细胞信号转导
- 格式:docx
- 大小:20.75 KB
- 文档页数:5


第十九章细胞信号转导第十九章细胞信号转导一、内容提要细胞信号转导是指特定的化学信号在靶细胞内的传递过程,主要由信号分子的识别与接受,信号在细胞内的放大与传递,以及特定生物学效应的产生三个过程组成。
信号分子是指由特定的信号源(细胞)产生的,可以通过扩散或体液转运等方式进行传递,作用于靶细胞并产生特异应答的一类化学物质,包括激素、神经递质、细胞因子、生长因子及无机物等几大类。
由信号细胞释放的信号分子,需经扩散或转运,才能够到达靶细胞产生作用。
根据传递距离的远近,可将信号分子的传递分为内分泌、旁分泌和自分泌信号传递三种方式。
受体是指存在于靶细胞膜上或细胞内的一类特殊蛋白质分子,它们能够识别与结合化学信号分子,并触发靶细胞产生特异的生物学效应。
按照受体存在的亚细胞部位的不同,可将其分为细胞膜受体和细胞内受体二大类,前者又分为跨膜离子通道受体、G蛋白偶联受体和单跨膜受体。
受体的作用特点包括高度的亲和力、高度的特异性、可逆性、可饱和性及特定的作用模式等。
由细胞内若干信号转导分子所构成的级联反应系统就被称为细胞信号转导途径,目前已经鉴定的细胞信号转导途径达10多条。
大多数的激素、神经递质、生长因子和细胞因子通过膜受体介导的信号转导途径传递信号,这些信号转导途径的共同特征都是通过一系列的级联反应,以激活特定的蛋白激酶并对其底物蛋白或酶进行共价修饰,从而产生特定的生物学效应。
在这些信号转导途径中,以环核苷酸(cAMP和cGMP)作为第二信使的信号转导途径是目前较为清楚的信号转导途径。
除此之外,以脂类衍生物,如IP3、DAG、PI-3,4-P2、PI-3,4,5-P3等作为第二信使的信号转导途径,以及以钙离子作为第二信使的Ca2+信号转导途径也越来越受到重视。
而胰岛素、生长因子及细胞因子则主要通过酪氨酸蛋白激酶(TPK)信号转导途径传递信号。
亲脂性的激素主要通过胞内受体介导的信号转导途径传递信号,这一途径通过活化受体调控特异基因的转录表达来产生特定的生物学效应。

细胞信号转导细胞信号转导(cell signal transduction):指的是偶联各种胞外刺激信号与其相应的生理反应之间的一系列分子反应机制。
其分子途径分为三个阶段:1、胞外刺激信号传递(1)环境刺激:(光、温度、水分、重力、伤害、病原菌毒物、矿物质及气体)最重要的环境刺激是光,光是光合作用的能源,光强、光质可作为信号激发受体,引起光形态建成。
(2)胞间信号传递:当环境刺激的作用位点与效应位点处在不同部位时,就必然发生信号的产生和传递。
这些胞间信号(化学信号和物理信号)及某些环境刺激信号就是细胞信号转导过程中的初级信号,即第一信使(first messenger)。
A、化学信号(chemical signals):指细胞感受环境刺激后形成,并能传递信息引起细胞反应的化学物质,如:植物激素(ABA、GA、IAA等)、植物生长活性物质。
胞间化学信号长距离传递的主要途径是韧皮部,并且可以同时向顶和向基传递,传递速度为0.1-1 mm·s-1;其次是木质部集流传递。
B、物理信号(physical signals):指细胞感受环境刺激后产生的具有传递信息功能的物理因子,如:电波、水力学信号等。
胞间物理信号电波长距离传递途径是维管束,短距离传递则通过共质体及质外体。
敏感植物动作电波的传播速度可达200 mm·s-1 。
2、膜上信号转换(1)受体(receptor):受体:指位于细胞质膜上能与化学信号物质特异地结合,并能将胞外信号转换为胞内信号,发生相应细胞反应的物质。
质膜表面有三种类型受体:a、G蛋白偶联受体(G-protein-linked receptor)b、酶联受体(enzyme -linked receptor)c、离子通道偶联受体(ion-channel-linked receptor)受体与化学信号物质的识别反应是细胞信号转导过程中的第一步。
(2)G蛋白G蛋白:GTP结合调节蛋白(GTP binding regulatory protein ),膜上信号转换是通过G蛋白偶联的。

细胞信号转导细胞信号转导是指细胞内外信息的传递和转化过程,这一过程起着调节和控制细胞生理活动的重要作用。
通过信号传递,细胞可以对外界环境做出适应性的反应,维持内部稳态,实现生长、分化和细胞命运决定等功能。
本文将从信号的产生、传递和转导机制等方面进行讨论。
一、信号的产生1. 内源性信号细胞自身产生的化学物质可以作为信号分子,以调节细胞内外环境。
例如,细胞内的离子浓度、pH值和代谢产物等,都可以通过信号传递机制发挥作用。
2. 外源性信号外界环境中的物质和刺激也可以作为细胞信号的来源。
例如,细胞表面的受体可以与激素、细菌毒素和细胞外基质等结合,引发相应的信号传递。
二、信号的传递细胞信号传递通常有三种主要方式:通过直接细胞接触、通过细胞间联系以及通过远距离的物质传递。
1. 直接细胞接触细胞表面的受体与邻近细胞的配体结合,通过接触传递信号。
这种方式在免疫系统的活化、神经细胞的传递和胚胎发育等过程中起重要作用。
2. 细胞间联系细胞通过细胞间连接物质(如细胞间隙连接、紧密连接和连接蛋白)进行信号传递。
这种方式在组织内细胞间的协调和相互影响中起到重要作用。
3. 物质传递一些信号分子可以通过远距离的物质传递,例如激素、细菌毒素和神经递质等。
它们通过血液、淋巴液和突触间隙等途径到达目的地细胞,触发相应的信号级联反应。
三、信号的转导机制1. 受体的激活和信号传导当信号分子结合至受体上时,受体会发生构象变化,从而激活相应的信号通路。
这种激活过程包括泛素化修饰、磷酸化等,促使信号传导的启动。
2. 信号通路的级联反应一旦信号通路被启动,连锁反应会引发一系列级联反应。
这些反应会通过激活一些键酶、转录因子和细胞器等,最终产生细胞内外多种生理活动的结果。
3. 信号的转导和传递信号通路中的组分和中介物质可以通过蛋白质相互作用、分子承载体和次级信号等方式,进行信号的转导和传递。
这种方式可以将信号的强度和特异性传递至下游组分,以发挥预期的生物学功能。

一、细胞信号转导概述(一)信号转导的概念在多细胞生物体中,细胞间的信号转导(signaltransduction)与交换对细胞的生存非常重要。
细胞的信号转导是通过多种分子相互作用的一系列有序反应,将来自细胞外的信息传递到细胞内各种效应分子,并产生生物效应的过程。
通常所指的信号转导是指跨膜信号转导(transmembrane signal transduction),即生物活性物质(如神经递质、激素、细胞因子等)通过受体或离子通道的作用,将其转变为细胞内各种分子数量、分布或活性的变化,从而对细胞的功能、代谢、生长速度、迁移等生物学行为产生影响。
(二)信号转导系统的基本组成细胞信号转导系统通常由信息分子(signaling molecule)、受体(receptor)、转导体(transducer)及效应体(effector)四个环节组成。
信息分子的受体位于靶细胞的质膜上、胞质或核内,与之相结合的相应信息分子统称为配体(ligand)。
配体与受体的结合可诱导受体的构象发生变化,激活转运体,进而启动细胞内的信息转导途径(如效应体的级联反应),最终导致细胞功能的改变。
(三)信号转导的主要途径根据介导的配体和受体的不同,信号转导可分为两大类,一类是水溶性配体或物理信号作用于膜受体,随后经历跨膜和细胞内信号转导体的依次作用,最终作用于效应体,产生效应。
依据膜受体特性的不同,这类信号转导又有多种通路,主要是由离子通道型受体、G蛋白耦联受体、酶联受体和招募型受体介导的信号转导。
另一类是脂溶性配体直接与胞质受体或核受体结合而发挥作用,这类方式通常都是通过影响基因表达而产生效应。
应当注意到膜受体介导的信号转导也大多可以影响转录因子的活性而改变基因的表达。
(四)信号转导途径间的交互联系细胞信号转导通路的细节非常复杂,涉及蛋白质等相互作用以及相关基因表达的过程,而且各种信号转导通路间存在更为复杂的联系,构成错综复杂的信号网络(signaling network)。

细胞信号转导细胞信号转导是细胞内外环境信息传递和响应的过程。
在细胞内外环境发生变化时,细胞通过感知这些信号并传导到细胞内部,最终引发一系列的生物学效应。
本文将介绍细胞信号转导的基本概念、机制与重要研究领域。
一、信号转导的基本概念细胞信号转导是细胞内外信号信息通过具体的分子机制传递到细胞内部,并且在细胞内引发相应的生物学反应。
信号可以是化学物质、光线、温度和压力等,这些信号通过细胞膜表面受体或胞浆内受体与信号分子特异性结合,从而激活一系列的信号转导分子。
细胞信号转导的过程通常包括受体激活、信号传导、增强或抑制等多个环节。
二、信号转导的机制在细胞信号转导的过程中,不同信号可以通过不同的机制进行转导,包括直接通过受体激活、信号级联放大、二级信号传导以及负反馈调控等机制。
1. 直接激活:有些信号可以直接通过受体激活下游分子,例如膜受体激活酪氨酸激酶,进而磷酸化下游调节因子。
2. 信号级联放大:部分信号转导可以通过级联放大的方式增强信号的强度和传递效果。
一个典型例子是G蛋白偶联受体信号转导通路,一个G蛋白偶联受体可以激活多个G蛋白,每个G蛋白可进一步激活下游信号转导分子。
3. 二级信号传导:某些信号分子可以通过激活下游信号分子形成二级信号传导,例如细胞内钙离子浓度的增加可以激活蛋白激酶C,进而磷酸化下游的蛋白质。
4. 负反馈调控:为了避免过度的信号激活,细胞常常会通过负反馈调控机制来抑制信号转导分子的活性,以保持信号的动态平衡。
三、细胞信号转导的重要研究领域细胞信号转导是生物学的重要研究领域,许多科学家致力于探索细胞内信号传导的机制和调控网络。
以下是其中的几个重要研究领域:1. 肿瘤信号转导:细胞信号转导的异常调控与肿瘤的发生和发展密切相关。
研究人员通过研究与肿瘤发生相关的信号转导通路,探索肿瘤的分子机制,并寻找新的治疗靶点。
2. 免疫信号转导:细胞信号转导在免疫系统中起着重要的作用。
研究人员致力于解析免疫应答的信号转导网络,以揭示免疫反应的机制,为免疫相关疾病的治疗提供新的思路。



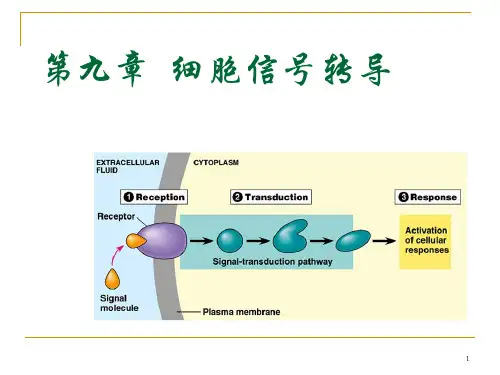

细胞信号转导综述09级临床2班隋德岭0941105217一、细胞信号转导的概念细胞信号转导是指细胞通过胞膜或胞内受体感受信息分子的刺激,经细胞内信号转导系统转换,从而影响细胞生物学功能的过程。
水溶性信息分子及前列腺素类(脂溶性)必须首先与胞膜受体结合,启动细胞内信号转导的级联反应,将细胞外的信号跨膜转导至胞内;脂溶性信息分子可进入胞内,与胞浆或核内受体结合,通过改变靶基因的转录活性,诱发细胞特定的应答反应。
传导通路示意图二、信号转导受体[1](一)膜受体1.环状受体(离子通道型受体)多为神经递质受体,受体分子构成离子通道。
受体与信号分子结合后变构,导致通道开放或关闭。
引起迅速短暂的效应。
2.蛇型受体7个跨膜α-螺旋受体, 有100多种,都是单条多肽链糖蛋白,如G蛋白偶联型受体[2] [3]。
G蛋白示意图3.单跨膜α-螺旋受体包括酪氨酸蛋白激酶型受体和非酪氨酸蛋白激酶型受体。
(1)酪氨酸蛋白激酶型受体这类受体包括生长因子受体、胰岛素受体等。
与相应配体结合后,受体二聚化或多聚化,表现酪氨酸蛋白激酶活性,催化受体自身和底物Tyr磷酸化,有催化型受体之称。
(2)非酪氨酸蛋白激酶型受体,如生长激素受体、干扰素受体等,。
当受体与配体结合后,可偶联并激活下游不同的非受体型TPK,传递调节信号。
(二)胞内受体位于胞液或胞核,结合信号分子后,受体表现为反式作用因子,可结合DNA顺式作用元件,活化基因转录及表达。
包括类固醇激素受体、甲状腺激素受体等。
胞内受体都是单链蛋白,有4个结构区:①高度可变区②DNA结合区③激素结合区④绞链区(三)受体与配体作用的特点是:①高度亲和力,②高度特异性,③可饱和性1.受体:位于细胞膜上或细胞内,能特异性识别生物活性分子并与之结合,进而引起生物学效应的特殊蛋白质,膜受体多为镶嵌糖蛋白:胞内受体全部为DNA 结合蛋白。
受体在细胞信息传递过程中起极为重要的作用。
2.G蛋白:即鸟苷酸结合蛋白,是一类位于细胞膜胞浆面、能与GDP或GTP 结合的外周蛋白,由α、β、γ三个亚基组成。
细胞的信号转导信号转导(signal transduction):指在信号传递中,细胞将细胞外的信号分子携带的信息转变为细胞内信号的过程完整的信号传递程序:1、合成信号分子;2、细胞释放信号分子;3、信号分子向靶细胞转运;4、信号分子与特异受体结合;5、转化为细胞内的信号,以完成其生理作用;6、终止信号分子的作用;第一节、细胞外信号1、由细胞分泌的、能够调节机体功能的一大类生物活性物质。
如:配体2、配体的概念:指细胞外的信号分子,或凡能与受体结合并产生效应的物质。
3、配体的类型:1)水溶性配体:N递质、生长因子、肽类激素2)脂溶性配体:甲状腺素、性激素、肾上腺激素4、第一信使:指配体,即细胞外来的信号分子。
第二节、受体一、受体的概念:细胞膜上或细胞内一类特殊的蛋白质,能选择性地和细胞外环境中特定的活性物质结合,从而引起细胞内的一系列效应。
二、受体的类型:细胞表面受体胞内受体(胞浆和核内)1、细胞表面受体类型1)离子通道偶联受体:特点:本身既有信号结合位点又是离子通道组成:几个亚单位组成的多聚体,亚单位上配体的结合部位,中间围成离子通道,通道的“开”关受细胞外配体的调节。
2)酶偶联受体:或称催化受体、生长因子类受体,既是受体,又是“酶”。
特点:N端细胞外区有配体结合部,C端细胞质区含特异酪氨酸蛋白激酶(TPK)的活性。
组成:一条肽链一次跨膜的糖蛋白。
3、 G蛋白偶联受体:是N递质、激素、肽类配体的受体。
1)特点:指配体与细胞表面受体结合后激活偶联的G蛋白,活化的G蛋白再激活第二信使的酶类。
通过第二信使引起生物学效应。
2)组成:由一条350-400个氨基酸残基组成的多肽链组成,具有高度的同源性和保守性。
3)G蛋白偶联受体作用特点:分布广,转导慢,敏感,灵活,类型多。
G蛋白偶联受体:G蛋白(由G蛋白偶联受体介导的信号转导)1)、G蛋白的概念:指鸟苷酸结合蛋白配体—G蛋白偶联受体—G蛋白2)、G蛋白的结构特征:①由α、β、γ3个不同的亚单位构成异三聚体(异聚体),β、γ二个亚单位极为相似且结合为二聚体,共同发挥作用。
细胞信号转导的四种途径
细胞信号转导是指外界刺激通过细胞表面受体传递到细胞内部的过程。
根据信号传递的途径和方式的不同,细胞信号转导可以分为以下四种途径:
1. 直接途径:有些信号分子可以直接通过细胞膜渗透到细胞内,与胞内的受体或靶分子结合,从而触发信号转导。
例如,甲状腺激素可以通过细胞膜渗透到细胞内,与核内的甲状腺激素受体结合,从而调节基因转录。
2. 离子通道途径:一些信号分子可以通过调节细胞膜上的离子通道的开关状态来转导信号。
例如,神经递质乙酰胆碱可以通过结合细胞膜上的乙酰胆碱受体,打开或关闭离子通道,引发细胞内的信号转导过程。
3. 酶级联途径:某些信号分子通过激活或抑制多个酶的活性来转导信号。
例如,酪氨酸激酶受体在受到外界信号刺激后,会自磷酸化并激活下游的信号转导酶,从而引发一系列的酶级联反应。
4. 细胞内信号传导通路:某些信号分子可以通过调节细胞内的信号传导通路来转导信号。
例如,细胞因子肿瘤坏死因子可以通过结合细胞膜上的肿瘤坏死因子受体,激活NF-κB信号传
导通路,引发细胞内的炎症反应。
细胞信号转导的途径不仅多样化,同时也会相互影响和交叉调控,从而产生复杂的信号网络,调节细胞的生理和病理过程。
细胞的信号转导是指外界信号通过细胞膜传递到细胞内部,触发一系列生化反应和细胞功能的调控过程。
细胞的信号转导可以通过多种方式进行,其中常见的几种方式包括:
1.直接通透型信号转导:某些小分子信号物质(如气体一氧化氮)、离子(如钙离子)或
水溶性小分子可直接穿过细胞膜,与胞浆内的靶分子发生作用,并触发相应的信号转导反应。
2.膜受体介导的信号转导:大部分信号分子无法直接通过细胞膜,而是通过与细胞膜上特
定的受体结合来传递信号。
这些受体可以是离子通道、酪氨酸激酶、鸟苷酸环化酶等类型的膜受体。
当信号分子与受体结合后,受体会激活下游的信号传递通路,如激活蛋白激酶级联反应或次级信号分子的释放,从而引发细胞内的信号转导。
3.细胞间接触介导的信号转导:有些细胞间信号传递是通过直接接触实现的。
例如,细胞
间的黏附分子可以通过细胞-细胞或细胞-基质之间的物理接触来传递信号。
这种方式通常使细胞与周围环境相互作用,调控细胞的形态、迁移和生长等过程。
4.核内受体介导的信号转导:某些脂溶性信号分子(如类固醇激素和甲状腺激素)可以通
过穿过细胞膜进入细胞,并与细胞核内的核受体结合。
与核受体结合后,信号分子与核受体复合物进入细胞核,影响特定基因的转录和表达,从而调控细胞功能。
这些信号转导方式可以单独存在,也可以相互作用,共同调节细胞的功能和生理过程。
不同的信号转导方式在细胞内部形成了复杂的网络,以确保信号的准确传递和细胞功能的精确调控。
植物Ca2+信号的研究进展摘要为了适应环境,调节自身代谢和生长, 在植物的生长发育过程中,需要对各种外界环境刺激以及植物内部生理信息做出反应,因此,植物产生了自己的信号系统。
Ca2+作为一种信号分子,它几乎参与了生命体所有的生理生化活动,在植物细胞的信号系统中也起着举足轻重的作用。
钙是植物生长发育必需的大量元素之一,在细胞水平上, 钙在细胞分裂、极性形成、生长、分化、凋亡等过程中均有重要的调节功能, 能维持细胞壁, 细胞膜及膜结合蛋白的稳定性并参与调节和控制植物的许多生理生化反应, 是植物代谢的重要调节者。
针对国内外对植物Ca2+信号的研究情况,综述了Ca2+信号的产生、Ca2+信号参与的各种植物生理过程、Ca2+信号的检测以及其研究的最新进展。
关键词:植物; Ca2+信号; 检测; 研究进展钙元素广泛存在于自然界和各种生物体内, 而游离态的Ca2+更是在生命活动中扮演着举足轻重的角色, 它几乎参与了生命体所有的生理生化活动。
作为一种信号分子, Ca2+在受精、胚胎发育、基因表达、细胞分化、组织形成、代谢调控等过程中都有参与, 可以说, Ca2+信号无处不在[1]。
1967年, Ridg-wang和Ashley通过向藤壶肌纤维中微注射水母发光蛋白, 第一次测定静息态胞内钙离子浓度[Ca2+]以来, 对于Ca2+信号的研究即风生水起。
虽然植物Ca2+信号的研究起步较动物细胞晚, 但依然取得了一些成果。
对植物Ca2+信号的研究, 不但能揭示生命的奥秘, 同时能帮助我们更加清楚地了解各种生命活动。
为此, 针对国内外对植物Ca2+信号的研究情况, 笔者对Ca2+信号的生理功能、信号的产生、Ca2+信号参与的各种植物生理过程、以及其研究的最新进展进行了综述。
1.Ca2+的功能Heilbrunn在1937~1952年发表的著作中, 提出了Ca2+在生物系统中复杂和多功能性的观点。
认为利用Ca2+是所有活细胞的基本特征。
在他提出的“细胞刺激理论”中认为:当细胞受到各种刺激时, 细胞内原来浓度很低的Ca2+水平明显增高。
Heilbrunn提出Ca2+的一些细胞效应有:(1)促进细胞黏合和胞间通讯;(2)影响酶活性, 如ATP酶酯酶等;(3)调节细胞分裂;(4)控制细胞的代谢活动;(5)调节细胞溶质中溶胶-凝胶状态转变;(6)高浓度Ca2+可能造成细胞死亡, 溶质中Ca2+浓度如果太高, 会与细胞内的磷酸根产生沉淀, 而磷酸根是细胞能量及物质代谢所必须的;(7)调节细胞膜的透性。
钙在维持细胞膜方面有着重要作用, 电镜观察表明, 缺钙导致细胞膜解体, 加钙又恢复常态。
可见钙有稳定细胞膜结构, 防止细胞膜损伤的作用。
有机酸是植物代谢的中间产物, 钙能和有机酸结合成为可溶性的钙盐结晶, 其中最为普遍的就是草酸钙。
据报道, 在外源Ca2+诱导下, 细胞内可形成草酸钙结晶移去外源Ca2+, 结晶会消失。
草酸钙的形成有以下生理作用:(1)消除有机酸在植物体内的过多积累。
(2)草酸钙的形成过程是可逆的,植物体内钙离子过多形成草酸钙, 消除过量钙对植物的伤害, 当钙离子浓度不能满足植物需要时,草酸钙释放出Ca2+以满足植物的需要。
2.植物Ca2+信号的产生和终止高度区域化的植物细胞内结构中, 在质膜液泡膜内质网膜上都存在着跨膜的钙离子电化学梯度, 细胞质和细胞核内游离钙离子也呈现不均匀分布, 这些梯度分布在静止状态是相对稳定的, 在受到刺激时会发生变化。
钙离子梯度是钙信号产生的基础,即植物细胞Ca2+空间分布的不均衡性是产生Ca2+信号的生物基础。
植物细胞中, 静息态的胞内Ca2+浓度([Ca2+] i)为100~200nM, 而细胞外(细胞壁)和细胞内(内质网、液泡、线粒体、高尔基体、细胞核)钙离子库中钙离子浓度却是胞内的数十倍, 达到了1~10mM[2,3]。
当细胞受到信号刺激时, Ca2+从钙离子库中释放, 使胞内Ca2+浓度瞬间升高,激活Ca2+依赖蛋白和激酶CPKs引起细胞代谢以及基因表达的改变。
当Ca2+重新进入细胞内钙离子库或流出细胞进入胞外钙离子库时, 信号得以终止。
钙离子浓度的调节是通过各种钙离子通道, 钙离子泵和钙离子转运来实现的[4]。
3.植物Ca2+信号的多样性Ca2+信号几乎参与了各种植物生理过程, 包括花粉管生长、细胞分裂、受精等;同时, Ca2+信号还参与植物的抗逆反应和对光线的感知。
由此可见, Ca2+信号在生命活动中的多样性和重要性。
在Digonnetetal. [5]的研究中发现,信号分子钙离子参与了植物精子与卵细胞融合;在Gehringet al.[6]得出Ca2+参与了植物的重力定向;在Hepler[7]和Messerli et al[8]的试验中发现,粉管的延伸, 根毛和不定根的生长以及细胞的延长中也有钙离子的转导作用。
在Knight et al[9]中,当有干旱胁迫时(Nacl),发现离子转运, K+浓度升高以及耐盐性和抗旱性也出现。
另外,在外界刺激下, 胞内钙离子增加, 这样细胞液中的钙受体蛋白(CaM)与Ca2+结合, Ca2+与钙受体蛋白结合后可激活相应的靶酶或靶蛋白,从而引起相应的生理生化反应。
当完成信息传递后, Ca2+又被迅速泵出胞外或被胞内储钙体吸收,细胞液中Ca2+又回落到静息态水平, Ca2+与受体蛋白分离。
通过这样的途径, 即依赖细胞液内Ca2+浓度变化而把细胞外的信息传递给细胞内各相关过程的功能, 称为Ca2+信号转导。
植物通过Ca2+信号转导途径来响应逆境胁迫, 发生一系列的生理生化反应, 从而使植物适应环境胁迫。
在植物处于低温、热激、缺氧、氧胁迫、病原菌的环境下时,都有钙离子信号转导的参与[10]。
4.植物Ca2+信号的研究新进展4.1 Ca2+与基因表达和植物生长MariaS.等发现, 在玉米发芽时, 生长素使细胞内Ca2+和H+浓度发生改变[11]。
Peng Gao 等人研究发现, 在棉花纤维细胞延伸生长时, Ca2+信号元件CaM和GAD等在纤维细胞中特殊表达, 提示Ca2+信号在细胞延伸时起重要作用[12]。
CAMTAs家族的研究, CAMTAs即CaM结合转录激活因子, CAMTAs家族成员有特异的DNA结合域CG-1, 该DNA结合域可以直接与DNA结合并激活转录, 或与其他转录因子作用共同激活转录。
CAMTAs家族在胚胎发生和调节生长过程中有重要作用[13]。
4.2 Ca2+与光Dolores等用红光和远红光处理离体黄瓜子叶发现, 组蛋白和内源多缩氨酸的磷酸化, 最终分离了75kD、57kD、47kD3种蛋白[14]。
这3种蛋白都能使组蛋白磷酸化并能自我磷酸化, 而且还发现, 75kD和57kD这两种蛋白是钙离子依赖型蛋白激酶。
植物体内Ca2+浓度会随光周期变化而昼夜波动, Ruhang Tang 等[15]发现, 这种波动与CAS-IP3相偶联, 提出并证明了Ca2+昼夜波动的模型。
4.3 Ca2+与植物损伤由生物和非生物因素引起的植物损伤反应信号通路中, Ca2+的生物学作用还不清楚。
研究发现,在番茄中, 损伤信号缩氨酸, systemin(一个18个氨基酸残基组成的多肽)引起细胞内Ca2+浓度升高。
这种胞内Ca2+的浓度变化是由胞外Ca2+内流和胞内钙离子库中Ca2+释放到细胞质中产生[16]。
通过检测Ca2+浓度和损伤反应, Dombrowski 和Bergey证实, 在番茄中, Ca2+通道在调控损伤反应的激活中起着重要作用。
另外还发现, Ca2+震荡和损伤基因的激活有着直接关系, 当Ca2+浓度增加systemin活性得到增强[17]。
在抗逆反应中, 植物通过改变细胞质内Ca2+浓度来抵御各种胁迫[18]。
最近发现, 在此过程中细胞核内Ca2+浓度也会变化[19,20]。
5.讨论与展望在过去的二三十年中, 植物Ca2+信号的研究虽然取得了巨大的成就, 但是同时也存在很多问题。
如在某一特定的生理过程中, Ca2+是如何起作用的;植物又是怎样通过Ca2+来调节其生长代谢的;还有众多Ca2+受体和钙依赖型蛋白激酶的功能;植物信号转导网络的完善等等问题都亟待解决。
随着检测技术的发展和研究的不断深入, 植物Ca2+信号转导途径中所存在的这些问题一定会逐步攻克, 与此同时, 对Ca2+信号转导的深入了解应用于生产也具有深远的意义。
参考文献[1] 王瑞云, 王玉国.钙在植物生理代谢中的作用[J] .世界农业, 2001, 6(266):41-43.[2]陈硕, 陈珈.植物中钙依赖蛋白激酶(CDPKs)的结构与功能[J] .植物学通报, 2001, 18(2):143-148.[3] 章文华, 陈亚华, 刘友良.钙在植物细胞盐胁迫信号转导中的作用[ J].植物生理学通讯, 2000,36(2):146-153.[4] 周卫, 汪洪.植物钙吸收、转运及代谢的生理和分子机制[J] .植物学通报, 2007, 24(6):762-778.[5] Digonnet, Aldon, Leduc, Dumas, Rougier.First evidenceofa calciumtransientinflowering plantsatfertilization[J].Development, 1997, 124:2867-2874.[6] Gehring, C.A., Williams, D.A, Cody, S.H., et al.Phototropismand geotropismin maize coleoptiles and spatiallycorrelated with increases in cytosolicfree calcium[J] .Nature, 1990, 345:528-530.[7] Hepler, P.K.Tip growth in pollen tubes:calciumleads theway[J].Plant Science, 1997(2):79-80.[8]Messerli, M.A., Creton, R., Jaffe, L.F., et al.Periodicincreases in elongation rate precede increases in cytosolicCa2+during pollentubegrowth[J].Dev.Biol., 2000, 222:84-98.[9] Knight, H., Veale, E.L., Warren, G.J., et al.The sfr6mutation in Arabidopsis suppresses low-temperature induction of genes dependent ontheCRT/DREsequencemotif[J].Plant Cell, 1999, 11:875-886.[10]尚忠林, 毛国红, 孙大业.植物细胞内钙信号的特异性[J] .植物生理学通讯, 2003, 39(2):92-100.[11]赵昕, 裴真明, 何奕昆.胞外Ca2+信号—动植物中的第一信使[J] .遗传, 2007, 29(3):269-275.[12]宗会, 李明启.钙信使在植物适应非生物逆境中的作用[J] .植物生理学通讯, 2001, 37(4):330-335.[13] 罗汉度.钙的生理作用[J] .植物生理学通讯, 1998,34(2):28-29.[14] 崔洪宇孙玉河.植物抗病的Ca离子信号转导的研究进展;天津农业科学;2007:48-51.[15] 宋秀芬, 洪剑明.植物细胞中钙信号的时空多样性与信号转导[J].植物学通报, 2001, 18(4):436-444.[16] 张宪政,陈凤玉.植物生理学[M] .长春:吉林科学出版社, 1996.[17] 简令成, 王红Ca2+在植物细胞对逆境反应和适应中的调节作用[J] .植物学通报, 2008,(3):255-267.[18] 林善枝, 张志毅.毛白杨幼苗低温锻炼过程中Ca2+的作用及细胞Ca2—ATP酶活性的变化[J].植物生理与分子生物学学报,2002,28(6):449-456.[19] 毛复军.钙和钙调素参与小叶锈菌互作的初步研究[ D] .石家庄:河北农业大学, 1995.[20] 孙大业.植物细胞信号转导研究进展[J].植物生理学通讯,1996.32(2):81-91.。