540.540.520.530.5374.169.784.1103.985.556.453.262.778.863.70.760.760.750.760.75LSD0.05 0.490.20ns 4.3 3.1 ns Tillage systems:Conventional ReducedNo-till 7.066.566.183.743.493.300.530.530.5388.982.579.066.363.059.90.710.760.76LSD0.05 0.310.18ns 3.1 3.20.04 N rate, kg ha-1:0 50 100 5.206.737.862.833.574.110.540.530.5261.483.3105.746.462.580.10.760.750.76LSD0.05 0.290.17ns 2.9 2.8 ns ns–no significanceTotalabove-ground biomass for mixture of oat-pea, white mustard and phacelia were 33, 16 and 12% higher compared with the no-mulch treatment. Reduced tillage523and no-tillage resulted in less biomass than conventional tillage (7–12%). The highest N accumulation in total dry matter followed the mixture of oat-pea (103.9 kg ha-1). On the other treatments N accumulation ranged between 69.7-85.5 kg ha-1 (33–18% less). Conventional tillage achieved slightly but significantly greater total N accumulation than ploughless treatments. Generally, total dry matter and N accumulation is favoured in tilled plots in wet years and in no-tillage during dry ones (Arshad & Gill, 1997; Cantero-Martinez et. al., 2003; Tørresen et al., 2003; Martin-Rueda et al., 2007). Nitrogen application increased above-ground dry matter and N accumulation in the whole plant. The greater N accumulation was associated with total dry matter rather than with protein concentration (data not shown). The positive relationship between rate of fertilizer N and total dry matter and plant N accumulation at maturity is consistent with the results of others (Bulman & Smith, 1993; Nyborg et al., 1995; Cantero-Martinez et al., 2003). In our investigation there was no evidence of tillage x N fertilization interaction on dry matter production and plant-N uptake. Other authors described different types of interactions between tillage systems and N fertilization on dry matter production. This type of response is produced when some factor other than nitrogen is the greater determinant of yield (Nyborg et al., 1995; Halvorson et al., 2000; Cantero-Martinez et al., 2003; Angas et al., 2006). Harvest index and nitrogen harvest index were not affected by mulches or N fertilization. Tillage systems had significant effect only on nitrogen harvest index. It was smaller for conventional tillage than for the reduced and no-tillage system.Cover crops may also have provided some of the non-N benefits to the subsequent crops: reduced incidence of root and leaf diseases, reduced weed populations, increased nutrient availability (Janzen & Schaalje, 1992; Arshad & Gill, 1997).WeedsThe effects of mulches, tillage systems and nitrogen fertilization on weeds are shown in Table 7.Cover crops and straw mulch significantly decreased total weed populations compared with the treatment without mulch (mean by 20%). Previous research reported that weed suppression usually increases as amount of crop residues increases (Teasdale et al., 1991).Total weed density increased from 108 plants per m2 in the no-tillage to 322 plants per m2 for reduced tillage, and to 416 plants per m2 for the conventional tillage over mulch. Some studies have indicated that reduced tillage can increase population of weeds (Buhler et al., 1994; Tørresen et al., 2003); in others, weed communities were influenced more by location, crop, and year than by tillage system (Teasdale et al., 1991; Swanton et al., 1999). The treatments had lower effects on weed biomass than on weed density. Compared with no-mulch treatment, cover crops reduced mean weed biomass by 8%. There was no difference between straw mulch and no-mulch treatment in weed biomass. Fresh weight of weeds increased from 120 g m-2 in no-tillage to 204 g m-2 for the reduced tillage and 207 g m-2 for conventional tillage. This is in agreement with previous research which showed higher weed biomass after tillage with a uniform soil seed density (Teasdale et al., 1991).524Table 7. Effect of mulches, tillage systems and nitrogen fertilization on density and fresh weight of weeds (mean 2001–2003).Treatments(No. m-2) (g m-2)NoneStrawWhite mustard Oats+pea Phacelia 337253269269282184194176154177LSD0.05 35.313.6 Tillage systems:Conventional ReducedNo-till 416322108207204120LSD0.05 20.811.5-10 50 100 269280297105199226LSD0.05 10.39.2The dominant weed species across all treatments were Viola arvensis, Veronicahederifolia, Chenopodium album, Polygonum convolvulus,Stelaria media, and Thlaspiarvense.Viola arvensis and Chenopodium album density increased in conventionaltillage compared with reduced and no-tillage systems. Species that increased in densityin no-tillage system were Taraxacum officinale, Plantago major, Geranium pusillum,Poa annua, and Brassica napus. Several studies have documented that conservationtillage increased the density of perennial weeds, some annual grasses, and volunteercrops but a significant problem arose after several years of research (Swanton et al.,1999). Total weed populations and increase in fresh biomass of weeds were directlyrelated to fertilizer N rates.DiseasesThe occurrence of stem base and root diseases in spring barley depended onexperimental factors (Table 8).The dominant pathogens identified in our experiment were Gaeumannomycesgraminis (Sacc.) v. Arx et Olivier causing the take-all disease, and Fusarium spp.causing brown foot rot. On average, the results of three-year investigations showed asignificantly higher infestation of spring barley with stem base and root diseases afterstraw mulch and no-mulch than after cover crops. It was observed in the percentage ofculms with symptoms of the take-all and the brown foot rot and the infestation index.Results of our investigations confirm the above-mentioned reports of other researchers.The use of a break crop in cereal rotation plays a valuable role in breaking the lifecycle of fungal diseases in the soil (Bailey, 1996; Krupinsky et al., 2002; Bailey &Lazarovits, 2003).525Table 8. The infection of spring barley by stem base and root diseases depending on mulches, tillage systems and nitrogen fertilization (mean 2001–2003).Gaeumannomyces graminis Fusarium spp.Treatmentsinfectionstems infectionindex% of infectionstemsinfectionindexMulch:NoneStrawWhite mustard Oats+pea Phacelia 43.456.231.828.730.612615841354373.673.749.749.049.8157159818189LSD0.05 15.831.219.430.9 Tillage systems:Conventional ReducedNo-till 31.938.544.0429910150.464.063.381127132LSD0.05 5.210.65.615.1 N rate, kg ha-1:0 50 100 42.237.834.4122625866.456.254.914410888LSD0.05 4.215.63.518.2The results of the Brody investigation indicated a negative influence of reducedtillage and no-tillage on the infestation of spring barley with stem base and rootdiseases in comparison with the conventional soil tillage. Reduced tillage and no-tillage leaves more crop residues on the soil surface, and this in turn may cause highersoil moisture content and lower temperature favouring the occurrence of stem base androot diseases (Rothrock, 1992; Bailey, 1996). On the other hand, the plant residues lefton the soil surface result in increased organic matter in the topsoil and hence increasedbiological activity which can reduce pathogen populations in the soil (Krupinsky et al.,2002). That is why research results concerned with the impact of soil tillage systems,especially no-tillage, on the occurrence of fungal diseases are quite inconsistent.According to many researchers, the impact of tillage systems on fungal diseases ismuch smaller than the effect exerted by environmental conditions (weather, soil), cropsequence, cultivars and type of seed dressing (Rothrock, 1992; Bailey, 1996). Both theinfestation index and the percentage of plants with symptoms of take-all and brownfoot rot disease depended on nitrogen fertilization. The stem base and root diseaseseverity was significantly higher in the plots where spring barley was grown without Nfertilization in comparison with fertilized plots. Balanced and adequate fertility for anycrop reduces plant stress, improves physiological resistance, and decreases disease risk(Krupinsky et al., 2002).526。
Unit1 Text AText-related exercisesI.1. Agronomy is the science and technology of producing and using plants for food, fuel, fiber, and reclamation2. They are involved with any issues including producing food, creating healthier food, managing environmentalimpact of agriculture, and creating energy from plants.3. To produce the best crops under various conditions.4. By change the genetics of the plants.5. In labs6. Compounds of nitrogen, phosphorus, and potassium.7. Contour plowing.8. Sustainable agriculture, organic farming, alternative food systems and the development of alternative croppingsystems.9. Quantitively.10. Four to twelve years.II.1. 作物育种2. 轮作,轮种3. 植物生理学4. 有机耕作5. 杂草控制,除草6. plant genetics7. selective breeding8. contour plowing9. soil fertility 10. (insect and)pest control III.1. plants2. combines3. healthier4. agriculture5. genetics6. yields7. nutritional8. civilization9. security 10. resistantIV.1. Agronomic/Agricultural biotechnology is increasingly being used applied for novel/new uses other than food.2. Agronomy involves with biology, ecology, chemistry and other sciences.3. Selective breeding of plants is to produce/develop/breed the best crops adapted to different environments andgrowing conditions.4. Agronomists often specialize in areas such as crop rotation, soil classification, weed control and insect andpest control.5. Agroecology studies the management of agricultural system on ecological and environmental perspetives. Theme-related ExercisesI.1-6 F T F T F FII.1. M potential2. K adoption3. I acute4. H combine5. F work6. G actively7. D optimally8. C initiative9. N available 10. A promoteText BText-related exercisesI.1. Ki ll pests’ natural enemies and their overuse can harm farmers, consumers and the environment.2. When the natural processes of pest regulation break down.3. Ecosystem approach4. To manage insect pest populations to the point where natural predation operates in a balanced way and croplosses to pests are kept to an acceptable minimum5. More than/exceed US $40 billion6. Almost the same/ little change. 30-40 percent.7. Less than 1%. The rest contaminates/pollutes the air, soil and water.8. Urban and peri-urban horticulture9. Because the pesticides are aggressively marketed.10. In 1960s. IPM has been based on ecology, the concept of ecosystems and the goal of sustaining ecosystemfunctions.II.a -- 3,b -- 6,c -- 1,d -- 2,e -- 7,f -- 4,g -- 5III.1. option2. contaminates3. negative4. vicious cycle5. Pesticide6. excessive7. Health8. Occupational9. pesticide residues 10. ecosystemsIV.1.The risks posed by pesticides to health and the environment have aroused public concernsworldwide/globally.2. Before the Industrial Revolution farming was a labor-intensive industry, and the vast majority of the humanpopulation labored in agriculture3. Usually proper use of pesticides can, to some extent, keep pests in check.4. Over-reliance on pesticides contributes to a vicious cycle of resistance in pests.5. Excessive use of pesticide exposes farmers to serious health risks and has negative consequencesfor/effects/influences on/ the environment.Theme-related ExercisesI.1-5 A C D B CII.作为基于生态系统的战略,病虫害综合治理在世界农业发展中已取得了一些显著的成绩。
农作物管理英语English:Crop management involves various practices aimed at optimizing the growth, health, and yield of crops. It encompasses a range of activities including soil preparation, planting, irrigation, fertilization, pest and weed control, and harvesting. Soil preparation is crucial as it ensures the right conditions for seed germination and root development. Planting involves selecting appropriate seeds or seedlings and placing them in the soil at the correct depth and spacing. Irrigation ensures crops receive adequate water, crucial for photosynthesis, nutrient uptake, and overall plant health. Fertilization involves supplying essential nutrients to the soil or directly to plants to support growth and development. Pest and weed control are essential to prevent damage to crops and competition for resources. Harvesting involves timing the collection of mature crops to ensure maximum yield and quality. Effective crop management requires knowledge of crop biology, environmental conditions, and careful planning to optimize resources and minimize environmental impact.中文翻译:农作物管理涉及各种旨在优化农作物生长、健康和产量的实践。
农村除草英语作文Rural WeedingWeeding is a crucial agricultural practice that plays a vital role in maintaining the health and productivity of crops in rural areas. It involves the removal of unwanted plants, commonly known as weeds, from cultivated land. Weeds can compete with crops for essential resources such as water, nutrients, and sunlight, thereby reducing crop yield and quality. Furthermore, weeds can harbor pests and diseases that can further threaten the well-being of the crops. Consequently, effective weed management is a fundamental aspect of successful farming in rural communities.In rural settings, weeding is often a labor-intensive task that is carried out manually by farmers and their families. This traditional approach to weed control has been practiced for generations and remains an essential component of many traditional farming systems. Manual weeding requires the physical removal of weeds by hand or with the use of simple tools such as hoes, sickles, or machetes. This method is particularly effective for small-scale farms and gardenswhere the terrain and crop layout may not be suitable for mechanical or chemical weed control methods.One of the primary advantages of manual weeding in rural areas is its low cost and accessibility. Farmers in these regions often have limited access to advanced agricultural technologies or financial resources, making manual weeding a more viable and sustainable option. Additionally, manual weeding can be a valuable source of employment and income for rural communities, providing opportunities for farmers and their families to contribute to the overall productivity of the land.Another benefit of manual weeding is its environmental sustainability. Unlike chemical-based weed control methods, manual weeding does not introduce harmful substances into the soil or surrounding ecosystems. This is particularly important in rural areas, where the preservation of natural resources and the maintenance of healthy ecosystems are crucial for the long-term viability of agricultural production.However, manual weeding is not without its challenges. It can be physically demanding and time-consuming, especially during peak growing seasons when weeds tend to proliferate rapidly. This can place a significant burden on rural farmers, who often have limited labor resources and must balance weed management with otheressential agricultural tasks.To address these challenges, some rural communities have developed innovative strategies to enhance the efficiency and effectiveness of manual weeding. For instance, the use of specialized tools, such as hand-held weeders or wheel hoes, can help to reduce the physical strain on farmers and increase the speed of weed removal. Additionally, the implementation of crop rotation and cover cropping practices can help to suppress weed growth and reduce the overall need for weeding.Another approach to improving manual weed control in rural areas is the incorporation of traditional knowledge and practices. Many rural communities have accumulated a wealth of knowledge about the local ecosystem, including the identification and management of various weed species. By drawing on this traditional knowledge, farmers can develop more targeted and effective weeding strategies that are tailored to the unique characteristics of their local environment.Furthermore, the integration of manual weeding with other weed control methods, such as the use of mulch or the strategic timing of planting and harvesting, can help to create a more comprehensive and sustainable weed management system. By combining different techniques, rural farmers can optimize the use of available resourcesand minimize the impact of weeds on their crops.In recent years, there has also been a growing interest in the development of low-cost, environmentally friendly weed control technologies that are suitable for rural settings. These include the use of cover crops, mechanical weeders, and even the incorporation of biological control agents, such as beneficial insects or pathogens, to manage weed populations.Despite the challenges, manual weeding remains a vital practice in many rural agricultural communities. It not only contributes to the overall productivity and sustainability of farming but also serves as an important cultural and social activity that brings communities together. By embracing traditional knowledge, leveraging innovative technologies, and fostering collaborative approaches, rural farmers can continue to refine and improve their weed management practices, ensuring the long-term viability and resilience of their agricultural systems.。