植物氮素营养诊断的进展与展望
- 格式:pdf
- 大小:144.52 KB
- 文档页数:5


植物营养诊断方法研究进展一、概述植物营养诊断,作为现代农业管理的重要环节,对于精准指导施肥、提高作物产量和品质具有不可替代的作用。
随着农业科技的不断进步,植物营养诊断方法也在不断创新和完善,为农业生产的可持续发展提供了有力支撑。
传统的植物营养诊断方法主要依赖于土壤分析和植物组织分析,通过对土壤和植物样本的化学元素含量进行测定,以判断植物的营养状况。
这些方法存在操作繁琐、耗时长、对样本破坏性强等缺点,难以满足现代农业高效、精准的需求。
随着科技的发展和研究的深入,植物营养诊断方法取得了显著进展。
新的诊断方法不仅提高了诊断的准确性和效率,还减少了对植物和土壤的破坏。
无损测试技术可以通过对植物叶片或冠层的光谱反射率进行分析,快速准确地评估植物的营养状况;生理生化分析诊断法则可以通过测定植物体内的酶活性、代谢物含量等生理生化指标,揭示植物营养状况的生理机制。
还有一些新兴的诊断方法,如基于机器学习或深度学习的智能诊断系统,可以通过对大量数据进行学习和分析,自动识别和预测植物的营养问题。
这些方法的出现,为植物营养诊断提供了更多的选择和可能性。
植物营养诊断方法的研究进展为现代农业的发展带来了新的机遇和挑战。
随着科技的不断进步和研究的深入,我们有理由相信,植物营养诊断方法将更加精准、高效、环保,为农业生产的可持续发展做出更大的贡献。
1. 植物营养诊断的重要性植物营养诊断是现代农业科技中的关键环节,对于提高作物产量、改善农产品品质和保护环境具有重要意义。
随着农业生产的不断发展,植物营养问题日益凸显,如何通过科学有效的方法进行营养诊断,从而合理施肥,成为当前农业研究的重要课题。
植物营养诊断有助于精准把握作物生长过程中的营养需求。
作物在不同生长阶段对营养元素的需求各异,通过营养诊断可以了解作物当前的营养状况,进而制定针对性的施肥方案,避免过量或不足施肥造成的资源浪费和环境污染。
植物营养诊断有助于提高作物抗逆性和产量。
作物在生长过程中常受到病虫害、干旱、盐碱等不利因素的影响,通过营养诊断可以及时发现并纠正作物营养失衡问题,增强作物的抗逆性,从而提高产量和品质。


α文章编号:10083464(2003)03022505作物对氮素养分高效吸收的根系形态学研究进展史正军1,樊小林2(1 深圳市园林科学研究所,广东深圳518003;2 华南农业大学资源环境学院,广东广州510642)摘要:本文综述了近年来与作物对氮素养分高效吸收有关的根系形态学研究情况,简要比较了解释氮素供应诱导作物根系形态变化的几种观点,着重阐述了用氮素诱导碳水化合物定向分配来解释根系变化的合理性和意义,并提出了根系研究在水分、养分效率和植物营养遗传研究中的展望。
关键词:氮素养分;高效吸收;根系形态学中图分类号:Q 948;S 511 文献标识码:AProgress i n root m orphology for n itrogen eff i c i en t acquisiti on i n cropSH I Zhengjun 1,FAN X iao lin2(1 Shenzhen In stitute of L andscape Garden ing ,Shenzhen 518003,Ch ina ;2 Co llege of N atural R es ources and Environm en t Science ,South Ch ina A gric .U n iv .,Guangzhou 510642,Ch ina )Abstract :T he paper summ arizes recen t p rogress in study on roo t mo rpho l ogy related to n itrogen effi 2cien t acquisiti on ,and briefly compares differen t hypo thesizes used to elucidate the phenom enon that roo t grow th can be induced by n itrogen supp ly .R ati onality and sign ificance of app lying the hypo thesis that n i 2trogen m ay induce tran s po rt and all ocati on of carbohydrate thereby induce the grow th of roo t are exp licated here .Pers pective of roo t research in the nutrien t efficiency and p lan t nutriti on hereditis m is p resen ted .Key words :n itrogen nutrien t ;efficien t acquisiti on ;roo t mo rpho l ogy氮素养分是农作物需求量最大的营养元素,氮的供应制约着作物产量的高低。

植物营养研究现状与未来趋势分析植物营养研究是农业领域中重要的一项研究方向,它关注的是植物的生长发育过程中所需要的营养物质及其吸收与利用方式。
随着人口的增长和资源的有限化,植物营养的研究越来越受到重视。
本文将回顾并分析当前植物营养研究的现状,并探讨未来的研究趋势。
目前的研究已经明确了植物所需的主要营养元素,包括氮、磷、钾、硫、钙、镁和微量元素。
研究表明,这些营养元素对于植物的生长和发育至关重要。
然而,随着人们对植物生长环境的进一步了解,越来越多的研究表明,除了基本营养元素外,植物还需要其他非营养因子的支持,如植物激素、有机酸等物质的调控。
在植物营养研究的现状中,一项重要的研究方向是植物对不同营养元素的吸收和利用机制研究。
这一方面的研究主要关注植物根系的发育和功能,以及植物与土壤的相互作用。
目前的研究表明,植物根系的形态和结构对于提高植物对营养元素的吸收能力至关重要。
同时,土壤中的土壤微生物和土壤化学性质也影响着植物对营养元素的利用效率。
另一个重要的研究方向是植物营养在农业生产中的应用。
随着全球化的发展和粮食安全的挑战,农业生产需要更高效、更可持续的方式来满足不断增长的需求。
植物营养的研究可以为农业生产提供科学依据和技术支持,例如通过优化植物对营养物的吸收和利用,可以提高农作物的产量和品质,减少肥料使用量,并减少对环境的污染。
未来的研究趋势将主要集中在以下几个方面。
首先,随着人工智能和大数据的发展,植物营养研究将更多地运用这些技术来提高研究效率和数据分析能力,更全面地理解植物营养的机制和调控网络。
其次,植物根际微生物的研究将成为未来的一个重点领域。
根际微生物与植物之间存在着复杂的互作关系,它们可以促进植物对营养物的吸收,提高植物的抗逆性和生长发育。
通过深入研究植物根际微生物的功能和机制,可以为农业生产提供新的思路和方法。
另外,基因编辑技术的发展也将推动植物营养研究的进展。
通过精确修改植物的基因,可以优化植物对营养元素的吸收和利用能力,提高作物的产量和营养品质。

氮素在植物中的利用综述
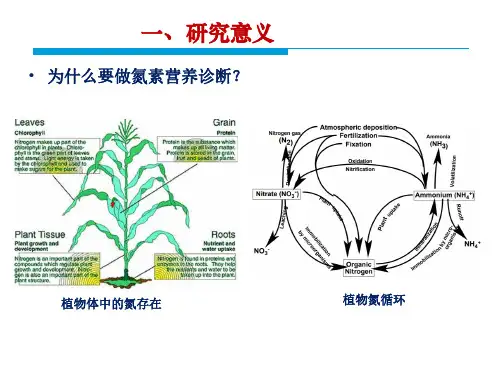
氮素是植物生长发育过程中必不可少的营养元素之一,它参与了蛋白质、核酸、酶等重要物质的合成,是植物生长发育的关键因素。
在自然界中,氮素以硝酸盐和铵盐的形式存在于土壤中,植物通过根吸收的方式获取土壤中的氮素。
氮素对植物的影响主要体现在以下几个方面:
1. 促进植物生长:氮素是植物体内最大的元素之一,参与了蛋白质、核酸等生物大分子的合成,是维持植物正常生长发育的重要组成部分。
缺乏氮素会导致植物生长缓慢、体型矮小、叶片黄化等表现。
2. 提高植物的抗逆能力:氮素可以增强植物的抗病虫害、抗胁迫能力,提高植物的适应性。
适量的氮素供应可以增加植物体内抗氧化酶、保护酶的活性,提高植物对逆境的抵抗力。
3. 促进植物的开花结果:氮素是植物体内调节花芽分化和开花的重要因子。
适量的氮素供应可以促进花芽分化,提高植物的开花率和果实的质量。
在农业生产中,氮素的供应对提高作物产量和品质至关重要。
过量施氮易导致农田环境污染和氮素的浪费,而缺乏氮素则会限制作物的生长和产量。
为了合理利用氮素,减少氮素的浪费和环境污染,农业生产中广泛采用了一系列氮素管理措施,包括合理施肥、控制氮素损失等。
调整施氮量的时间和方式,采用分蓄封闭灌溉、覆膜等措施,可以减少氮素的挥发和淋失,提高氮素利用率,降低对环境的污染。
还可以通过通过调整土壤中的微生物群落结构,促进土壤中硝化还原过程的平衡,提高氮素的有效利用率。
对氮素的合理利用和高效供应,对于促进植物生长发育,提高农作物产量和品质,减少环境污染,具有重要的意义。
研究氮素在植物中的利用机制,对于优化氮素管理策略,提高农业生产效益具有重要的指导意义。

2015年第07期□李悦书关于作物氮素营养诊断方法的研究摘要:氮素作为构成活体生物组织的基本元素之一,在生物体内有着维持生命体征,控制生命系统的内部动态平衡有重要作用。
改善和改进对于作物氮素的管理方法,对于农作物的高效丰产和生态环境的保护都有重要意义。
文章根据近年来的研究和相关报道,总结了各种作物氮素的诊断方法的优点和不足,指出了今后氮素诊断方法的发展趋势,并展望了国内发展前景。
关键词:氮素管理;环境保护;诊断方法;发展趋势文章编号:1004-7026(2015)05-0018-01中国图书分类号:文献标志码:A(东北农业大学农学院黑龙江省农业科学院黑龙江哈尔滨150086)1几种常见的氮素营养诊断方法1.1传统诊断法传统的氮素营养诊断法,多用于对小区域内的土壤氮素成分进行分析,该诊断法的优点是简单易操作,而且取样简单,通过对试验田内部的少量土壤进行随机抽样,在实验室内借助简单检测仪器就可以完成。
缺点是测试周期长,即使对微小区域内的土壤进行检测分析,通常也需要花费数周时间才能得出结论。
另外如果需要进行深度分析,受限于使用仪器和检测条件,也会使使用结果不甚明显。
随着科学技术手段的发展,传统的氮素营养诊断法已经逐步被新兴的诊断法取代,目前只有在相对简陋的条件下才会使用。
1.2肥料窗口法此种方法的操作流程是:首先选取一大面积的被测试验田,在试验田中央或随机抽样部位,画出一小片的“微区”,通过对“微区”内作物的氮元素吸收程度和氮素含量进行跟踪监测。
如果“微区”内实验对象出现缺氮现象,则表明整个试验田初中缺氮的边缘,需要及时补充氮肥。
肥料窗口诊断法操作也相对简单,而且适用性更加广阔,尤其是在我国的平原地带,常常用该方法做初步的氮素检测。
缺点是如果试验田内缺氮地区分布不均匀,而选取的“微区”避开了缺氮区域,就会造成检测的误判,从而影响后续的改善措施。
1.3作物症状诊断法大量实验表明,植物缺少氮素就会表现出枝叶枯萎的症状,缺钾就会是叶子变黄,所以经验丰富的科学工作者,可以根据植株表现出的症状对作物进行大致的病情诊断。


实验报告-氮素对植物生长的影响(已评)
引言
本实验旨在研究不同氮素浓度对植物生长的影响。
氮素是植物生长所必需的主要营养元素之一,但过高或过低的氮素浓度可能会对植物生长产生负面影响。
通过本实验,我们将探究不同氮素浓度对植物生长的影响程度,从而提供合理的氮素施肥策略。
实验方法
1. 选取相同品种的植物作为实验对象
2. 将植物分为若干组,每组设置不同浓度的氮素处理
3. 每组设置多个重复样本,以减少实验误差
4. 按照规定浓度施加氮素,并记录植物生长的相关指标
5. 在一定时间后,测量植物的生长高度、叶片数量、根系长度等指标
6. 对实验结果进行统计分析和比较,以获取不同氮素浓度下植物生长的差异
实验结果
实验结果显示,不同氮素浓度处理对植物生长有显著影响。
随着氮素浓度的增加,植物的生长高度、叶片数量以及根系长度呈现出先增加后减少的趋势。
然而,过高的氮素浓度会导致植物生长不良甚至枯萎,而过低的氮素浓度则会限制植物的养分吸收和生长速度。
结论
根据实验结果分析,合理的氮素浓度对植物生长至关重要。
适度的氮素浓度可以促进植物的生长和发育,而过高或过低的氮素浓度都可能会对植物产生不良影响。
因此,在实际种植过程中,应根据具体植物品种和生长环境,科学施肥,以确保植物获得适宜的氮素供应,从而实现更好的生长效果。
参考文献
(请根据实际引用文献情况填写)。

植物的矿质与氮素营养植物是我们日常生活中不可或缺的一部分,它们为我们提供了许多重要的物质和服务。
植物的生长和发展取决于其对矿质和氮素的吸收和利用。
本文将讨论植物的矿质与氮素营养。
一、植物的矿质营养矿质营养是指植物从土壤中吸收的无机营养元素。
植物需要吸收的矿质元素有很多,包括镁、钙、铁、锌、铜、锰、硒、钼、氯等。
这些矿质元素在植物的生长和发展中起着重要作用。
1. 镁镁是植物体内的重要成分之一,它参与了许多生物化学反应,如光合作用和呼吸作用等。
镁的缺乏会导致叶片中叶绿素含量降低,影响植物的光合作用和生长发育。
2. 钙钙是植物细胞壁和中枢神经系统的组成成分,它对细胞分裂和细胞壁稳定性有着重要的作用。
钙的缺乏会导致植物的胶质变化,影响其正常生长发育。
3. 铁铁是植物体内的重要元素,它存在于许多酶中,参与了氧化还原反应和电子传递过程。
铁的缺乏会导致植物叶片的黄化,严重时可能导致植物死亡。
4. 锌锌是植物生长和发育的必需元素之一,它促进植物的生长发育和增强植物的免疫力。
锌的缺乏会导致植物叶片出现白色黄斑、萎缩等现象。
5. 铜铜是植物体内多种酶的组成成分,它对植物机体有重要的作用。
铜的缺乏会影响植物的代谢和生长发育。
二、植物的氮素营养氮素是植物生长必需的主要成分之一,植物需要从土壤中吸收氨、硝酸盐等氮源物质。
氮素对植物的生长发育有着重要的影响。
1. 生物固氮蚯蚓、田间杂草、青苔等具有固氮作用的微生物,能够把空气中的氮分子转变成可被植物吸收的氨态氮,为实现土地生态平衡起到了重要的作用。
2. 植物对氮素的吸收和利用一般情况下,植物吸收的氮素主要以硝酸盐形式存在。
植物的生长发育需要合适的氮素浓度。
氮素过多或不足都会影响植物的生长和质量。
3. 氮素对植物品质的影响植物体内氮素含量的增加,能够促进植物的生长发育和增加产量,但同时也会导致产量质量的降低。
植物倾向于把氮素转移到叶子和果实中,而不是转移到根系中,导致根系生长不良。
中国生物固氮研究现状和展望沈世华荆玉祥*(中国科学院植物研究所中国科学院光合作用和环境分子生理学重点实验室, 北京100093. *联系人, E-mail: yxjing@ )摘要生物固氮是生命科学中的重大基础研究课题之一, 它在生产实际中发挥着重要作用: 为植物特别是粮食作物提供氮素、提高产量、降低化肥用量和生产成本、减少水土污染和疾病、防治土地荒漠化、建立生态平衡和促进农业可持续发展. 本文在介绍国际生物固氮研究进展的同时, 着重叙述了生物固氮研究取得的重大进展和成果: 收集了根瘤菌资源, 建立了最大的数据库, 修正和发展了国际上对根瘤菌的分类; 发现了固氮基因, 证实了克氏杆菌固氮基因操纵子的连锁性及正调控基因的调节机制和对氧、温度的敏感性; 发现苜蓿根瘤菌结瘤调控基因nodD3的产物对结瘤基因表达的启动不受宿主类黄酮的作用; 发现苜蓿根瘤菌的碳利用基因和固氮生物氮代射和碳代谢基因表达及其调节的偶联作用; 化学合成了根瘤菌的结瘤因子; 在固氮基因表达调节基础上, 构建了固氮基因工程菌株, 并在生产中得到应用; 提出了化学模拟固氮酶的结构和功能, 固氮酶活性中心的模型和合成了模型化合物, 受到了国际高度评价. 根据国际上研究的趋势并结合国内的研究进展, 提出了生物固氮研究的发展方向, 建议在联合(内生)固氮菌固氮基因调控及其提供氮素的作用, 根瘤菌与豆科植物共生结瘤固氮的信号传递和分子相互作用, 氮、碳代谢和固氮与光合作用的偶联与共生结瘤固氮中功能基因组学等方面展开积极研究.关键词固氮生物固氮酶基因表达化学模拟微生物与植物相互作用功能基因组空气中约80%的氮气不能被植物直接利用, 只有固氮微生物具有将氮气转化成氨的能力, 人们称为生物固氮. 据联合国粮农组织(FAO)1995年粗略估计, 全球每年由生物固定的氮量已近2 × 106t(相当于4 × 108 t尿素), 约占全球植物需氮量的3/4. 所以, 生物固氮是地球上最大规模的天然氮肥工厂. 但是, 迄今为止所发现的绝大多数固氮微生物均不能在粮食作物水稻、小麦、玉米以及多种果树、蔬菜上固氮, 即使少数可以的话, 其固氮量也很少, 所以这些植物的高产不得不依赖化学氮肥. 30年后我国人口将达到16亿, 年需粮食6.4 × 108 t, 总计需尿素64 × 108 t. 按此需要, 至少还要新建很多氮肥厂, 投资上千亿元. 一方面, 适量使用化学氮肥可使粮食高产; 另一方面, 生产化学氮肥要大量消耗能源, 加重大气污染和温室效应. 大量施用化肥, 不仅提高农业生产成本, 而且导致水土污染, 影响健康和破坏生态平衡. 对于提高农业产量, 降低化肥用量和农业生产成本, 减少水土污染和疾病, 治理占我国国土面积约27%的荒漠化地区, 发展可持续农业, 生物固氮将起重要作用.研究生物固氮的作用机制有3个目的: (1) 提高固氮效率, 在理论上阐明影响固氮效率的原因, 在生产实际中提出有效措施; (2) 在研究根瘤菌与豆科植物相互作用和共生固氮的基础上, 扩大根瘤菌的宿主范围, 使其能在非豆科植物, 特别是主要粮食作物上固氮, 或将固氮基因转移到非豆科植物上, 实现其自主固氮; (3) 在研究固氮酶结构与功能的基础上, 进一步探讨化学模拟固氮酶作用机制, 发展化学催化理论, 改革目前合成氨工艺, 提供廉价氮肥.生物固氮是生命科学的重大问题之一, 是跨世纪的研究课题. 在当前生命科学的发展中由于基因组学和功能基因组学的建立和高新技术的创新, 又赋予生物固氮研究新的内涵和研究策略, 为实现固氮研究的目标增添了新的动力.本文叙述生物固氮的研究现状和发展, 着重介绍我国的研究概况和取得的成果, 并结合当前生命科学的进展, 展望生物固氮研究的前景.1生物固氮的研究现状当前, 国内外生物固氮研究已进入一个新阶段, 其特点是多学科交叉, 将基础研究和应用前景相结合, 开拓了思路. 当前生物固氮研究正在分子和原子水平上开展, 如: 固氮基因表达的铵阻遏和氧敏感机制; 共生结瘤固氮中植物与微生物相互关系的基因表达和调控; 根瘤菌结瘤因子的结构和生物合成; 根瘤菌及其宿主植物的基因组学、转录组学和蛋白质组学; 固氮酶的结构和功能及其化学模拟; 固氮效率的提高及其在农业和环境保护中的应用等. 这些研究要求生物学、农学、化学和物理学等学科的交叉和结合, 引入新概念和新技术, 综合进行.1.1固氮资源的发掘和应用生物固氮系统分为根瘤菌与豆科植物的共生结瘤固氮系统、联合(包括内生)固氮系统和自生固氮系统. 在共生固氮系统方面, 世界上有豆科植物19700种, 其中已知可以结瘤固氮的有2800多种, 占15%, 而对其共生固氮体系进行过研究的只占0.5%[1]. 不少国家, 特别是美洲和非洲国家, 积极发展种植大豆或其他豆科植物(美国大豆播种面积约占30%), 以发挥生物固氮作用, 减少化学氮肥用量, 取得了明显的经济效益. 在对联合(内生)固氮系统的研究中, 发现禾本科植物甘蔗内有内生固氮菌, 以光合产物为能源进行固氮, 可为甘蔗提供60%的氮素[2]. 这一发现为进一步开发联合(内生)固氮体系提供了突破空间和潜在的应用前景. 在自生固氮体系中, 发现一株嗜热放线菌(Streptomyces thermoautotrophicus)有耐氧的固氮酶[3], 为最终通过转基因手段实现非豆科植物自主固氮提供了可能的突破点.我国传统农业耕作过程中采用豆科植物与其他农作物套种、轮作等手段改良土壤环境, 提高农作物产量. 当前, 苜蓿等豆科植物在我国西部地区的开发及开展生态农业、退耕还林还草过程中正起着不可替代的作用. 我国共有豆科植物约1400多种. 多年来, 我国科学家以豆科植物根瘤菌为重点, 逐步摸清了我国豆科植物的根瘤菌资源, 进行了系统分类, 发现了一些新属、新种[4~7], 并建立了我国最大的根瘤菌数据库. 其中一个重要的发现是, 一种植物在不同的生态环境可与多种根瘤菌共生, 例如我国的大豆可与3个属、7个种的根瘤菌共生固氮, 而一种根瘤菌(如海南根瘤菌Rhizobium hanaese)可从13 属14种豆科植物的根瘤中分离. 其他很多植物与根瘤菌的关系也是如此. 这一研究说明豆科植物与根瘤菌共生的多样性, 修正并发展了传统的根瘤菌“寄主专一性”和植物“互接种族”的概念. 将为利用现代基因组学、功能基因组学和蛋白质组学手段, 探索最佳的结瘤固氮模式和微生物与植物相互作用的机理提供良好的研究材料.1.2生物固氮调控机理及植物与微生物的相互作用用自生固氮菌——克氏肺炎杆菌(Klebsiella pneumoniae) 研究固氮基因及其表达和调控机理, 有很多开创性工作, 如发现了固氮基因nifC,7个固氮基因nif操纵子连锁以及正调控基因nifA的调节机理及其对温度和氧的敏感性[8~10]. 豆科植物与根瘤菌之间的分子对话机理研究有了重大进展. 在能量供应方面, 弄清了根瘤菌在豆科植物根瘤中依靠植物提供四碳二羧酸糖作碳源用于固氮, 并发现了dctABD基因[11]; 重组根瘤菌已经构建成功, 并用于提高固氮效率[12,13]; 与粮食作物联合固氮的固氮螺菌(Azospirillum brasilence Yu62)的固氮调节机理也已逐步明朗, 为构建铵阻遏条件下也能固氮的基因工程菌株打下了理论基础[14]. 我国科学家在深入研究正调节基因(nifA)的表达及其产物(NifA)活性调节机制的基础上, 构建了不受铵阻遏的组成型表达的nifA质粒, 将其引入大豆根瘤菌(Bradyrhizobium japonica)和阴沟肠杆菌(Enterbacter cloacae)后, 固氮作用不受铵阻遏[15~19]. 用此基因工程菌株接种水稻可以增产[20,21]. 此外, 还开展了固氮基因负调节基因(nifL)的研究[22,23]. 我国分离的巴西固氮螺菌Yu62的固氮酶基因表达和活性双重调节机制研究已经比较清楚, 为构建耐铵泌铵的基因工程菌株和降低玉米化肥用量提供了理论基础[24~26]. 田间实验结果有实效, 施用工程菌株比不施用的对照增产21.1%, 比野生型菌剂增产8.5%. 在相同产量水平下可降低20%的氮肥使用量. 在共生固氮体系结瘤固氮基因表达调节研究的基础上, 发现苜蓿根瘤菌结瘤基因nodD3的表达不受苜蓿类黄酮物质的启动[27,28], 为扩大根瘤菌的宿主范围提供了理论根据. 同时, nodD3基因表达受到两个启动子的控制[29], 第2个启动子可以被NtrC激活[30]; 化学合成了苜蓿根瘤菌的结瘤因子[31]; 根据宿主植物对根瘤菌识别因子和固氮嫌氧机制的研究, 将豆科植物的凝集素基因和血红蛋白基因转入烟草和水稻, 获得基因表达, 为进一步研究非豆科植物和根瘤菌侵染的关系奠定了技术基础[32,33]. 近年来, 我国开展了特有的华癸根瘤菌(Sinorhizobium huakui)结瘤固氮基因表达调节的研究[34,35], 发现了微生物体内碳代谢与固氮及氮代谢的基因表达调节之间存在着偶联关系[36,37]. 这一发现不仅对生物固氮调控有重要意义, 也对基因表达调控基础研究有重要贡献, 为进一步研究光合和固氮之间的偶联提供了理论基础.1.3固氮酶的生物化学特性及其化学模拟国际上已经对固氮酶高分辨率的空间结构进行了研究, 阐明了其活性中心的原子簇FeMoco及其周围蛋白分子的三维结构[38,39]. Schmid等人[40]对棕色固氮菌缺失FeMoco的突变种nifB-Av1的钼铁蛋白组分做了晶体衍射结构分析, 发现4个亚单位中的1个构象发生了较大变化, 存在一个带正电的漏斗状(funnel)结构, 它足够容纳带负电的FeMoco的插入, 成为具有固氮功能的钼铁蛋白组分. 与此同时, 化学模拟固氮酶在温和条件下合成氨有了很大进展[41]. 在这个领域里我国也做了大量非常出色的工作: 固氮酶催化HD的形成绝对依赖于氮[42]; 在固氮酶催化还原N2的放氢机制中, 率先提出了双位点放H2模式, 对了解固氮酶催化机制有所发展[43]. 美国1992年用X光衍射确定固氮酶活性中心原子簇是由MoS3Fe和FeS3Fe3两个缺口的立方烷型簇合物组成[38], 通过3个非蛋白配体S桥联而成为一个笼(其顶端分别是Fe和Mo). 其实在此之前, 我国就已经合成了这两个簇合物[44]; 根据配位催化原理和化学探针思路, 提出活性中心原子簇笼应是活口的, N2还原成氨和质子还原成H2都是在笼内进行, 提出用于还原底物有两条质子通道的设想[45~47]. 这些进展对指导合成高效催化剂, 实现在温和条件下固氮有重要意义.1.4我国生物固氮研究成果的国际认可和曾经面临的困境“生物固氮”成为科学定义并开始大力研究已有114年的历史. 我国自1937年开始生物固氮研究, 已有65年历史. 20世纪70年代生物固氮研究在生物化学和分子遗传学等方面取得突破后, 我国也取得了一系列重要成果, 在国际上占有一定的地位, 在某些方面还具有重要影响. 因此, 国际生物固氮研究委员会主席W. Newton曾多次建议在中国召开国际生物固氮研究大会, 经研究决定2003年在北京召开第14届国际生物固氮大会.我国生物固氮研究的道路曲曲折折, 曾经有两种错误认识: 一是受到假冒伪劣生物固氮肥料的宣传的干扰, 认为生物固氮问题已经解决; 二是对国际和国内生物固氮研究的突破性进展了解不够, 认为难度大, 进展甚微, 国内经多年研究也未出成果. 两者的结果使我国的生物固氮研究面临严重困境.为防止困境再现, 经我国有关决策者和研究人员的共同努力, 恢复了固氮研究应有的地位. 这就为巩固研究成果, 继续发展, 不失时机地迎接生物固氮的重大突破的新时代的到来, 并把生物固氮研究中与生命科学其他学科相关的重大科学问题提高到一个新水平, 使其进一步为我国农业可持续发展做出重要贡献.2生物固氮研究的展望根据国际上生物固氮研究的发展和我国的具体情况, 我国生物固氮研究应着重下列几个方面.2.1联合(内生)固氮菌固氮基因调控及其在提供氮素中的作用联合或内生固氮菌大多数是自生固氮菌, 其固氮作用大小极易受外界环境因素(氧、铵、温度等)的制约. 在自然状态下有些固氮菌与植物有着松散的联合, 或进入植物成为内生菌, 这就为进一步利用这些固氮生物给植物提供氮素创造了更多的机会. 在研究固氮基因表达和调控的基础上, 有针对地进行固氮菌的遗传改造, 构建高效的固氮菌株, 以提高固氮效率, 减少化肥施用, 为作物提供更多的固氮量[13,14,20,21].2.2根瘤菌与豆科植物共生结瘤固氮的信号传递和分子相互作用豆科植物共生固氮由于其固氮作用的高效率, 始终是生物固氮研究的焦点之一. 根瘤菌与豆科植物之间的信号传递、相互识别、基因的顺序性表达和调节对根瘤的形成、发育和固氮作用的大小等有着错综复杂的联系. 苜蓿根瘤菌和苜蓿的共生结瘤固氮是共生固氮的模式系统, 研究最为集中, 但仍然有很长的路要走. 如果考虑到地区不同和自然环境的差异, 即使是同一模式系统也会有不同的差异表现, 更何况还有特异的共生固氮系统? 在分子水平上阐明两者相互作用的机理, 一方面旨在提高共生固氮的效率, 另一方面还可以为扩大根瘤菌宿主范围, 为实现粮食作物共生固氮提供理论依据和技术措施.2.3氮、碳代谢和固氮与光合作用的偶联氮、碳代谢是一切生物最基本的代谢作用, 而且是相互联系的. 固氮作用需要消耗作为能源的碳源. 植物共生固氮中固氮作用的能源直接来自光合作用. 固氮生物有选择性地利用碳源, 其中以四碳二羧酸糖的利用较好. 固氮生物在氮、碳代谢的基因表达中, 分别利用RNA聚合酶的σ54和σ70. 碳代谢调控蛋白CRP(carbon receptor protein)与结合在启动子上的σ54相互作用, 使依赖σ54的dctA和glnAp2等基因启动子的表达受到抑制[36,37], 其结果就在分子水平上将氮、碳代谢联系起来. 最近研究证明, CRP-cAMP同样抑制肺炎克氏杆菌nif基因的表达, 而且其抑制作用的大小与启动子上有无潜在的CRP结合位点直接相关[48,49].豆科植物是C3植物, 固氮作用需消耗光合作用能量的10%, 因此减产达5%, 固氮不增产[50]. 虽然在大豆根瘤菌中增加nifA正调控基因的拷贝数, 可以提高固氮作用, 增加产量[51,52], 但仍然需要植物提高光合作用效率, 才能满足既不施或少施氮肥, 又能达到增产的目的. 如何提高豆科植物的光合作用效率, 是光合作用和固氮作用的共同研究问题. 现有报道表明, 通过诱变获得高光效的大豆品种, 产量可提高30%左右[53]. 这一品种将为固氮和光合偶联研究提供了材料. 另一途径是通过转基因技术将C4植物的基因转入豆科植物, 使其变成C4植物. 这种可能性是存在的, 最近已经将玉米光合作用C4途径的基因转入水稻, 获得高光效增产幅度较大的转基因水稻[54~56], 这为获得C4豆科植物提供了借鉴.2.4共生固氮中包括蛋白质组学在内的功能基因组学研究共生固氮功能基因组学和蛋白质组学研究包括根瘤菌和宿主植物两个方面. 功能基因组学研究的前提是对目的生物的基因组进行全序列分析. 目前国际上已经对苜蓿根瘤菌基因组进行了全序列分析[57], 接着是大豆根瘤菌和百脉根根瘤菌(Rhizobium loti)基因组. 在宿主植物方面已经启动了对苜蓿Medicago sativa Lin)、大豆(Glycine max Lin)和百脉根(Lotus corniculatu)基因组序列的分析[58~60]. 这些研究成果将为固氮功能基因组和蛋白质组学研究奠定基础和建立技术平台. 目前, 固氮功能基因组和蛋白质组学已经陆续有所报道[61~63]. 固氮资源生物多样性研究表明, 不同根瘤菌可与同一豆科植物相互作用结瘤固氮, 但它们之间的结瘤固氮效率却大不相同. 同样, 同一根瘤菌可与不同属的豆科植物结瘤固氮[64]. 这一结果为开展共生固氮功能基因组学和蛋白质组学研究奠定了基础. 可以充分利用公布的苜蓿根瘤菌基因组序列, 通过RNA 和蛋白质差异显示法和微阵列法, 对不同苜蓿根瘤菌基因组及其突变株在共生条件下进行功能比较, 对不同根瘤菌在同一豆科植物结瘤的不同根瘤素基因表达进行比较, 将可大大推进共生结瘤固氮中微生物与植物相互作用机理的研究. 在此基础上, 还可寻找非豆科植物, 特别是禾本科植物中是否有以及有多少类似于豆科植物的根瘤素存在, 从而最终为非豆科植物的共生固氮和自主固氮提供策略和技术路线. 无疑, 共生固氮功能基因组和蛋白质组学研究将具有更为重大的科学意义和潜在的实际意义.致谢感谢沈善炯、李季伦和朱家壁教授对文稿的建议和修改及林敏、陈文新、周朝晖等教授所提供资料. 本工作为国家重点基础研究发展规划资助项目(批准号: 2001CB108904).参考文献1 Denarie J, Roche P. Rhizobium nodulation signals. In: Verma D PS. Molecular Signals in Plant-Microbe Communications. Boca Raton/Ann Arber/London: CRC Press, 1991. 296~3242 Pliverira A L M, Urquiaga S, Dobereiner J, et al. Biologicalnitrogen fixation (BNF) in micropropagated sugarcane plants inoculated with different endophytic diazotrophic bacteria. In: Pedrosa F O, Hungria M, Yates M G, eds. Nitrogen Fixation: From Molecules to Crop Productivity. Dordrecht/Boston/London: Kluwer Academic Publishers, 1999. 4253 Ribbe M, Gadkari D, Meyer O. N2 fixation by Streptomycesthermoautotrophicus involves a molybdenum-dinitrogenase and a manganese-superoxide oxidoreductanse that couple N2 reduction to the oxidation of superoxide produced from O2 by a molybdenum-CO dehydrogenase. J Biological Chemistry, 1997, 272: 26627~266334 Tan Z Y, Xu X T, Wang E T, et al. Phylogenic and geneticrelationoships of Mesorhizobium tianshanenese and related rhizobia. Int J Syst Bacteriol, 1997, 47: 874~8795 Wang E T, van Berkum P, Sui X H, et al. Diversity of rhizobiaassociated with Amorpha fruticosa isolated from Chinese soils and description of Mesorhizobium zmorpphae sp nov. Int J Syts Bacteriol, 1999, 49: 51~656 Tan Z Y, Wang E T, Peng G X, et al. Characterization of bacteriaisolated from wild legumes in the North-Western regions of China.Int J Syst Bacteriol, 1999, 49: 1457~14697 Yan A M, Wang E T, Kan F L, et al. Sinorhizobium melilotiiassociated with Medicago sativa and Melilotus spp. Int J Syst Bacteriol, 2000, 50: 1887~18918 Shen S C. Organization and regulation of nitrogen fixation genes:1974~1995. In: Kung S D, Yang S F, eds. Discoveries in Plant Biology. Vol Ⅲ. Dordrecht/Boston/London: World Scientific Press, 2000. 383~3929 朱家壁, 俞冠翘, 江群益, 等. 基因nifA产物对肺炎克氏杆菌(Klebsiella pneumoniae)gln突变型的Nif−表型的校正和固氮酶的组成型合成的作用. 中国科学, B辑, 1983, (8): 688~69610 Hu B, Zhu J B, Shen S C, et al. A promoter region binding proteinand DNA gyrase regulae anaerobic transcription of nifAL in Enterbacter cloacae. J Bacteriol, 2000, 182: 3920~392311 Wang Y P, Birkenhead K, Boesten B, et al. Genetic analysis andregulation of the Rhizobium melilotii genes controlling C4- dicarboxylic acid transport. Gene, 1989, 85: 135~14312 Bosworth A H, Williams M K, Albrecht K A, et al. Alfalfa yieldresponse to inoculation with recombinant strains of Rhizobiummelilotii with an extra copy of dctABD and/or modified nifAexpression. Appl Environ Microbiol, 1994, 60: 3815~383213 林敏, 尤崇杓, 刘永正, 等. 重组耐铵固氮菌株的田间长期定点释放试验. 生物技术学报, 1995, 1: 28~3314 李永兴, 李久蒂, 卢林刚, 等. 玉米联合固氮工程菌Enterobactergergivuae E7 在田间的接种效应. 中国农业科学, 2000, 33: 72~7715 Shen S C, wang S P, Yu G Q, et al. Expression of the nodulationand nitrogen fixation genes in Rhizobium melilotii during development. Genome, 1989, 31: 354~36016 王水平, 朱家璧, 俞冠翘, 等. 苜蓿根瘤菌(Rhizobium meliloti)nifA基因的异源表达及其产物的氧敏感性. 中国科学, B辑, 1990, (3): 261~26617 Deng X P, Shen S C. Structure and oxygen sensitivity of nifLApromoter of Enterobacter cloacae. Science in China, Ser B, 1995,38(1): 60~6618 赵洁平, 戴小密, 许玲, 等. 固氮正调节基因nifA促进大豆根瘤菌的结瘤效率. 科学通报, 2001, 46(23): 1984~198719 高云峰, 吴桐, 朱家璧, 等. 苜蓿根瘤菌固氮酶基因启动子P1转录起始点下游顺序(DS)的特性. 中国科学, C辑, 1996, 26(2): 100~10620 沈炳福. 水稻对耐铵工程固氮菌株的响应. 植物生理学报, 1995,21: 302~30621 张福星, 尤崇杓, 卢婉芳. 环境因子变化的水稻氮素吸收及接种效应的影响. 农业生物技术学报, 1995, 1: 93~9822 Hu B, Zhu J B, Shen S C, et al. A promoter region binding proteinand DNA gyrase regulae anaerobic transcription of nifAL in Enterbacter cloacae. J Bacteriol, 2000, 182: 3920~392323 Xiao H, Shen S C, Zhu J B. NifL, an antagonistic regulator ofNifA interacting with NifA. Science in China, Ser C, 1998, 41(3): 303~30824 何路红, 阎大来, 马旅雁. 肺炎克氏杆菌nifA基因在巴西固氮基因表达的铵调节中的作用. 生物工程学报, 1995, 11: 385~388 25 马旅雁, 吴奥, 赵银锁. 巴西固氮螺菌Yu62 dragTG基因及其下游区域的定位诱变. 生物工程技术学报, 1999, 15: 281~28726 马旅雁, 李季伦. 巴西固氮螺菌Yu62 dragTG基因启动子区域的核苷酸序列及其功能分析.生物工程学报, 1997, 13: 343~34927 朱冰, 戴小密, 朱家璧, 等. 苜蓿根瘤菌nod D3P1启动子下游序列的调节功能. 科学通报, 1999, 44(21): 2308~231228 Yu G Q, Zhu J B, Gu J, et al. Evidence that the nodulationregulatory gene nodD3 of Rhizobium melilot i is transcribed from two separate promoters. Science in China, Ser B, 1993, 36: 225~ 23629 吴桐, 朱家璧, 俞冠翘, 等. 苜蓿根瘤菌多拷贝固氮基因启动子对根瘤发育的抑制. 中国科学, B辑, 1994, 24(10): 1053~105930 陈迪, 刘彦杰, 朱家璧, 等. 苜蓿根瘤菌(Sinorhizobium meliloti)nodD3P1启动子下游序列的缺失和互补分析. 中国科学, C辑, 2002, 32(6): 512~51831 Wang L, Li C, Wang Q, et al. Chemical synthesis of NodRm-1:thenodulation factor involved in Rhizobium melilotii-legume symbiosis. J Chem Soc Perkin Trans, 1994, 1: 621~62832 ZhangJ X, Jing Y, Shen S H, et al. Transformation of twonitrogen-fixation-related plant genes into tobacco and their expressions. Acta Botanica Sinica, 2000, 42: 834~84033 Zhang J X, Wang Y P, Sheng S H, et al. Transformation of pealectin gene and Parasponia haemoglobin gene into rice and their expressions. Acta Botanica Sinica, 2001, 43: 267~27434 金润之, 江群益, 沈思师, 等. 紫云英根瘤菌nif DNA的分子克隆. 科学通报, 1992, 37(17): 1603~160635 金润之, 朱劲松, 江群益, 等. 紫云英根瘤菌Ra159的巨大质粒上存在有nod和nif基因的证明. 微生物学报, 1993, 33: 170~173 36 Wang Y P, Kolb A, Buck M, et al. CRP interacts withpromoter-bound σ54 RNA polymerase and blocks transcriptional activation of the dctA promoter. EMBO J, 1998, 17: 786~79637 Tian Z X, Li Q S, Buck M, et al. The CRP-cAMP complex anddownregulation of the glnAp2 promoter provides a novel regulatory linkage between carbon metabolism and nitrogen assimilation in E. coli. Mol Microbiol, 2001, 4: 911~92438 Kim J, Rees D C. Structural models for the metal centers in thenitrogenase molybdenum-ion protein. Science, 1992, 257: 1677~ 168239 Chan M K, Ress D C. The nitrogenase FeMo-cofactor andP-cluster pair: 0.22 nm resolution structure. Science, 1993, 260: 797~794 40 Schmid B, Ribbe M W, Einsle O, et al. Structure of acofactor-deficient nitrogenase MoFe protein. Science, 2002, 296: 352~35641 Nishibayashi Y, Iwai S, Hidai M. Bimetallic system for nitrogenfixation: ruthenium-assisted protonation of coordinated N2 on tungsten with N2. Science, 1998, 279: 540~54242 Li J, Burris R. Influence of pN2 and pH2 on HD formation byvarious nitrogenase. Biochemistry, 1983, 22: 4472~448043 张振挥, 吴柏和, 李季伦. 固氮酶催化的放H2反应. 微生物学报, 1993, 33: 320~33044 吴新涛, 卢嘉锡. 固氮酶活性中心网兜模型的回顾和前瞻. 科学通报, 1995, 40(7): 577~58145 Tsai K R, Wan H L. On the structure-function relationship ofnitrogenase M-cluster and P-cluster pairs. J Cluster Sci, 1995, 6: 485~50146 周朝晖, 颜文斌, 张凤章, 等. 固氮酶催化作用机理及其化学模拟, 厦门大学学报, 2001, 40: 320~32947 Zhou Z H, Yan W B, Wan H L, et al. Synthesis and characterization ofhomochiral polymeric S-malato molybdate (Ⅵ): toward the potentially stereospecific formation and absolute configuration of iron-molybdenum cofactor in nitrogenase. J Inorganic Biochem, 2002, 90: 137~14348 李稚婷, 孙义成, 毛贤军, 等. 碳代谢总体调控蛋白CRP对肺炎克氏杆菌启动子的抑制作用. 科学通报, 2002, 47(15): 1133~113949 李稚婷, 张维佳, 王忆平. 碳代谢总体调控蛋白CRP对nifA启动子的抑制作用不依赖于该启动子上游CRP与nifA竞争的靶位点. 科学通报, 2002, 47(16): 1242~124650 Bergersen F J. The Central reaction of nitrogen fixation. Plant andSoil, 1971, Special Vol: 511~52451 陈昌斌, 戴小迷, 俞冠翘, 等. 组成型nifA对根瘤菌(Rhizobiumfredii) HN01 lux结瘤效率的促进作用. 科学通报, 1999, 44(5):529~53352 Li Y, Zhou J C. Influence of introduced extra nifA gene onrhizosphere colonization and competition for nodule occupancy by Sinorhizobium fredii strain HN02 NL. J Huazhong Agrecultural University, 2000, 19: 198~20353 Hao N B, Du W G, Ge Q Y, et al, Progress in the breeding ofsoybean for high photosynthetic efficiency. Acta Botanica Sinica, 2002, 44: 253~25854 Ku M S B, Agarie S, Nomura M, et al. High-level expression ofmaize phosphoenolpyruvate carboxylase in transgenic rice plants.Nat Biotech, 1999, 17: 76~8055 焦德茂, 李霞, 黄雪清, 等. 转PEPC基因水稻的光合CO2同化和叶绿素荧光特性. 科学通报, 2001, 46(5): 411~41856 Huang X Q, Jiao D M, Chi W, et al. Characteristics of CO2exchange and chlorophyll fluorescence of transgenic rice with C4 genes. Acta Botanica Sinica, 2002, 44: 405~41257 Galibert F, Finan T M, Long S L, et al. The composite genome ofthe legume symbiont Sinorhizobium melilotii. Science, 2001, 293: 668~67258 Bell C J, Dixon R A, Farmer A D, et al. The Medicago genomeinitiative: A model legume database. Nucleic Acids Res, 2001, 29(1): 114~11759 Shoemaker R, Keim P, Vodkin L, et al. A compilation of soybeanESTs: generation and analysis. Genome, 2002, 45: 329~33860 Marek LF, Mudge J, Damielle L, et al. Soybean genomic survey:BAC-end sequences near RFLP and SSR markers. Genome, 2001, 44: 572~58161 Panter S, Thomson R, de Bruxelles G, et al. Identification withproteomics of novel proteins associated with the peribacteroid membrane of soybean root nodules. Molecular Plant-Microbe Interactions, 2000, 13: 325~33362 Mathesius U, Keijzers G, Natera S H, et al. Establishment of aroot proteome reference map for the model legume Medicago truncatula using the expressed sequence tag database for peptide mass fingerprinting. Proteomics, 2001, 1: 1424~144063 Natera S H, Guerreiro N, Djoefievic M A. Proteome analysis ofdifferentially displayed proteins as a tool for the investigation of symbiosis. Mol Plant-Microbe Interact, 2000, 13: 995~100964 张海瑜, 张海予, 李小红, 等. 一株能在苜蓿上结瘤的费氏中华根瘤菌. 微生物学报, 2001, 41: 129~132(2002-08-28收稿, 2002-11-28收修改稿)・动 态・ 第14届国际固氮大会将在北京召开 生物固氮与光合作用是自然界给予人类的两大贡献,它们分别提供了植物生长的氮源和碳源, 从而提供了人类社会发展的最基础的物质条件, 是农业生物学最基础的研究课题.国际固氮大会是生物固氮研究领域规模最大、规格最高、权威性最强的国际学术会议. 它每两年召开一次, 此前历届都由欧美各国主办, 本届大会是首次在中国召开,这也是亚洲首次取得举办权. 大会将于2003年11月1~6日在北京国际会议中心举行. 在国际固氮管理委员会的大力支持下, 囊括国际上本研究领域最著名专家学者(36人)的国际顾问委员会以及囊括国内本研究领域最著名专家学者(30人, 其中包括6位中国科学院院士)的国家委员会已经成立.大会预计将有1000名左右中外科学家参加, 将特邀本研究领域最著名的专家学者到会并做大会报告, 其中包括国际固氮管理委员会成员10名、国际固氮咨询委员会成员36名及其他大会特邀报告人40名左右.本届大会将包括4个全会和12个分会. 全会议题为: 1. 生物固氮基础研究; 2. 可持续农业与生物固氮的限制性因素; 3. 生物固氮研究前沿(新技术、基因组研究、固氮生物体系); 4. 根瘤菌-豆科植物共生体系. 分会议题为: 1. 固氮酶的生物化学及其化学模拟; 2. 固氮遗传与调控;3. 根际联合固氮及其内生固氮;4. 放线菌共生体系与林业;5. 光合固氮菌;6. 固氮微生物及其相关植物的基因组研究;7. 固氮微生物在可持续农业及其生物修复中的应用;8. 胁迫应答(耐盐)及其他生物固氮限制因素;9. 固氮体系的系统发育及其共生进化; 10和11. 根瘤菌-豆科植物共生体系; 12. 与发展中国家合作的生物固氮研究等当前生物固氮研究领域的各个方面.详细情况请参照网站提供的信息. 会议费用及要求等信息将在网站上公布.联系方式: 地 址: 北京大学生命科学学院(100871)联系人: 李凤梅王忆平电 话:010-6275 1006传 真: 010-6275 6325E-mail:mm307@。
植物氮素利用效率的研究进展氮素是植物生长中的必需元素之一,而植物的氮素利用效率会直接影响植物生长发育和农业生产。
因此,对于氮素利用效率的研究一直是植物生长领域的热点之一,今天我们就来谈一谈植物氮素利用效率的研究进展。
一、植物对于氮素的吸收和利用植物对于氮素需要通过根系吸收,一般来说,被吸收的氮素有两种形式:一种是无机氮素,如硝酸盐(NO3-)和铵盐(NH4+),另一种是有机氮素,如氨基酸和蛋白质。
吸收后的氮素将会进入植物体内进行利用。
植物内部的氮素转运过程主要是由nrt1.1、nrt2.1、nrt2.2等基因编码的氮素转运蛋白进行调节。
二、植物氮素利用效率的调控机制植物的氮素利用效率受到多种因素的影响,在这其中,植物生理性状对氮素吸收、转运和利用的影响是最为重要的。
近来,一些研究表明,调节植物氮素利用效率的物质主要包括:激素、信号传递分子、转录因子等。
1. 激素调节激素的调节作用对于植物的氮素利用效率非常重要。
比如,生长素和赤霉素可以促进植物对氮素的吸收,而赤霉素还会作用于NRT1.1和NRT2.1等基因,从而促进植物对氮素的转运。
2. 信号分子调节在植物体内,一些信号分子也能够调节植物对氮素的吸收和利用。
例如,在有机酸泵抑制素(OGDs)和一氧化氮(NO)的作用下,植物根部会释放出有机酸和NO分子,从而提高植物对于氮素的吸收能力。
3. 转录因子调节转录因子是指能够影响植物基因表达的蛋白质家族。
在植物对于氮素的利用过程中,转录因子也会扮演一个重要的角色。
例如,植物体内的MYB、WRKY和NAC等转录因子家族均能够协调植物自身对于氮素的吸收和利用,从而提高植物的氮素利用效率。
三、植物氮素利用效率的提高途径在应对氮素资源缺乏等情况下,提高植物的氮素利用效率也是农业生产过程中一个非常重要的问题。
那么,有哪些途径能够提高植物的氮素利用效率呢?1. 穴盘法耕作穴盘法耕作又称作净化机耕作,该法耕作具有良好的保水性和保肥性,相比于传统溅水法耕作,穴盘法耕作不仅能够降低氮素和钾素的损失率,同时也能够提高氮素利用效率。
利用SPAD值进行作物氮营养诊断及追肥量的确定当作物在缺氮时一般会表现出叶片叶绿素含量降低导致的叶色变浅,而氮素过多,叶色颜色变深等一些明显的症状。
因此,可以通过植物叶片含氮量与叶片叶绿素含量密切相关性及变化相似性来监测植物的氮素状况。
而作物叶片光反射特性与叶色深浅存在定量关系,并且植物在可见光波段的反射率主要受叶绿素的影响,根据这一原理日本已经开发了手持式叶绿素计(SPAD502)来进行作物氮营养诊断及指导施肥,并且在大田作物如玉米、小麦、大豆及水稻上均有应用。
本研究通过田间试验与室内生化测定相结合的方式,田间测定玉米水稻叶片SPAD值,实验室测定叶片氮含量及拷种,根据试验与调查结果,明确在不同条件下SPAD值与玉米、水稻氮素营养间的关系,以便用户可根据实际栽培条件,确定合适的追肥量和追肥时间,同时为以后进一步推广应用提供技术支撑。
主要结论如下:(1)随着施氮量的增加水稻产量呈现先增加而后趋于稳定的趋势,呈现出二次函数曲线,经计算得到当施氮量为170kg N/ha时产量最高能达到7078kg/ha。
(2)水稻不同SPAD值条件下追肥效果不同,但是SPAD值动态监测趋势基本一致,施氮量较低时穗肥施用后效果明显好于施氮量较高时的效果。
(3)水稻叶片SPAD值出现40、38时3天之内追施氮肥最优,而最迟不能超过15天,同时测定日期应该控制在扬花期以前。
(4)当水稻叶片SPAD值为40,总体施氮量为180kg N/ha,分为基肥(占总施氮量的40%)、拔节肥(占总施氮量的30%)和穗肥(占总施氮量的30%)3次分别施入时水稻氮素利用率最高。
(5)随着施氮量的增加玉米叶片SPAD值明显升高,在整个监测期间玉米叶片SPAD值有一个明显的升高阶段,后期玉米叶片SPAD值基本上趋于平衡状态。
(6)玉米叶片叶基部到叶尖不同位置SPAD值符合二次函数曲线变化,从叶基部开始40%~70%的区域测定值变异最小,平均测定值为58.5。