Toll
- 格式:pdf
- 大小:172.82 KB
- 文档页数:4
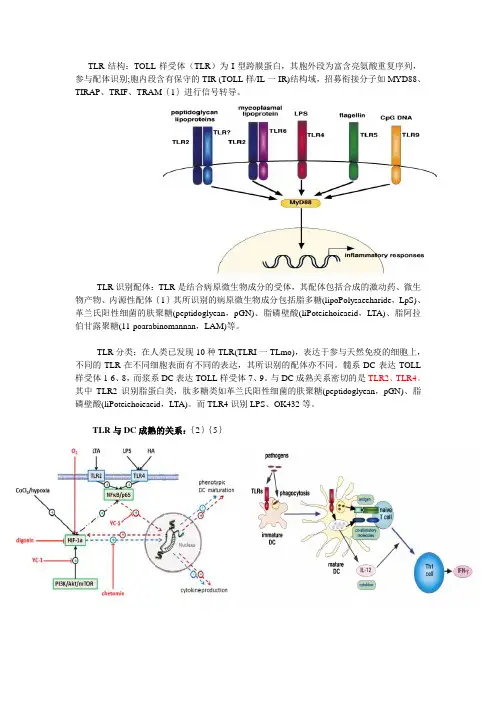
第6卷第1期 2002年3月 生命科学研究 lj Scienee Research v01.6 No l Mar 2002 Toll样受体研究进展 张德明,毛宝龄 (第三军医大学新桥医院全军呼吸中心,中国重庆400037) 摘要:T(Iu样受体(Toll-like receptors,11 )是新近发现的先天性免疫系统中的细胞跨膜受体I醍病原模式识 别受体之一,在急性是症反应细胞吞噬作用的调节和细胞信号转导I醍细胞凋亡中起重要作用简要综述了 Toil样受体的发现、分布、基因定位与结构特点、配体特异性I醍其介导的信号通路,并对其研究意卫与前号作 了简连. 关键词:1铷样受体;先天性免疫;盘症;信号转导 中图分类号:R392 文献标识码:A 文章编号:1007—7847{2002)01.OO36434 Recent Advances on Researches of Toll—like Receptors ZANG De-ruing,MAO Bao-ling (The ofRespiratoryDiseases PIA,妇聊 Hospital, 聊 430007,CA/rat) Abstract:Toll—like receptors,a kind of[1"dtns一12 ̄llbrane receptors and pathogen pattern recognition receptors,p1.dY important roles in dati“g cytoplu ̄, si al transduetion and apoptosis during an acute inflarmr ̄tion.Their dis・ eovery,distribution,gene locus,structure,ligands specificity.and signaling pathway were reviewed and also their re— search importance and developing trends in the future were b env discussed. Key words:Toll—like receptors;the innate immune system;inflammation;signal transduction (1. ̄fe Science Research,2002,6(1):036~039) 1 TLRs的发现 l3年前,人们在研究果蝇胚胎发育过程中, 发现有一个基因( l0l1)决定着果蝇的背腹侧分 化 ,其编码的一种跨膜受体蛋白称为Toll受体 (Toll Receptor).8年后.Lemaltre等 发现,dToll不 仅在胚胎发育中起重要作用,而且参与成蝇的免 疫反应,dTofl功能缺失的果蝇显示对真菌感染的 高敏感性,说明Toll受体具有介导抗真菌感染信 号转导的功能 Toll家族的另一成员18一wheel与 产生一种抗细菌肽attacin相关联 .此二者的发 现提示,Toll受体在抗真菌、细菌等病原感染中的 重要意义,同时也引起人们对人类Toll受体挖掘 的兴趣.Jmmway等 首次发现与dToll同源的人 hToll及编码的Toll样受体蛋白,并命名为TLR4. 到目前为止至少TLRs家族的10名成员已被发 现,其中TLR1一TLR5的结构已被确定,但只有 TIB2与TLR4的功能才部分揭示,TLR4主要介导 G菌感染LPS的信号转导,而TIB2主要介导G 感染的脂蛋白、脂多肽等的信号转导,它们都最终 导致NF- ̄B的转位与相应免疫基因的活化而转 录,释放前炎症因子及辅助刺激分子(Co-stimulate moleculars),起到调节炎症反应的作用.因而提示 1u 可能在先天性免疫系统(the innate immune system)中起重要作用 ,并成为联系后天性免疫 系统(the adaptive immune system)的纽带 收稿日期:20014)4-23;修回日期:∞ _01—06 基金项目:国家自然基金资助项目(3 3021O) 作者简介:张德明(1964.),男.湖南沅江凡,博士,主要从事急性肺损伤的研究. ;+864)23 ̄,8755320;毛宝龄t19"20-),男.河南 凡,第三军医大学一级教授,博士生导师.主要从事急性肺损伤的研究.
维普资讯 http://www.cqvip.com 第1期 张德明等:ToH样受体研究进展 2 TLRs的基因定位与结构特点 1998年R0ck等 用荧光原位杂交和标记后 序列位点数据分析法,研究1LR1—1.1J 的基因 定位.发现TLR1、TIAq2、TLB3分别位于4号染色 体的4p14、4q32、4q35,TLR4位于9q32—33,TLR5位 于lq33.3.但到目前为止,只有TLR4的基因被确 定,Poltorak等。 证明TER4是由内毒素基因(1ps) 所编码.相信其它TLRs基因将陆续被确定.TLRs 是一组跨膜受体蛋白,与dToll具有中等程度的同 源,更重要的是几乎享有相同的结构.包括胞外丰 富的亮氨酸重复序列(1eucine rich repeats,Llm)和 胞内与白介素一1受体信号域显著同源的尾状结 构域(tale domain),称为TdUIL-1同源域 TLR4 的LRR为22个拷贝791.LRR主要与识别特异性 配体——病原相关模式分子(pathogen-association pattern molecular)有关,如TLR4的胞外域就是用 来识别LPS;胞内的尾状域主要负责信号转导,如 TLR4的尾状域将LPS刺激信号传递给白介素.1 受体相关激酶(IL-1 receptor.associated kinase. IRAK). 3 11L 的分布 物种分布.虽然TLRs首次发现于果蝇,但其 尚分布于其他物种,从低等的植物如烟草 J、低等 动物小鼠、大鼠、仓鼠到高等动物人类都发现有 TLRs的存在,人们由此推测TLRs与物种生物进 化密切相关. 组织分布.Rock等 用mRNA斑点技术分析 了TⅡ 不同组织的分布情况.结果显示TLR1分 布广泛且比其他TLRs表达要高,短的3.0kb和长 的8。0 kb TLR1转录形式分别发现在卵巢和脾脏, TLR1还高表达于帕金氏淋巴瘤组织;TLR2不如 TLR1表达广泛,在肺组织可测到4。0kb TLR2 mR— NA的表达,4.4 kb则表达心脏、脑和肌肉;TLR3 呈现两个大约4.0 kb和6。0 kb主要转录子。最高 表达于胎盘和胰腺;相比之下,TLR4和TLR5显示 严格的组织特异性,TLR4仅以7.0 kh转录子表达 于胎盘组织,TLR5微弱的4。0 kb信号发现在卵巢 和外周血单核细胞.后来人们还发现TLR4尚可 表达在其他组织如肺 。 细胞分布.TⅡ 细胞分布广泛,2年时间的研 究发现,TLRs分布的细胞已达20多种。Muzio 等 对TIItl—TLR5在人白细胞的表达分布研究, 发现TLR1能在所有细胞包括单核细胞、多形核 细胞、T、B淋巴细胞、NK细胞中表达,TLR2、 TLR4、I"126只在髓源性细胞表达,TLR3只在树突 状细胞表达.提示TLRs可根据分布特性如广泛性 (TLR1)、限制性(TLR2、TLR4、TLR5)和特异性 (Tim)进行分类.TLR4尚广泛分布于其他组织细 胞.如人胚肾细胞(ttEK293) 、心肌细胞和微血 管内皮细胞“ 、人气道上皮细胞” 、肺巨噬细胞 和腹腔巨噬细胞 J、枯否氏细胞 、脂肪细 胞 和肠上皮细胞 :TIB2还分布在成纤维细 胞 、星形胶质细胞 及巨噬细胞 和树突状 细胞圳. 4 TLRs配体的特异性 Tills是先天性免疫模式识别受体(pattem recognition receptor)之一,其相应受体具有特异性, 发现TLRs特异性配体是TLRs研究的热点之一. 近两年主要发现的是TLR4与TLR2的特异配体 TER4的配体主要是内毒素(LPS)中的类脂A , 其次有抗肿瘤药物泰素 、内源性配体热体克蛋 白60( ̄p6O) 等.TER4配体的病原也具特异性, 主要有G一菌、厌氧菌 、致密螺旋体 】.TLR2的 配体较TER4广泛,包括脂蛋白、脂多肽、脂壁酸、 阿糖甘聚糖脂(1ipoarabinomannm ̄LAM)、酵母多 糖 、细菌总的DNA(the capacity of baeteria DNA CpG DNA) .Underhill 等认为TLR2配体广泛 多样性,原因:1)TLR2本身可直接结合这些受 体;2)TLR2结合负责特异区别配体的内源性巨 噬细胞蛋白;3)TLR2可能与其他TLRS聚合.从 而增加配体结合靶位TLR2配体的病原也较 TLR4广泛,目前发现的有G 菌如葡萄球菌和肺 炎链球菌 致密及疏螺旋体、支原体、分枝杆 菌 、酵母菌 、大肠埃希氏 等.相信随着 TLRs的深入研究.将不断有新的TLRs配体及病 原被人们认识。 5 lIARs介导的信号通路 目前只有TLR4和吮介导的信号通路部 分明确。参照B ll 以_兀 4介导LPS的NF- B通路为例说明。LPS与血清中LPS结合蛋白 (LBP)结合后再与CD14形成复合物,此复合物中 的LPS解聚后与TLR4结合导致TLR4的聚合而 活化 ,在一种分泌蛋白MD-2的辅助下 活化的 TI
lt4胞内尾状结构与接头蛋白MyDSS的TLRs受 维普资讯 http://www.cqvip.com 生命科学研究 2∞2年 体域结合,MvD88通过其死域(death domain)再与 IRAK结合,二者作用导致IRAK的自身磷化,从 而激活肿瘤坏死因子受体相关因子.6(TRAF一6, TNF—d receptor association factor 6),使有丝分裂原 结合蛋白激酶(MAPK)家族活化,其中NF-tcB诱导 激酶激活I, B家族n、 激酶,导致I-xB家族的 广泛磷酸化而降解,使NF—xB转位到胞核,其活 性二聚体启动细胞因子如(ILl、IL-6、IL-8、IL一12 等)和辅助刺激分子CD80和CD86基因的转录 以上是TLR4介导LPS活化NF,xB信号公认的通 路,但也有报道LPS无需与LBP和CD14直接结 合TLR4 TLR2介导的信号通路大致与TLR4相 同,但尚未发现MD一2类似蛋白及其作用.除NF— xB信号通路,其他信号通路也发现与TLR4密切 相关 ”.TLR4主要活性形式(dominant active form) 过度表达不但激活NF—xB,而且也激活AP1及Jun N端激酶( K),MAPK尚可激活TLRs下游的其 他通路,不过仍不十分清楚.此外,B细胞特异性 TLR RP105,可激活 家族酪氨酸激酶Lyn、蛋白 激酶C-p及MAP激酶通路 j.相信将不断有新的 证据,证明TLRs激活更多不同信号通路,有助于 阐明基因活化和细胞炎症反应的调节作用. 6 TLRs的研究意义及展望 机体在B、T细胞克隆选择并对病原产生反 应前,尚需3~5 d时间,先天性免疫系统是此其 间机体主要的防御体系,包括分泌抗微生物肽、细 胞吞噬作用和多种补体途径的活化,用以快速控 制感染性病原的复制 .Ⅱ 是先天免疫模式识 别的主要受体,通过识别病原相关模式分子,激活 先天免疫系统产生前炎症因子、抗微生物肽抵御 病原所致损害,同时作为预警信号,向抗原递呈细 胞发出警报,从而启动后天免疫系统.故Patti Godowki认为Ⅱ 领域的研究具有划时代的意 义 .除急性的微生物感染外,Ⅱ 还在慢性炎 症如哮喘及自身免疫性疾病如关节炎等方面具有 重要的研究价值.Ⅱ 抗感染机制主要有三个方 面”:一是激活iNOS抗炎;二是介导细胞凋亡, 下调感染反应性,减轻组织损伤;三是介导释放抗 炎因子如ID10、IL-12、ll,4等.人们还发现TLRS 的点突变如TLR4 712 bp突变使组氨酸代替脯氨 酸后,机体对LPS刺激的敏感性降低 ,这为生产 治疗c菌感染后内毒素血症的ⅡRs基因生物制 剂,提供了理论基础和契机. 目前TLRs热方兴未艾,今后的研究内容众 多,主要可能有以下几个方面:TLRs家族新成员 的发现及其结构与功能的阐明;新的TLRs配体的 不断挖掘;TLRs在物种、组织、细胞的新分布; TLRs与细胞吞噬和细胞凋亡的调节;新TLRs信 号通路的发现lTLRs家族成员之间功能上的互相 联系与作用;以及有关TLRs药物与疫苗的研制与 开发等. 参考文献(R曲 lIoes): 1 J HASHIMOID C HUDSONKL.?d' ̄DRSONKV.Thetoll gene dmsq ̄la r 【LI red r drosol-,' ̄tral ar1b …pol ̄l:,appears to em.ode a lrand—n ̄lJOet33tlle protein[J] -【.1998.52:Ⅻ.279 l2 u ¨mE B、N1CO1.AS E,MICHAUT L. The drosol…一 tral regulto ̄, e c 8eIle Spotzle/TaU/Catus ̄ltrols the potent  ̄ltihmg.al m in dro ̄i La adults[J: _I-t996,86:973— 983 [3 ' ̄ILLIAMSM J.RODRIGDEZ A.KIMBELL D A.et a/The18一 wh 【rr ̄ation ̄'eals cⅢl】 ex amJb ̄.tefi*l 8嘶e Te mⅢin drosophila hoa tjeI哪ce[J]FJ, ̄O J.1997.16:6120-6 L30. 【4 1 MEDZHITOY R,PRES ̄ON H P.JA_NAWAY C A A h山咖ho.  ̄togae of山 la Toll protein Erlals咖vml帆of adaptive im- munJly【JJ H l .1997.388:394-397 l5]GRETCHEN V Fly development es lead to immune findlJ]. sc….I998.25(281):1942-1944. 6J ROCK F L.HARDlMANG.TIMMS J C a/Afamily of hmmn oe m stmc.qrare Ly ̄lated Io drosophia Toll:J] Natl Acad Scl USA.1998.282}588-593. 。7]POLTOI ̄.K A.HE X,SMIRNOV A,d a/.Def ̄'tive LPS la【一 in C3H/HeJⅢd C57/10ScCrmice:raumfioninlI1lH4 晒ence,l998.282:588-593. 18J KOPG E B.MEDZHITOY R The Toll mc D fmnl and c ̄trol innate immunity.J]cu Opin Imrauno1.1999.I I:L3-I8 【9J WPJGHT S D Toll,Ⅱ new eceinthe ptmle innate…. ty J J J EXP MED,I999.I89(4):605—609 【10 J KLEEBERGER S R.REDDY S.ZHANG I Y. a/Gene ̄ic  ̄ptibility Io Ozone-inducnd lung hypermeabi i,rote of tof[- like receptor 4【J J Am J Respir Cell Md Bid.2(100,22:620- . 【11 MUZIO M,BOSISI O.11} u 卜r 咖C N. …一 糊0n and regulali ̄of toll—like recept ̄(TtRS)in h…leu k帆 :sdecIi pres目恤 TLRS in dendric -k J]+J Im- Ⅱ岫 .2000.164:59984:004 .12J JESSE c C,DONTqA W ' ̄roNG,DOUL ̄S T G,d Tol1.1ike r D mediate lipopoly ̄hafide-indm'ed signal transdm ̄tion [¨J Biol .1999,274(16):10689.10692 【13J STEFANF,LEATERK.YONG D K. a/.ToIJM.TLR4in d - y0c 部in nort r ̄.ridfai Li my.oca ̄lium J]J C]inInvest, l999 104:271-280 【14J NAUCY C A,Ev^L,BP.L ̄"I C S,d TLR4删 …