XXXX-XXXX年度吉林省农业技术推广奖立项项目名单(1)
- 格式:doc
- 大小:103.50 KB
- 文档页数:2

吉林省科技厅关于授予2014年吉林省科学技术奖的决定【法规类别】机关工作综合规定【发布部门】吉林省科学技术厅【发布日期】2014.11.04【实施日期】2014.11.04【时效性】现行有效【效力级别】XP10吉林省科技厅关于授予2014年吉林省科学技术奖的决定各市、州科技局、长白山管委会社会科技管理部门、省直有关厅(局)、高等院校、科研单位、企业:经省政府批准,决定授予《多功能稀土发光材料的控制合成及在显示与生物医学领域的应用基础》等10个项目吉林省自然科学奖一等奖;授予《高粱、玉米自交系和杂交种的表观遗传变异及其与杂种优势关系研究》等16个项目吉林省自然科学奖二等奖;授予《振荡流热管自激强化传热特性实验研究与理论分析》等15个项目吉林省自然科学奖三等奖。
授予《大气信道无线激光通信技术及应用研究》等4个项目吉林省技术发明奖一等奖;授予《镁合金表面处理环保新技术的开发与应用》等6个项目吉林省技术发明奖二等奖;授予《大型、精密多工位模具研发应用项目》等14个项目吉林省技术发明奖三等奖。
授予《空间快速响应高分辨率成像技术》等19个项目吉林省科学技术进步奖一等奖;授予《采暖期间大型供热机组与风力发电联合调峰优化运行研究与应用》等84个项目吉林省科学技术进步奖二等奖;授予《中药大品种银花泌炎灵片成果转化关键技术研究》等118个项目吉林省科学技术进步奖三等奖。
授予德国奥尔登堡大学的范思哲教授和韩国建国大学的全炳台教授吉林省国际科学技术合作奖。
希望全省科技工作者向获奖人员学习,继续发扬刻苦钻研、开拓创新精神,深入实施“创新驱动”战略,奋力攀登科技新高峰,为我省经济发展和社会进步做出更大贡献。
附件:2014年吉林省科学技术奖获奖项目名单吉林省科学技术厅2014年11月4日附件:2014年吉林省科学技术奖获奖项目名单省自然科学奖一等奖(共10项)序号项目名称主要完成人主要完成单位1多功能稀土发光材料的控制合成及在显示与生物医学领域的应用基础1林君,2李春霞,3程子泳,4杨飘萍,5马平安,6侯智尧,7李国岗,8代云路,9尚蒙蒙,10杨冬梅1中国科学院长春应用化学研究所2稀土单分子磁体弛豫机理与调控1唐金魁,2张洪杰,3郭云南,4林双燕,5张鹏,6赵朗1中国科学院长春应用化学研究所3人参活性多糖的系统研究1周义发,2台桂花,3程海荣,4高娟,5范玉莹,6崔思思1东北师范大学4基于基因组条形码的重大疾病诊断靶标泛1李凡,2王国庆,3倪朝辉,4王放,5刘彬,6高航,7杨青,1吉林大学基因组偶联的系统研究8黄红兰,9王槐栋,10张晓天5相干光学介质的动态量子操控和可逆信息存储1吴金辉,2高锦岳,3王海华,4崔淬砺,5魏小刚,6王春亮,7崔海宁,8康智慧,9李爱军,10姜云1吉林大学6吉林西部燕麦种植模式、水肥生理及加工利用技术理论基础研究1任长忠,2曾昭海,3胡跃高,4胡新中,5李再贵,6郭来春,7赵桂琴,8沙莉,9魏黎明,10王春龙1吉林省白城农业科学院,2中国农业大学,3陕西师范大学,4甘肃农业大学7恶性疟原虫、日本血吸虫免疫逃避和致病的机理研究1陈启军,2姜宁,3尹继刚,4陆慧君,5王心蕊,6常巧呈,7杜承,8张岩,9武闯1吉林大学8沼泽湿地碳循环及其对气候变化和人类活动的响应机理1宋长春,2张金波,3王毅勇,4汤洁,5孙丽,6王丽丽,7宋艳宇1中国科学院东北地理与农业生态研究所,2吉林大学9共轭高分子复合薄膜形态调控与性能1杨小牛,2鲁广昊,3黎立桂,4唐浩为,5赵晓礼1中国科学院长春应用化学研究所10有机聚合物光电功能材料的基础研究1田文晶,2徐斌,3温善鹏,4刘雷静,5张继博,6郭庆1吉林大学二等奖(共16项)序号项目名称主要完成人主要完成单位1高粱、玉米自交系和杂交种的表观遗传变异及其与杂种优势关系研究1张美善,2刘宝,3赵欣欣,4徐春明,5于晓明,6赵娜,7王欢,8杨巍1吉林农业大学,2东北师范大学2人源表观遗传调控酶的发现以及其在肿瘤发病中的作用1金景姬,2蔡勇,3王勇,4孙文涛,5王飞,6倪劲松1吉林大学3磺化聚芳醚类质子交换膜的结构构筑及传导特性调控1王哲,2那辉,3倪宏哲,4赵成吉,5张会轩,6张明耀,7徐晶美,8程海龙1长春工业大学,2吉林大学4分数阶微积分控制理论及系统鲁棒性增强策略研究1王春阳,2白端元,3李明秋,4田成军,5姜淑华,6陈宇,7刘妍妍,8詹伟达1长春理工大学5钙调蛋白磷酸酶、钙调素与tau蛋白相互1于大禹,2魏群,3骆静,4乔楠,5佟丽1东北电力大学,2北京师范大学作用分子机制的研究6ZnO基纳米材料Ca2+电化学生物传感器的研制及其性能研究1郎集会,2韩强,3高铭,4张旗,5刘晓艳,6杨景海,7杨丽丽,8曹健1吉林师范大学,2江苏大学7复杂知识处理的基本方法研究1刘大有,2杨博,3王生生,4欧阳继红,5虞强源,6贾海洋,7朱允刚,8李丽娜1吉林大学8新型介电陶瓷材料结构的微观设计、缺陷评价与高介电性能1路大勇,2孙秀云,3王严东,4韩丹丹,5刘巧丽,6岳洋,7张丽,8崔淑珍1吉林化工学院9不确定性结构动力学、智能结构控制及结构拓扑修改动态重分析1陈塑寰,2陈宇东,3姚国凤,4刘昕晖,5曹宗杰,6王忠东,7宋敏1吉林大学10炎性信号的生物学作用及炎症相关的信号转导机制研究1巴雪青,2霍洪亮,3赵岩,4曾宪录,5薛嬿,6刘琳琳,7可月双1东北师范大学,2吉林省疾病预防控制中心11关于植物应对环境变化的生活史策略研究1赵骥民,2张彦文,3杨春峰,4王星1长春师范大学12雌激素受体ERα和ERβ调控基因表达的制约机制1刘雅文,2赵春艳,3程熠,4李勇,5吴兴波,6魏卓,7官鑫,8翁熹君1吉林大学13中药生物碱CE-ECL分离与检测增强机制研究1高英,2杨晓东,3向前,4吴仙花,5李敬,6许元红,7王淑萍,8王琨琦1长春工程学院14常用中药材中治疗糖尿病等症的活性成分分离筛选及结构表征研究1刘春明,2张语迟,3刘舒,4郑梅竹,5王晶,6刘志强,7时东方,8皮子凤1长春师范大学,2中国科学院长春应用化学研究所15H6PD及11β-HSD1相互调节在2型糖尿病发病机制中的作用1陈立,2张明,3杜红伟,4李晶,5王春艳,6吕晓艳1吉林大学基础医学院16脑缺血再灌注性过程中蛋白聚集及缺血后处理的脑保护作用机制研究1葛鹏飞,2梁建民,3罗毅男,4王海峰,5罗天飞,6付双林,7冯春生,8陈大伟1吉林大学三等奖(共15项)序项目名称主要完成人主要完成单位号1振荡流热管自激强化传热特性实验研究与理论分析1商福民,2刘登瀛,3刘建红,4冼海珍,5韦节廷1长春工程学院2生物可降解载体材料的设计与应用1陈莉,2邓明虓,3丁建勋,4张喆,5孙敬茹1东北师范大学,2中科院长春应用化学研究所3反射地震Kirchhoff型有限孔径成像的数学理论与实现技术1孙建国,2孙章庆,3韩复兴1吉林大学地球探测科学与技术学院4新型复合纳米颗粒的制备及生物检测应用1刘丽炜,2谭勇,3张喜和,4胡思怡,5冯悦姝1长春理工大学5新型抗菌化合物的设计、合成与活性研究1朴虎日,2郑昌吉,3宋明霞,4孙良鹏,5陈振华1延边大学6CoPt纳米结构制备及磁学性能演化的调控1张永军,2王雅新,3刘洋,4李佳,5翟宏菊1吉林师范大学7基于本体的文本挖掘方法研究1孙铁利,2杨凤芹,3吴迪,4孙红光,5杨喜权1东北师范大学8基于光子和原子的量子计算与量子纠缠网络1王洪福,2张寿1延边大学9土壤动物对受损生态系统的指示及修复作用研究1董炜华,2殷秀琴,3魏健,4辛树权,5褚丽娟1长春师范大学,2东北师范大学10植物适应碱化环境的生物学响应机制系列研究1郭立泉,2麻莹,3石德成,4阚君满,5马传福1吉林工商学院11红色荧光转基因五指山小型猪的克隆1尹熙俊,2崔成哲,3康锦丹,4李所,5秦炜赜1延边大学,2延边朝鲜族自治州农业科学院(延边特产研究所)12重大心脏疾病心肌重塑的药物作用与细胞内信号转导通路的研究1王艳春,2张大维,3孙红霞,4任旷,5王立英1吉林医药学院,2北华大学,3吉林大学13辐射诱导免疫细胞间反应机制及其在肿瘤基因-放射治疗中的应用1金顺子,2孙世龙,3武宁,4张海玉,5杨建征1吉林大学14不同应激条件诱导肿瘤细胞内质网应激-自噬反应作用机制的研究1徐冶,2曹慧玲,3李质馨,4于洋,5刘师兵1吉林医药学院15胃肠激素调节饮食诱导肥胖能量摄入的机制1王舒然,2李杰,3张娜,4赵丹,5麻微微1吉林医药学院省技术发明奖一等奖(共4项)序号项目名称主要完成人主要完成单位1大气信道无线激光通信技术及应用研究1姜会林,2宋路,3张立中,4佟首峰,5李洪祚,6高天元,7朱一峰,8于林韬,9唐雁峰,10王彤宇,11刘岩,12杨絮,13胡源,14孟立新,15陈桂芬1长春理工大学2用于合成橡胶新型高效稀土催化体系的研发及产业化应用1张学全,2白晨曦,3姜连升,4代全权,5毕吉福,6于琦周,7张春雨,8张贺新,9李柏林,10那丽华,11蔡洪光,12柳希春,13李继文,14赵丽萍1中国科学院长春应用化学研究所3氩氧精炼铁合金工艺及其测控技术1尤文,2张德江,3曹志强,4邵安林,5林晓梅,6郭军,7韩顺杰,8马海涛,9谢慕君,10吴化,11王淮,12卢秀和,13张袅娜,14候云海,15王盛慧1长春工业大学,2中钢集团吉林铁合金股份有限公司,3鞍钢集团矿业公司4旋毛虫病早期诊断抗原基因及其独特表观遗传学调控机制的发现1刘明远,2吴秀萍,3付宝权,4王学林,5刘晓雷,6白雪,7黎诚耀,8高飞,9卢强,10李扬1吉林大学,2中国疾病预防控制中心寄生虫病预防控制所,3中国农业科学院兰州兽医研究所,4南方医科大学,5深圳华大基因二等奖(共6项)序号项目名称主要完成人主要完成单位1镁合金表面处理环保新技术的开发与应用1常立民,2段小月,3刘丹,4刘伟,5史凯,6孙小,7徐佳琦,8时杰丽1吉林师范大学2软件系统建模、验证及支撑平台关键技术1刘淑芬,2王晓燕,3李兵,4包铁,5姚志林,6曲明,7张欣1吉林大学,2长春吉大斯博莱科技有限责任公司及应用佳,8吴姚睿,9彭君,10韩璐,11庞世春,12兰庆国,13郑万波3六自由度轮胎试验机1郭孔辉,2杨杰,3董立波,4王英麟,5郭川,6吴平,7施光涛,8李忠岩1长春孔辉汽车科技有限公司4草莓花瓣诱导突变体培育新品种和高效脱毒技术1顾地周,2朱俊义,3姜云天,4陆爽,5冯颖,6夏广清1通化师范学院5脲酶阴性、无豆腥味、可直接食用的大豆分离蛋白生产方法1李荣和,2张雁南,3高长城,4李丹,5吴淑清,6牟宗毅,7李玉馨,8刘辉,9刘雷,10雷海容,11吴修利,12许伟良,13梁洪祥1长春大学,2通榆县益发合大豆制品有限责任公司6氧化樟脑的合成工艺及产业化研究1杜培革,2关大伟,3姜爽,4安丽萍,5苑广信,6翁双凤,7李洪宇,8孙彧峰,9赵南晰,10李坦城1北华大学,2长春大政药业科技有限公司三等奖(共14项)序号项目名称主要完成人主要完成单位1大型、精密多工位模具研发应用项目1王中,2刘继彦,3赵晓峰,4袁野,5李玉鹏,6张磊,7黄文胜,8金波1吉林省元隆达工装设备有限公司2一种水位计的自冲洗方法及装置1兰凤友,2孙立东,3王井华,4于兴萍1吉林省隆华测控设备制造有限公司3微纳光通信器件与系统1梁静秋,2王维彪,3梁中翥,4秦余欣,5田超,6崔乃迪,7吕金光,8陈松1中国科学院长春光学精密机械与物理研究所4一种低碳低硅铝镇静钢的制备工艺1王磊,2陈景彦,3林瑞民,4孙志明,5朱学刚1吉林建龙钢铁有限责任公司55-20kW偏低风速高可靠离网型风力发电系统1张雪明,2徐明奇,3郭景富,4朱挽强,5董永军,6陈健梅,7吴百公,8赵阳1东北师范大学6多点控制三维拉弯扭转成形模具1于沛洲1吉林省洲海科技有限公司7抗起球腈纶纤维1邵宝忠,2李子,3张海鸥,4杨雪峰,5刘明哲,6杨标,7郝1吉林奇峰化纤股份有限公司朋林,8杨立杰8开利纯电动汽车核心零部件研究开发1王东晨,2吴畏,3拱印生,4魏强,5姜鹏,6黄敏,7朱庆林,8丁勇1启明信息技术股份有限公司,2一汽-大众汽车有限公司9大豆食心虫干扰驱避剂的研究与应用1徐伟,2付晓霞,3臧连生,4史树森,5袁海滨,6田径,7毕锐,8崔娟1吉林农业大学10树脂基牙科修复材料的研究与产业化1闫鹏涛,2马荣堂,3慕琪,4王继英,5谢起俭1吉林省登泰克牙科材料有限公司11专利技术在启脾口服液的生产及质量控制中的应用1丁爱英,2徐阳,3崔业波,4樊美玲,5张晓峰,6罗敏,7马晓静,8李艳茹1吉林益民堂制药有限公司12胃秘镁颗粒工艺研究1杨尚华,2郭智华,3王丽玲,4薛芬,5刘洋,6毕英南,7刘启鹏,8常永亮1弘美制药(中国)有限公司13单唾液酸神经节苷脂钠注射液产业化1芦志刚,2熊峰,3韩荔,4王化宇,5孙伟强,6李洪岩,7李弈辛,8唐浩1吉林英联生物制药股份有限公司,2磐石妇幼保健院14一种治疗急性肠炎的中药分散片及其制备方法和应用1解钧秀,2王永宽,3于江波,4曲波,5王永彬,6杜凤娟,7张秀云,8祖双1吉林敖东延边药业股份有限公司省科学技术进步奖一等奖(共19项,其中包括科技成果转化贡献奖1项,序号1)序号项目名称主要完成人主要完成单位1350km/h高速动车组1赵明花,2李军,3梁树林,4常振臣,5于洪波,6王锋,7王金田,8何广忠,9周文平,10刘长青,11朱彦,12王树宾,13沙淼,14孔风,15邓海1长春轨道客车股份有限公司2甲状腺癌诊疗体系的创新及其关键技术的应用1孙辉,2马庆杰,3王辉,4王丽萍,5刘晓莉,6张广,7张大奇,8周乐,9付言涛,10边学海,11高识,12王清,13张文杰,14赵劼,15孙文伟1吉林大学3超高产耐盐碱水稻新品种东稻4的育成与推广1杨福,2梁正伟,3王志春,4李彦利,5李景鹏,6李景宏,7金哲宇,8马红媛,9黄立华,101中国科学院东北地理与农业生态研究所王明明,11杨帆,12杨昊谕,13刘淼,14安丰华,15张艳清4空间快速响应高分辨率成像技术1张学军,2李志来,3鲍赫,4薛栋林,5郑立功,6曲宏松,7高劲松,8吴清文,9董得义,10樊延超,11魏君成,12徐伟,13柴方茂,14杨会生,15曲利新1中国科学院长春光学精密机械与物理研究所5大规模风电基地功率集中外送输电容量优化规划方法及其工程应用1穆钢,2严干贵,3陈兴良,4肖白,5聂昕,6崔杨,7高晓峰,8杨林,9杨修宇,10李军徽1东北电力大学,2国网吉林省电力有限公司6轨道车辆整车及系统参数测定优化关键技术开发1王金田,2苏建,3谭富星,4滕万秀,5周殿买,6吕义,7刘玉梅,8刘洪涛,9程冰,10徐连萍,11程亚军,12陈熔,13林慧英,14王伟,15高阳1长春轨道客车股份有限公司7紧临既有线承压富水砂土互层地铁暗挖关键技术研究1曲建生,2姜永军,3宋云财,4曹伍富,5徐润泽,6梅源,7王新强,8赵元根,9徐俊峰,10贾少春,11张立华,12汪本刚,13王晶龙,14戴文革,15黎龙1中铁现代勘察设计院有限公司,2西安建筑科技大学,3北京市轨道交通建设管理有限公司,4中铁十三局集团有限公司第二工程公司,5中国铁建十三局集团有限公司8利用再制造技术开发防腐防垢金属陶瓷内衬油管1徐滨士,2杨永利,3石亮亮,4戚东涛,5魏世丞,6李厚补,7于宏超,8李永明,9付文耀,10田启武,11李俊成,12王守泽,13孙世杰,14刘化天,15韩飞1松原大多油田配套产业有限公司,2中国人民解放军装甲兵工程学院,3中国石油集团石油管工程技术研究院9数控机床可靠性综合设计与试验技术1杨兆军,2申桂香,3陈菲,4李国发,5许彬彬,6郝庆波,7贾亚洲,8陈传海,9朱晓翠,10张英芝,11呼烨,12张新戈,13杨永海,14赵宏伟,15贾庆祥1吉林大学10吉林省粮食安全储藏体系结构分析及新型产地储藏装备的开发与应用1吴文福,2孟凡刚,3张亚秋,4韩峰,5王昕,6徐岩,7张立辉,8陈思羽,9肖英奎,10刘春山,11薛恩儒,12刘哲,13王桂英,14吴玉柱,15秦骁1吉林大学,2吉林省粮油科学设计研究院,3长春吉大科学仪器设备有限公司11玉米绿色供应链关键技术研究与产业化应用1刘景圣,2张大力,3郑明珠,4闵伟红,5孙永海,6贾洪雷,7蔡丹,8王玉华,9张贵林,10刘回民,11许秀颖,12修琳,13詹冬玲,14杨青山,15刘惠麟1吉林农业大学,2吉林大学,3吉林天景食品有限公司,4吉林佳粮玉米食品有限公司12水貂、蓝狐精准营养研究与饲料高效利用技术1杨福合,2高秀华,3张铁涛,4耿业业,5杨颖,6任二军,7李光玉,8邢秀梅,9张海华,10王雷,11刘汇涛,12吴学壮,13蒋清奎,14孙伟丽,15刘志1中国农业科学院特产研究所,2中国农业科学院饲料研究所,3北华大学,4石家庄市农林科学研究院13植物重要基因资源挖掘与大豆特异种质创制及新品种选育1李海燕,2王振民,3董园园,4王法微,5王南,6李晓薇,7陈欢,8崔喜艳,9陈喜凤,10杨巍,11刘伟灿,12康波,13邓少华,14周永刚,15邓宇1吉林农业大学14赤眼蜂高效利用与生产关键技术研究及其大面积推广应用1阮长春,2孙光芝,3臧连生,4张俊杰,5朱兴友,6陈立玲,7邵玺文,8李天昊,9杜文梅,10王秀梅,11刘志,12刘显娇,13白庆荣,14金峰,15方丽1吉林农业大学15高清晰高均匀度全色LED大屏幕显示器1王瑞光,2郑喜凤,3陈宇,4黄凌碧,5邸楠,6罗锦,7邢建枫,8肖传武,9甘春和,10汪洋,11张鑫,12邓意成,13苗静,14刘双双,15马新峰1长春希达电子技术有限公司,2中国科学院长。

吉林省农业委员会关于公布吉林省休闲农业与乡村旅游星级示范企业的通知文章属性•【制定机关】吉林省农业委员会•【公布日期】2017.08.22•【字号】•【施行日期】2017.08.01•【效力等级】地方规范性文件•【时效性】失效•【主题分类】村镇建设正文吉林省农业委员会关于公布吉林省休闲农业与乡村旅游星级示范企业的通知各市(州)农委,长白山管委会环资局,公主岭、梅河口市农业局,各县(市、区)农业局:为了进一步推动全省休闲农业与乡村旅游快速发展,打造休闲农业品牌,创建休闲农业与乡村旅游星级示范企业,促进农村一二三产业融合发展。
省农委印发了《关于开展2017年全省休闲农业与乡村旅游星级示范创建工作的通知》(吉农办加[2017]6号文件)。
在各地农业行政主管部门推荐的基础上,省农委组织考核小组,按照评分标准及现场考查,最后确定104家全省休闲农业与乡村旅游星级示范企业,其中,五星级31家,四星级36家,三星级37家,现予以公布。
星级示范企业实行动态管理,有效期从2017年8月-2020年7月。
希望被评定的休闲农业与乡村旅游星级示范企业,加强自我管理、自我规范、自主提升,进一步提高发展水平,发挥休闲农业与乡村旅游星级企业示范引领带动作用,转变农村发展方式,调整农村产业结构,为推动全省农村经济社会协调发展做出更大贡献。
附件:吉林省休闲农业与乡村旅游星级示范企业名单吉林省农业委员会2017年8月22日附件吉林省休闲农业与乡村旅游星级示范企业名单五星级1、吉林省荣发生态农业开发有限公司2、吉林省缘山湖农业园有限公司3、农安县春江堰家庭农场4、吉林省玉林湾旅游开发有限公司5、长春奢爱农业科技发展有限公司6、长春宏禹生态园餐饮有限责任公司7、吉林省智成农业科技有限责任公司8、农安县顺民心农牧合作社9、榆树市明月生态园有限公司10、九台区碧水庄园度假有限公司11、德惠市洪瑞德休闲农业游专业合作社12、长春市红奇休闲度假中心13、长春北城水上乐园有限公司14、农安县三角湖现代农业发展有限责任公司15、吉林省森特旅游开发有限公司16、吉林市春新生态家庭农场17、吉林省天鼎旅游产业发展有限责任公司18、延吉小桥流水人家餐饮有限公司19、吉林省北方巴厘岛游乐有限公司20、通化县转水湖农业开发有限公司21、集安豆谷离宫大酒店有限公司22、通化永生神龙商务有限公司23、通化市金江花海游览有限公司24、集安太极湾旅游管理有限公司25、大安牛心套保旅游产业发展有限公司26、通榆向海乡休闲旅游农业产业园区27、吉林省鴜鹭湖生态旅游度假发展有限公司28、白山中天农业科技发展有限公司29、长白朝鲜族自治县紫罗兰旅游文化有限公司30、松原兴源种植农民专业合作社31、松原市惠益丰农业开发有限公司四星级1、长春绿屋家庭农场2、吉林省金德瑞农业科技开发有限公司3、长春市富仁潭游乐中心4、榆树市都市休闲家庭农场5、长春向阳源生态农业有限公司6、九台市东师物业有限公司7、长春市兴晟家家乐旅游有限公司8、长春龙福田园观光旅游有限公司9、吉林省彬生农业科技发展(集团)有限公司10、长春德胜宝农业有限公司11、桦甸市红石镇如意休闲娱乐山庄12、吉林市红原源家庭农场13、吉林市黑屯旅游度假有限公司14、吉林市龙潭区清泉休闲山庄15、磐石市神宇休闲渔业有限公司16、磐石市烟筒山镇金雨湖养殖园17、蛟河市文化山庄18、图们市百年部落民俗文化旅游有限公司19、敦化市雁鸣小镇休闲农业旅游度假村20、延吉市世外桃源开发有限公司21、四平市铁西区赵家大院22、伊通满族自治县铠绎农牧业发展有限公司23、伊通满族自治县德盛农牧科技发展有限公司24、吉林省广吉行文创乡村旅游开发有限公司25、集安市鸭江谷酒庄有限公司26、通化市大明乳业有限责任公司27、通榆县天意农产品经贸有限责任公司28、白城市升华农林有限公司29、镇赉县建平乡金丰旅游度假山庄30、临江市珍珠旅游开发有限公司31、临江市生记种植养殖农民专业合作社32、白山市江源区圣达农业发展有限公司33、靖宇县蓝缇蓝莓种植专业合作社34、吉林省吉松食品有限公司35、长白山瑞仙双莓景区36、公主岭市二十家子明亨旅游度假村有限公司三星级1、吉林天安农业开发有限公司2、九台区平安堡乡村旅游合作社3、长春市盛泽农业有限公司4、永吉麒麟山滑雪场有限公司5、磐石市烟筒山镇鹿鸣山庄6、吉林过亿休闲农业旅游度假村有限公司7、和龙市青山里土特产经销专业合作社8、吉林省多帮生态农业有限公司9、通化县韵欢文化艺术传播有限公司10、通化鱼乐源餐饮有限公司11、通化县东来乡鹿圈子关东民俗村12、集安市永泉民俗旅游开发有限责任公司13、通化市园林休闲农业科技有限公司14、白城市丰谷农业产业发展有限责任公司15、大安市裕丰粮贸有限公司16、大安市久华农业有限公司17、大安市农业科技示范场18、洮南市林桥生态旅游观光休闲有限公司19、吉林省盛保乐休闲农庄有限公司20、吉林省东合农业股份有限公司21、抚松县黄家崴子果蔬种植专业合作社22、抚松县松江河白家渔村养殖有限公司23、抚松县兴隆龙鲤旅游服务开发有限公司24、长白朝鲜族自治县有珍生态园25、长白朝鲜族自治县金斗笠高丽食品贸易有限公司26、长白朝鲜族自治县石门湖旅游有限公司27、长白朝鲜族自治县吴老五家庭农场28、长白朝鲜族自治县南川村玉光棚膜经济种植合作社29、长白朝鲜族自治县景田山庄30、靖宇县碧水川生态园有限公司31、靖宇县三合苑科学技术推广专业合作社32、白山市飞宇旅游开发有限公司33、临江市岭上枫林林业牧产业有限责任公司34、临江市五河山庄35、吉林省亿荣农业开发有限公司36、梅河口市宝金果蔬种植专业合作社37、双辽一马树森林公园。

㊀山东农业科学㊀2023ꎬ55(9):87~93ShandongAgriculturalSciences㊀DOI:10.14083/j.issn.1001-4942.2023.09.012收稿日期:2022-11-21ꎻ修回日期:2023-05-25基金项目:2019年度吉林省科研院所引进高层次科技创新人才资助计划项目ꎻ吉林省农业科技创新工程基本科研经费项目(KYJF2021JQ103)作者简介:瞿子惠(1995 )ꎬ女ꎬ硕士ꎬ研究实习员ꎬ从事动物营养与饲料研究ꎮE-mail:47992301@qq.com通信作者:郎洪彦(1973 )ꎬ女ꎬ硕士ꎬ副研究员ꎬ从事动物科学研究ꎮE-mail:bluewater603@163.com陈龙(1989 )ꎬ男ꎬ博士ꎬ副研究员ꎬ从事动物营养与饲料科学研究ꎮE-mail:chenliang198931@163.com贝莱斯芽孢杆菌CL-4固态发酵对豆粕营养品质的影响瞿子惠ꎬ刘歆ꎬ郑琳ꎬ魏炳栋ꎬ闫晓刚ꎬ于维ꎬ陈龙ꎬ郎洪彦(吉林省农业科学院动物营养与饲料研究所ꎬ吉林公主岭136100)㊀㊀摘要:本试验选用吉林省农业科学院动物营养与饲料研究所分离鉴定的贝莱斯芽孢杆菌CL-4对豆粕进行固态发酵ꎬ通过对发酵前后豆粕中营养成分㊁大豆抗原蛋白㊁酶活力㊁活菌数㊁抗菌活性及表观形态等指标的测定ꎬ评价贝莱斯芽孢杆菌CL-4固态发酵豆粕营养品质的提升效果ꎮ结果表明:贝莱斯芽孢杆菌CL-4在大豆抗原蛋白筛选平板上显示出较大直径的水解圈ꎬ具有降解大豆抗原蛋白的能力ꎮ固态发酵24h显著提高了豆粕营养品质和功能代谢产物ꎬ具有更高浓度的酸溶蛋白㊁钙㊁灰分和总磷含量ꎬ其中粗蛋白含量由46.78%增加到51.28%ꎬ总氨基酸含量由41.72%显著提高至48.14%ꎻ半纤维素含量从19.92%下降到13.23%ꎬ纤维素含量由7.41%降低到5.85%ꎻ大豆球蛋白和β-伴球蛋白的降解率可达84.91%和80.95%ꎮ综上ꎬ贝莱斯芽孢杆菌CL-4作为发酵豆粕的新型菌种资源ꎬ可有效降解豆粕中抗营养因子ꎬ提高豆粕营养品质和饲料效率ꎮ关键词:贝莱斯芽孢杆菌ꎻ固态发酵ꎻ豆粕ꎻ营养品质中图分类号:S816.6㊀㊀文献标识号:A㊀㊀文章编号:1001-4942(2023)09-0087-07EffectsofSolidStateFermentationwithBacillusvelezensisCL ̄4onNutritionalQualityofSoybeanMealQuZihuiꎬLiuXinꎬZhengLinꎬWeiBingdongꎬYanXiaogangꎬYuWeiꎬChenLongꎬLangHongyan(InstituteofAnimalNutritionandFeedꎬJilinAcademyofAgriculturalSciencesꎬGongzhulingꎬ136100ꎬChina)Abstract㊀InthisexperimentꎬBacillusvelezensisCL ̄4isolatedandidentifiedbytheAnimalNutritionandFeedResearchInstituteofJilinAcademyofAgriculturalScienceswasusedforsolidfermentationofsoy ̄beanmeal.Thenutrientcompositionsꎬsoybeanantigenproteinꎬenzymeactivityꎬviablebacteriacountꎬanti ̄bacterialactivityandapparentformofsoybeanmealweredeterminedbeforeandafterfermentationinordertoevaluatethenutritionalqualityimprovementeffectofsolidstatefermentationwithB.velezensisCL ̄4.There ̄sultsshowedthatB.velezensisCL ̄4showedalargediameterhydrolyticringonthesoybeanantigenproteinscreeningplateꎬwhichhadtheabilitytodegradesoybeanantigenprotein.Thenutritionalqualityandfunction ̄almetabolitesofsoybeanmealweresignificantlyimprovedby24hsolidfermentationꎬandthecontentsofacid ̄solubleproteinꎬcalciumꎬashandtotalphosphoruswerehigher.Thecrudeproteincontentincreasedfrom46.78%to51.28%ꎬandthetotalaminoacidcontentincreasedfrom41.72%to48.14%.Hemicellulosecon ̄tentdecreasedfrom19.92%to13.23%ꎬandcellulosecontentdecreasedfrom7.41%to5.85%.Thedegrada ̄tionratesofsoybeanglycininandβ ̄conglycinincouldreach84.91%and80.95%ꎬrespectively.Inconclu ̄sionꎬB.velezensisCL ̄4ꎬasanewstrainresourceforfermentationofsoybeanmealꎬcouldeffectivelydegradeanti ̄nutritionfactorsinsoybeanmealꎬandimprovethenutritionalqualityandfeedefficiencyofsoybeanmeal.Keywords㊀BacillusvelezensisꎻSolidstatefermentationꎻSoybeanmealꎻNutritionalquality㊀㊀豆粕是食品和饲料领域常见的优质植物性蛋白来源ꎮ豆粕中主要的抗原蛋白是大豆球蛋白和β-伴球蛋白ꎬ分别占豆粕总蛋白的30%和40%左右[1]ꎮ当幼龄仔猪摄入这类蛋白质时ꎬ会引起过敏ꎬ导致吸收不良综合征㊁生长抑制和腹泻ꎮ此外ꎬ豆粕中还含有非淀粉多糖ꎬ主要由纤维素㊁半纤维素和果胶组成ꎬ被证实是导致断奶仔猪肠道疾病的诱因[2]ꎮ发酵豆粕通过添加有益微生物ꎬ如少孢根霉(Rhizopusoligosporus)㊁米曲霉(Asper ̄gillusoryzae)㊁短乳杆菌(Lactobacillusbrevis)或枯草芽孢杆菌(Bacillussubtilis)ꎬ可以有效去除部分对动物有害的抗营养因子ꎬ从而改善豆粕营养品质ꎬ提高动物消化利用率[3-5]ꎮ贝莱斯芽孢杆菌(Bacillusvelezensis)作为芽孢杆菌中新划分的一个种ꎬ于2016年与B.meth ̄ylotrophicus㊁B.amyloliquefacienssubsp.plantarum㊁B.oryzicola重新归类并命名为B.velezensis[6]ꎮ有关B.velezensis的研究集中于生物防治和促进植物生长等方面[7-8]ꎮ贝莱斯芽孢杆菌于2020年被列入欧盟安全资格认定(QPS)推荐的生物制剂列表中ꎬ可作为新型发酵饲料菌种[9]ꎬ有关B.velezensis在畜禽应用的报道逐渐增多[10]ꎬ主要集中在饲料霉菌毒素[玉米赤霉烯酮(zearalenoneꎬZEN)和黄曲霉毒素B1(AFB1)]脱毒[11]和水产益生菌方面[12]ꎮ本研究团队主要开展有关B.velezensis在木质纤维素利用方面的研究ꎬ前期成功分离并鉴定一株来自鸡盲肠内容物的B.velezensisCL-4ꎬ具有富产木质纤维素酶优势ꎬ同时具有抑制病原细菌和真菌的能力ꎬ对动物安全无毒ꎬ具有良好的益生特性[13]ꎮ现已完成了该菌株的全基因组测序ꎬ并成功用于发酵玉米胚芽粕ꎬ获得授权发明专利«一株禽源贝莱斯芽孢杆菌CL-4及其应用»(专利号:202110109964.X)ꎮ豆粕常用发酵菌多为枯草芽孢杆菌[2]㊁酿酒酵母菌[14]㊁植物乳杆菌[15]等ꎬ仅有少数文献报道了贝莱斯芽孢杆菌发酵豆粕的应用[16]ꎮ因此ꎬ本研究利用B.velezensisCL-4发酵豆粕ꎬ探究发酵前后豆粕中抗营养因子㊁营养成分㊁微生物㊁酶活力以及表观形态等变化ꎬ旨在为生物蛋白饲料提供新型优良菌种ꎬ为进一步改善发酵豆粕营养品质提供理论依据和数据支撑ꎮ1㊀材料与方法1.1㊀试验材料1.1.1㊀菌株和发酵原料㊀菌种贝莱斯芽孢杆菌(B.velezensis)CL-4分离自肉鸡盲肠内容物ꎬ病原指示菌为金黄色葡萄球菌ATCC25923㊁大肠埃希菌ATCC25922ꎬ均由吉林省农业科学院动物营养与饲料研究所保存ꎬ豆粕购自吉林省公主岭禾丰牧业有限责任公司ꎮ1.1.2㊀主要试剂和仪器㊀DNS试剂㊁LB培养基㊁大豆球蛋白和β-伴球蛋白ELISA试剂盒购自北京龙科方舟生物工程技术有限公司ꎬ植物蛋白提取试剂盒购自南京凯基生物有限公司ꎮ控摇床ꎬ恒温培养箱ꎬ高压灭菌锅ꎬ超净工作台ꎮ1.2㊀试验方法1.2.1㊀豆粕抗原蛋白平板制备及菌株降解能力测定㊀抗原蛋白培养基的制备:称取5g豆粕ꎬ磨碎后过60目筛ꎬ加入pH8.5的Tris-HCl缓冲液75mLꎬ30~50ħ㊁200r/min振荡1hꎬ9000r/min离心40minꎬ沉淀再浸提一次ꎬ合并两次上清液ꎮ向上清液中加入NaHSO3至0.01mol/Lꎬ用2mol/LHCl调pH至6.4ꎬ4ħ沉淀过夜ꎮ于6500r/min㊁4ħ离心30minꎬ得到大豆球蛋白沉淀ꎮ上清液加NaCl至0.25mol/Lꎬ调pH至4.0~6.0ꎬ室温搅拌30minꎬ9000r/min㊁4ħ离心30minꎬ上清液稀释2倍ꎬ调pH至4.8ꎬ6500r/min离心20min得到β-伴球蛋白沉淀ꎮ将所有沉淀溶于ddH2Oꎬ调pH至5.5~6.5ꎬ加入1.5%(w/v)琼脂ꎬ115ħ灭菌20minꎮ抗原蛋白平板制备:在灭菌培养皿中加入15mL抗原蛋白培养基ꎬ待冷却后再加入营养培养基(蛋白胨10g/L㊁牛肉膏3g/L㊁氯化钠5g/L㊁琼脂20g/Lꎬ121ħ高压灭菌15min)15mLꎬ冷却至凝固ꎬ待培养基表面无明显水迹后ꎬ将已灭菌的牛津杯置于试验平板中ꎬ轻轻加压ꎬ使其与培养皿接触无空隙ꎬ4ħ保存备用ꎮ菌株降解豆粕抗原蛋白能力测定:根据88㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀Wongputtisin等[17]的方法制备候选菌株B.velezensisCL-4粗上清液ꎬ过0.22μm微孔滤膜ꎮ取100μL粗上清液加入抗原蛋白筛选平板的牛津杯中培养24hꎬ以添加100μL生理盐水为对照ꎮ若菌株对抗原蛋白有降解作用ꎬ即可见到水解圈ꎮ根据水解圈直径与牛津杯孔径比值测定B.velezensisCL-4的豆粕抗原蛋白降解能力ꎮ1.2.2㊀发酵豆粕的制备㊀将B.velezensisCL-4在37ħ液体LB培养基中培养12h以备固态发酵ꎮ将豆粕121ħ高压灭菌处理20minꎬ称取灭菌后的豆粕100g于500mL烧瓶中ꎬ发酵菌种添加量为107CFU/gꎬ最终含水量为40%ꎬ搅拌均匀后用滤菌呼吸膜封住瓶口于37ħ下发酵24hꎬ然后105ħ㊁30min阻断发酵ꎮ以0.85%无菌生理盐水为对照ꎬ重复3次ꎮ将发酵样品65ħ烘干24hꎬ冷却研磨过60目筛ꎬ用于SDS-PAGE和营养成分分析ꎮ1.2.3㊀pH值和发酵代谢产物相关指标测定㊀准确称取0㊁24h的发酵样品各1.00g溶于9.0mL蒸馏水中ꎬ室温150r/min振荡10minꎬ静置1min后测定pH值ꎻ采用倍比稀释法测定发酵样品中活菌数ꎻ通过DNS法测定纤维素酶㊁木聚糖酶和果胶酶活力ꎬ中性蛋白酶活力测定参考行业标准SB/T10317 1999ꎻ使用琼脂扩散法测定发酵后豆粕的抑菌性ꎬ以金黄色葡萄球菌ATCC25923和大肠埃希菌ATCC25922作为抑菌试验的指示剂ꎮ1.2.4㊀营养成分分析㊀根据AOAC(2005)测定发酵前后豆粕中干物质㊁粗纤维㊁粗蛋白㊁纤维素㊁半纤维素㊁总磷㊁钙和灰分等含量ꎮ根据Ovissipour等[18]的方法测定发酵前后豆粕中酸溶蛋白含量ꎮ采用氨基酸自动分析仪测定发酵前后豆粕中氨基酸含量ꎮ1.2.5㊀豆粕抗原蛋白定量检测㊀利用间接竞争性ELISA法测定发酵前后豆粕中大豆球蛋白和β-伴球蛋白含量ꎬ采用北京龙科方舟试剂盒进行ꎮ1.2.6㊀SDS-PAGE分析㊀根据植物蛋白提取试剂盒说明书提取发酵0㊁12㊁24h豆粕可溶性蛋白ꎬ使用Bio-Rad蛋白定量试剂盒将上清液定量至50μg/mLꎬ分别配制12%分离胶和5%浓缩胶ꎬ采用稳流35mA电泳至蛋白进入分离胶ꎬ然后设定稳流45mA电泳至溴酚蓝离胶底1cmꎬ最后采用考马斯亮蓝染色和脱色液脱色ꎬ直至凝胶背景脱净ꎮ1.2.7㊀扫描电镜观察㊀取发酵前后豆粕样品0.1g包裹于滤纸内ꎬ用2.5%戊二醛4ħ浸泡过夜ꎮ扫描电镜观察倍数分别为1000㊁1500㊁3000ꎮ1.3㊀数据统计与分析使用SPSS软件(24.0)通过Student st-test和单因素方差分析(ANOVA)对数据进行统计分析ꎬ各组间数据显著差异水平设定为P<0.05ꎬ数值表示为平均值ʃ标准差ꎮ2㊀结果与分析2.1㊀B.velezensisCL-4降解豆粕抗原蛋白能力测定如图1所示ꎬB.velezensisCL-4胞外上清液在大豆抗原蛋白筛选板上显示出较大直径水解圈ꎬ而生理盐水对照没有出现水解圈ꎬ初步推断B.velezensisCL-4胞外上清液具有降解大豆抗原蛋白的能力ꎮa和b为生理盐水对照ꎬc和d为B.velezensisCL-4胞外上清液ꎮ图1㊀B.velezensisCL-4降解豆粕抗原蛋白能力2.2㊀豆粕发酵前后营养成分比较分析如表1所示ꎬ与发酵前相比ꎬB.velezensisCL-4固态发酵24h后ꎬ豆粕干物质含量由93.25%ʃ0.36%下降至92.69%ʃ0.32%ꎬ粗蛋白含量由46.78%ʃ0.32%增加到51.28%ʃ0.24%ꎬ酸溶蛋白含量由5.15%ʃ0.04%显著提升至10.74%ʃ0.12%ꎬ钙㊁灰分和总磷含量均有所提高ꎮ粗纤维含量显著降低ꎬ其中半纤维素含量从19.92%ʃ0.11%下降到13.23%ʃ0.09%ꎬ纤维素含量由7.41%ʃ0.05%降低到5.85%ʃ0.08%ꎮ各种氨基酸含量均呈上升趋势ꎬ除精氨酸㊁蛋氨酸㊁丙氨酸㊁酪氨酸和脯氨酸外ꎬ其他必需和非必需氨基酸显著提升(P<0.05)ꎮ与原始豆粕相比ꎬ固态发酵饲料的总氨基酸含量由41.72%ʃ0.40%显著提高至98㊀第9期㊀㊀㊀㊀㊀瞿子惠ꎬ等:贝莱斯芽孢杆菌CL-4固态发酵对豆粕营养品质的影响48.14%ʃ0.14%ꎮ因此ꎬB.velezensisCL-4固态发酵可显著提高豆粕营养品质ꎬ降低粗纤维含量ꎮ㊀㊀表1㊀B.velezensisCL-4发酵前后豆粕营养成分分析%成分原始豆粕发酵豆粕干物质93.25ʃ0.36a92.69ʃ0.32b粗蛋白46.78ʃ0.32b51.28ʃ0.24a酸溶蛋白5.15ʃ0.04b10.74ʃ0.12a粗纤维5.49ʃ0.05a5.12ʃ0.08b纤维素7.41ʃ0.05a5.85ʃ0.08b半纤维素19.92ʃ0.11a13.23ʃ0.09b灰分6.14ʃ0.06b6.68ʃ0.05a钙0.33ʃ0.01b0.36ʃ0.01a总磷0.61ʃ0.01b0.72ʃ0.01a必需氨基酸精氨酸3.19ʃ0.03a3.23ʃ0.02a组氨酸1.07ʃ0.02b1.27ʃ0.01a异亮氨酸1.99ʃ0.05b2.25ʃ0.01a亮氨酸3.62ʃ0.04b4.03ʃ0.03a赖氨酸2.54ʃ0.02b2.88ʃ0.01a蛋氨酸0.26ʃ0.01a0.32ʃ0.03a苯丙氨酸2.09ʃ0.02b2.42ʃ0.02a苏氨酸1.76ʃ0.03b2.00ʃ0.01a缬氨酸2.13ʃ0.07b2.52ʃ0.03a非必需氨基酸天冬氨酸5.14ʃ0.01b5.65ʃ0.03a丝氨酸2.18ʃ0.02b2.45ʃ0.01a谷氨酸7.79ʃ0.01b9.43ʃ0.06a甘氨酸1.94ʃ0.04b2.29ʃ0.01a丙氨酸1.98ʃ0.04a2.03ʃ0.02a半胱氨酸0.41ʃ0.01b0.53ʃ0.01a酪氨酸1.21ʃ0.02a1.38ʃ0.01a脯氨酸2.33ʃ0.03a2.47ʃ0.02a总氨基酸含量41.72ʃ0.40b48.14ʃ0.14a㊀㊀注:同行数据肩标不同大㊁小写字母分别表示差异极显著(P<0.01)㊁显著(P<0.05)ꎬ下同ꎮ2.3㊀发酵豆粕抗菌活性图2显示ꎬ与未发酵豆粕的上清液相比ꎬ固态发酵24h后的豆粕上清液在MH固体培养基上对金黄色葡萄球菌ATCC25923和大肠埃希菌ATCC25922具有明显的抑菌圈ꎮ因此ꎬB.velezen ̄sisCL-4固态发酵豆粕具有一定的抗菌活性ꎮ2.4㊀发酵豆粕pH值㊁活菌数及酶活力变化由表2可知ꎬ与发酵前相比ꎬB.velezensisCL-4固态发酵豆粕的活菌数从(8.13ʃ0.04)logCFU/g显著增加到(10.28ʃ0.29)logCFU/gꎻpH值从6.64ʃ0.02小幅增加到7.01ʃ0.05ꎻ纤维素酶活力由(7.57ʃ0.41)U/g提升至(18.73ʃ1.67)U/gꎬ木聚糖酶活力由(7.21ʃ0.48)U/g提升至(23.92ʃ1.48)U/gꎬ果胶酶活力由(5.52ʃ0.38)U/g上升至(14.05ʃ2.71)U/gꎬβ-甘露聚糖酶活力由(6.52ʃ0.12)U/g提升至(17.64ʃ0.84)U/gꎬ中性蛋白酶活力由(7.90ʃ0.74)U/g提升至(235.93ʃ10.19)U/gꎬ各种酶活力均显著提高ꎮ1㊁2㊁3为B.velezensisCL-4胞外上清液重复ꎮ图2㊀B.velezensisCL-4固态发酵豆粕的抗菌活性㊀㊀表2㊀B.velezensisCL-4发酵豆粕pH值㊁活菌数及酶活力变化项目原始豆粕发酵豆粕pH值6.64ʃ0.02a7.01ʃ0.05a活菌数/(logCFU/g)8.13ʃ0.04b10.28ʃ0.29a纤维素酶活力/(U/g)7.57ʃ0.41b18.73ʃ1.67a木聚糖酶活力/(U/g)7.21ʃ0.48b23.92ʃ1.48a果胶酶活力/(U/g)5.52ʃ0.38b14.05ʃ2.71a中性蛋白酶活力/(U/g)7.90ʃ0.74B235.93ʃ10.19Aβ-甘露聚糖酶活力/(U/g)6.52ʃ0.12b17.64ʃ0.84a2.5㊀发酵豆粕抗原蛋白降解效果SDS-PAGE分析结果(图3)显示ꎬ在24h发酵过程中ꎬ豆粕分子量大于35kDa的蛋白亚基逐步降解ꎬ而15~24kDa的蛋白含量逐渐提高ꎮ大豆抗原蛋白亚基中的β-伴球蛋白亚基(α和αᶄ)分子量在70~100kDa左右ꎬ发酵12h基本降解ꎬ1㊁2㊁3分别代表发酵0㊁12㊁24hꎮ图3㊀B.velezensisCL-4发酵豆粕可溶性蛋白分子量变化09㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀而β-伴球蛋白βᶄ亚基分子量为60kDa左右ꎬ于24h被降解ꎮ因此ꎬB.velezensisCL-4固态发酵可将豆粕中大分子抗原蛋白降解成小分子肽类ꎮELISA定量检测结果(表3)显示ꎬ与发酵前相比ꎬB.velezensisCL-4固态发酵24h后ꎬ豆粕中大豆球蛋白含量由(176.14ʃ3.15)mg/g降低至(26.58ʃ1.22)mg/gꎬ降解率可达84.91%ꎻβ-伴球蛋白含量由(134.66ʃ2.24)mg/g下降至(25.65ʃ0.75)mg/gꎬ降解率可达80.95%ꎮ表明B.velezensisCL-4固态发酵可显著降低豆粕中大豆球蛋白和β-伴球蛋白含量ꎮ2.6㊀发酵过程中豆粕表观形态变化扫描电镜观察结果(图4)显示ꎬ发酵前豆粕结构紧凑㊁表面光滑ꎮB.velezensisCL-4固态发酵24h后ꎬ豆粕的块状结构被大量分解ꎬ呈现出碎片㊁破裂和多纤维素结构ꎬ表明B.velezensisCL-4固态发酵可明显改变豆粕表观形态ꎬ有效降解木质纤维素ꎮ㊀㊀表3㊀B.velezensisCL-4发酵豆粕抗原蛋白的降解效果项目大豆球蛋白含量/(mg/g)降解率/%β-伴球蛋白含量/(mg/g)降解率/%原始豆粕176.14ʃ3.15a134.66ʃ2.24a发酵豆粕26.58ʃ1.22b84.9125.65ʃ0.75b80.95㊀㊀注:同列数据肩标不同字母表示差异显著(P<0.05)ꎮA~C分别代表原始豆粕放大1000㊁1500㊁3000倍ꎻD~F分别代表发酵24h豆粕放大1000㊁1500㊁3000倍ꎮ图4㊀B.velezensisCL-4发酵过程中豆粕形态变化3㊀讨论与结论豆粕来源广泛且营养丰富ꎬ是动物饲料中主要的植物源性蛋白资源ꎮ然而ꎬ豆粕中含有多种抗营养因子ꎬ限制了其在幼龄动物日粮中的广泛应用[17]ꎮ研究表明微生物发酵可以部分降解豆粕中抗营养因子ꎬ从而改善其营养品质[19-20]ꎮ本研究中ꎬ抗原蛋白平板法测定验证了新型菌种B.velezensisCL-4可降解豆粕抗原蛋白ꎬB.velezensisCL-4固态发酵24h后ꎬ豆粕中大豆球蛋白和β-伴球蛋白的降解率可分别达84.91%和80.95%ꎮ由于原料在发酵前已经灭菌且发酵过程也是无菌的ꎬ不涉及外源或天然微生物影响ꎬ因而B.velezensisCL-4在豆粕发酵过程中发挥主要作用ꎮSDS-PAGE测定的豆粕可溶性蛋白分子量变化与酶联免疫吸附法测定的大豆球蛋白和β-伴球蛋白在发酵过程中的降解趋势一致ꎮ此前研究也在B.velezensisCL-4全基因组序列中检测到丝氨酸蛋白酶㊁氨基肽酶㊁金属蛋白酶等多种蛋白水解酶的基因[13]ꎮ在酶活力检测中也发现ꎬB.velezensisCL-4中性蛋白酶活性显著提高ꎬ有效分解豆粕中抗原蛋白ꎮWang等[4]采用两段发酵法通过枯草芽孢杆菌CW4和粪肠球菌CWEF发酵豆粕和玉米混合底物ꎬ营养价值显著提高ꎮYao等[21]发现枯草芽孢杆菌N-11厌氧发酵豆粕可增加酸溶蛋白(ASP)含量ꎬ最高达到13.48%ꎬ大19㊀第9期㊀㊀㊀㊀㊀瞿子惠ꎬ等:贝莱斯芽孢杆菌CL-4固态发酵对豆粕营养品质的影响豆球蛋白和β-伴球蛋白分别降低82.38%和88.32%ꎮShi等[2]发现在玉米-豆粕混合饲料中接种枯草芽孢杆菌B.subtilis和屎肠杆菌E.faeci ̄umꎬ大豆球蛋白和β-伴球蛋白的降解率分别为86.12%和77.53%ꎮ以上研究与本试验结果一致ꎬ在后续研究中还需要通过2DE电泳和蛋白质组学对发酵产物中的蛋白质作进一步研究ꎮ本研究中ꎬB.velezensisCL-4固态发酵豆粕与原始豆粕相比含有更高含量的粗蛋白和氨基酸含量ꎬ与前人的报道一致[3ꎬ22]ꎮ发酵过程中干物质的损失也可能导致粗蛋白和氨基酸的增加[23]ꎮB.velezensisCL-4可显著提高豆粕中酸溶蛋白含量主要是由于在发酵过程中ꎬ豆粕抗原蛋白或其他蛋白水解形成小分子肽和游离氨基酸[24]ꎮ本研究中ꎬB.velezensisCL-4发酵豆粕对金黄色葡萄球菌ATCC25923和大肠埃希菌ATCC25922具有一定抑制能力ꎬ可部分替代饲料中的抗生素ꎮ本研究中ꎬ与原始豆粕相比ꎬ发酵豆粕中纤维素和半纤维素降解率分别为21.05%和33.58%ꎮ在豆粕发酵过程中几种非淀粉多糖降解酶(纤维素酶㊁木聚糖酶㊁β-甘露聚糖酶和果胶酶)的活力均显著上升ꎮ扫描电镜观察发现与原始豆粕相比ꎬ发酵豆粕表面结构呈现开裂和多孔结构ꎬ说明其中木质纤维素组分可能被部分降解ꎬ而这与非淀粉多糖降解酶密切相关ꎮ此外ꎬ随着纤维素和半纤维素的降解ꎬ豆粕中蛋白组分更容易被B.velezensisCL-4分泌的蛋白酶所分解ꎬ因此发酵豆粕可能会具有更高的养分消化率ꎮ目前生物发酵饲料常用的发酵菌种为芽孢杆菌㊁乳酸菌以及酵母菌ꎮ中国生物饲料产业创新战略联盟最新发布并实施的«发酵饲料技术通则»中明确指出发酵饲料菌种只允许添加饲料添加剂品种目录(2013年)规定的相应菌种ꎬ可用菌种约为35种ꎬ而欧盟食品安全局(EFSA)可利用的菌种数量可达80余种[25]ꎮ因而ꎬ新型发酵菌种的研发和应用急需开展ꎮ贝莱斯芽孢杆菌菌株通常从土壤㊁植物根际㊁河流㊁动物肠道和发酵食品等来源分离获得[26]ꎬ其相关研究集中于生物防治和促进植物生长等方面[27-28]ꎮ贝莱斯芽孢杆菌已于2020年被列入欧盟安全资格认定(QPS)推荐的生物制剂列表中ꎬ可作为新型发酵饲料菌种[29]ꎮ全基因组学分析发现B.velezensisCL-4有大量编码木质纤维素降解酶的基因ꎬ其发酵产生的碳水化合物酯酶㊁果胶酸裂解酶和碳水化合物结合模块(CBMs)也可能影响纤维素和半纤维素降解[13]ꎮ此外ꎬ在GH1-13[30]㊁FZB42[8]㊁ZY ̄1 ̄1[31]㊁LS69[32]和UCMB5113[33]等B.velezensis基因组中均发现参与降解纤维素和半纤维素的酶基因ꎮ但有关将B.velezensis应用于动物饲料益生菌和生物发酵饲料中的报道仍然较少ꎮ本研究通过高产蛋白酶和木质纤维素降解酶的B.velezensisCL ̄4发酵豆粕ꎬ可降解豆粕中抗营养因子(大豆抗原蛋白㊁纤维素和半纤维素)ꎬ显著改变了原始豆粕的营养特性ꎬ提高了营养品质和功能代谢物(活菌数㊁酶活力以及抑菌活性)ꎬ可作为新型发酵豆粕菌种ꎬ具有广阔的应用前景ꎮ参㊀考㊀文㊀献:[1]㊀MaruyamaNꎬSatoRꎬWadaYꎬetal.Structurephysicochemi ̄calfunctionrelationshipsofsoybeanbeta ̄conglycininconstitu ̄entsubunits[J].JournalofAgriculturalandFoodChemistryꎬ2000ꎬ48(2):576-580.[2]㊀ShiCYꎬZhangYꎬLuZQꎬetal.Bacillussubtilissolidstatefermentationofcornsoybeanmealmixedfeedwithandforde ̄gradingantinutritionalfactorsandenhancingnutritionalvalue[J].JournalofAnimalScienceandBiotechnologyꎬ2017ꎬ8(4):50-52.[3]㊀FengJꎬLiuXꎬXuZRꎬetal.Effectoffermentedsoybeanmealonintestinalmorphologyanddigestiveenzymeactivitiesinweanedpiglets[J].DigestiveDiseasesandSciencesꎬ2007ꎬ52(8):1845-1850.[4]㊀WangCꎬShiCYꎬSuWFꎬetal.Dynamicsofthephysico ̄chemicalcharacteristicsꎬmicrobiotaꎬandmetabolicfunctionsofsoybeanmealandcornmixedsubstratesduringtwostagesol ̄idstatefermentation[J].Systemsꎬ2020ꎬ5(1)32-35. [5]㊀HongKꎬLeeCꎬKimS.AspergillusoryzaeGB ̄107fermentationimprovesnutritionalqualityoffoodsoybeansandfeedsoybeanmeals[J].JournalofMedicinalFoodꎬ2004ꎬ7(4):430-435. [6]㊀DunlapCAꎬKimSJꎬKwonSWꎬetal.BacillusvelezensisisnotalaterheterotypicsynonymofBacillusamyloliquefaciensꎻBacillusmethylotrophicusꎬBacillusamyloliquefacienssubsp.plantarumand Bacillusoryzicola arelaterheterotypicsyno ̄nymsofBacillusvelezensisbasedonphylogenomics[J].Interna ̄tionalJournalofSystematicandEvolutionaryMicrobiologyꎬ2016ꎬ66(3):1212-1217.[7]㊀AdenijiAAꎬLootsDTꎬBabalolaOO.Bacillusvelezensis:phylogenyꎬusefulapplicationsꎬandavenuesforexploitation[J].AppliedMicrobiologyandBiotechnologyꎬ2019ꎬ103(9):3669-3682.[8]㊀FanBꎬWangCꎬSongXFꎬetal.BacillusvelezensisFZB42in2018:thegram ̄positivemodelstrainforplantgrowthpromotionandbiocontrol[J].FrontiersinMicrobiologyꎬ2018ꎬ9:2491-29㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第55卷㊀2502.[9]㊀KoutsoumanisKꎬAllendeAꎬAlvarez ̄OrdóñezAꎬetal.Up ̄dateofthelistofQPS ̄recommendedbiologicalagentsintention ̄allyaddedtofoodorfeedasnotifiedtoEFSA11:suitabilityoftaxonomicunitsnotifiedtoEFSAuntilSeptember2019[J].EFSAJ.ꎬ2020ꎬ18(2):05965.[10]KhalidFꎬKhalidAꎬFuYꎬetal.PotentialofBacillusvelezen ̄sisasaprobioticinanimalfeed:areview[J].JournalofMi ̄crobiology(SeoulꎬKorea)ꎬ2021ꎬ59(7):627-633. [11]WangMYꎬHuangSꎬChenJꎬetal.Completegenomese ̄quenceofzearalenonedegradingbacteriaBacillusvelezensisA2[J].CurrentMicrobiologyꎬ2021ꎬ78(1):347-350. [12]ZhangDXꎬKangYHꎬZhanSꎬetal.EffectofBacillusvelezensisonAeromonasveronii ̄inducedintestinalmucosalbarri ̄erfunctiondamageandinflammationincruciancarp(Carassiusauratus)[J].FrontiersinMicrobiologyꎬ2019ꎬ10:2663-2670. [13]ChenLꎬChenWYꎬZhengBYꎬetal.FermentationofNaH ̄CO3treatedcorngermmealbyBacillusvelezensisCL ̄4pro ̄moteslignocellulosedegradationandnutrientutilization[J].AppliedMicrobiologyandBiotechnologyꎬ2022ꎬ106(18):6077-6094.[14]ChenKLꎬKhoWLꎬYouSHꎬetal.EffectsofBacillussubti ̄lisvar.nattoandSaccharomycescerevisiaemixedfermentedfeedontheenhancedgrowthperformanceofbroilers[J].PoultryScienceꎬ2009ꎬ88(2):309-315.[15]YehRHꎬHsiehCWꎬChenKL.Screeninglacticacidbacte ̄riatomanufacturetwo ̄stagefermentedfeedandpelletingtoin ̄vestigatethefeedingeffectonbroilers[J].PoultryScienceꎬ2018ꎬ97(1):236-246.[16]LiuZYꎬGuanXFꎬZhongXXꎬetal.BacillusvelezensisDP ̄2isolatedfromDouchianditsapplicationinsoybeanmealfer ̄mentation[J].JournaloftheScienceofFoodandAgricultureꎬ2021ꎬ101(5):1861-1868.[17]WongputtisinPꎬKhanongnuchCꎬKhongbantadWꎬetal.ScreeningandselectionofBacillusspp.forfermentedcorticatesoybeanmealproduction[J].JournalofAppliedMicrobiologyꎬ2012ꎬ113(4):798-806.[18]OvissipourMꎬAbedianAꎬMotamedzadeganAꎬetal.TheeffectofenzymatichydrolysistimeandtemperatureonthepropertiesofproteinhydrolysatesfromPersiansturgeon(Acipenserpersicus)viscera[J].FoodChemistryꎬ2013ꎬ115(1):238-242. [19]MedeirosSꎬXieJJꎬDycePWꎬetal.Isolationofbacteriafromfermentedfoodandgrasscarpintestineandtheirefficien ̄ciesinimprovingnutrientvalueofsoybeanmealinsolidstatefermentation[J].Anim.Sci.Biotechnol.ꎬ2018ꎬ9:29-32. [20]ZhengLꎬLiDꎬLiZLꎬetal.EffectsofBacillusfermentationontheproteinmicrostructureandanti ̄nutritionalfactorsofsoy ̄beanmeal[J].LettersinAppliedMicrobiologyꎬ2017ꎬ65(6):520-526.[21]YaoYHꎬLiHYꎬLiJꎬetal.Anaerobicsolid ̄statefermenta ̄tionofsoybeanmealwithBacillussp.toimprovenutritionalquality[J].FrontiersinNutritionꎬ2021ꎬ8:706977-706980. [22]FriasJꎬSongYSꎬMartínez ̄VillaluengaCꎬetal.Immunoreac ̄tivityandaminoacidcontentoffermentedsoybeanproducts[J].Agric.FoodChem.ꎬ2008ꎬ56(1):99-105.[23]ShiCYꎬZhangYꎬYinYꎬetal.Bacillussubtilisaminoacidandphosphorusdigestibilityoffermentedcornsoybeanmealmixedfeedwithandfedtopigs[J].JournalofAnimalScienceꎬ2017ꎬ95(9):3996-4004.[24]GilbertEꎬWongEꎬWebbK.Boardinvitedreview:peptideab ̄sorptionandutilization:implicationsforanimalnutritionandhealth[J].JournalofAnimalScienceꎬ2008ꎬ86(9):2135-2155.[25]蔡辉益ꎬ刘世杰ꎬ邓雪娟ꎬ等.生物饲料团体标准开启产业健康发展新时代[J].中国畜牧杂志ꎬ2018ꎬ54(9):143-146.[26]Ruiz ̄GarciaCꎬBejarVꎬMartinez ̄ChecaFꎬetal.Bacillusvelezensissp.nov.ꎬasurfactant ̄producingbacteriumisolatedfromtheriverVelezinMalagaꎬsouthernSpain[J].Interna ̄tionalJournalofSystematicandEvolutionaryMicrobiologyꎬ2005ꎬ55(1):191-195.[27]YeMꎬTangXFꎬYangRꎬetal.Characteristicsandapplica ̄tionofanovelspeciesofBacillus:Bacillusvelezensis[J].ACSChemicalBiologyꎬ2018ꎬ13(3):500-505.[28]RabbeeMFꎬAliMSꎬChoiJꎬetal.Bacillusvelezensis:aval ̄uablememberofbioactivemoleculeswithinplantmicrobiomes[J].Moleculesꎬ2019ꎬ24(6):14-16.[29]KoutsoumanisKꎬAllendeAꎬAlvarez ̄OrdóñezAꎬetal.Scien ̄tificopinionontheupdateofthelistofQPS ̄recommendedbio ̄logicalagentsintentionallyaddedtofoodorfeedasnotifiedtoEFSA(2017-2019)[J].EFSAJ.ꎬ2020ꎬ18(2):05966. [30]KimSYꎬSongHꎬSangMKꎬetal.Thecompletegenomese ̄quenceofBacillusvelezensisstrainGH1 ̄13revealsagricultural ̄lybeneficialpropertiesandauniqueplasmid[J].JournalofBi ̄otechnologyꎬ2017ꎬ259:221-227.[31]ZhangZYꎬRazaMFꎬZhengZꎬetal.Completegenomese ̄quenceofBacillusvelezensisZY ̄1 ̄1revealsthegeneticbasisforitshemicellulosic/cellulosicsubstrateinduciblexylanaseandcellulaseactivities[J].Biotech.ꎬ2018ꎬ8(11):465-469. [32]LiuGQꎬKongYYꎬFanYJꎬetal.Wholegenomesequen ̄cingofBacillusvelezensisLS69ꎬastrainwithabroadinhibitoryspectrumagainstpathogenicbacteria[J].JournalofBiotechnol ̄ogyꎬ2017ꎬ249:20-24.[33]NiaziAꎬManzoorSꎬAsariSꎬetal.GenomeanalysisofBacil ̄lusamyloliquefacienssubsp.plantarumUCMB5113:arhi ̄zobacteriumthatimprovesplantgrowthandstressmanagement[J].PLoSONEꎬ2014ꎬ9(8):e104651.39㊀第9期㊀㊀㊀㊀㊀瞿子惠ꎬ等:贝莱斯芽孢杆菌CL-4固态发酵对豆粕营养品质的影响。

吉林省科学技术厅关于调整第六届吉林省科学技术奖励委员会委员的通知正文:---------------------------------------------------------------------------------------------------------------------------------------------------- 关于调整第六届吉林省科学技术奖励委员会委员的通知各有关单位:经省政府批准,第六届吉林省科学技术奖励委员会于2019年组建,本年度因部分委员工作变动等原因,已不能履行委员职能,按照《吉林省科学技术奖励办法》要求,经省科技厅提名报请省政府批准,对第六届吉林省科学技术奖励委员会委员进行调整,现将调整后的第六届吉林省科学技术奖励委员会委员名单公布如下:主任委员:安立佳男吉林省人民政府副省长、中国科学院院士副主任委员:于化东男吉林省科学技术厅厅长委员:张希男吉林大学校长、中国科学院院士XXX 男中国科学院长春光学精密机械与物理研究所研究员、中国科学院院士陈学思男中国科学院长春应用化学研究所研究员、中国科学院院士李玉男吉林农业大学教授、中国工程院院士姜会林男长春理工大学教授、中国工程院院士金宁一男军事科学院军事医学研究院军事兽医研究所研究员、中国工程院院士张志勇男吉林省发展和改革委员会二级巡视员孙长智男吉林省教育厅副厅长李道恒男吉林省科学技术厅副厅长王大宁男吉林省工业和信息化厅副厅长赵春林男吉林省人力资源和社会保障厅副厅长王慧群男吉林省财政厅副厅长贾平男中国科学院长春光学精密机械与物理研究所所长、研究员杨小牛男中国科学院长春应用化学研究所所长、研究员姜明男中国科学院东北地理与农业生态研究所所长、研究员董英山男吉林省农业科学院院长、研究员刘益春男东北师范大学校长、教授杨华民男长春理工大学校长、教授蔡国伟男东北电力大学校长、教授冯江男吉林农业大学校长、教授宋柏林男长春中医药大学校长、教授秘书长:李道恒(兼)吉林省科学技术厅2021年5月21日——结束——。

吉林省人民政府关于公布第九批农业产业化省级重点龙头企业名单的通知文章属性•【制定机关】吉林省人民政府•【公布日期】2011.01.06•【字号】吉政函[2011]4号•【施行日期】2011.01.06•【效力等级】地方规范性文件•【时效性】现行有效•【主题分类】正文吉林省人民政府关于公布第九批农业产业化省级重点龙头企业名单的通知(吉政函〔2011〕4号)各市(州)人民政府,长白山管委会,各县(市)人民政府,省政府各厅委办、各直属机构:为深入实施“把农业产业化经营作为农业现代化主推模式”的战略部署,加快培植一批强势龙头企业,形成群体规模优势,提升带动功能,根据《吉林省农业产业化省级重点龙头企业认定和运行监测管理暂行办法》(吉农产发〔2002〕1号)的规定,省农业产业化领导小组办公室组织开展了第九批农业产业化省级重点龙头企业评定工作。
依据评审结果并经省政府审定,吉林德生牧业有限公司等77户企业为农业产业化省级重点龙头企业。
农业产业化省级重点龙头企业是推动农业产业化经营和农产品加工业又好又快发展的骨干力量,在调整优化农业和农村产业结构、转变农业增长方式、增加农民收入、保障农产品质量安全、提高我省农业国际竞争力、推进社会主义新农村建设等方面具有重要作用。
农业产业化省级重点龙头企业要按照省委、省政府的部署,紧紧抓住发展这一主题,立足农村、面向农业、服务农民,科学谋划发展项目,积极转变增长方式,着力提升企业核心竞争力和辐射带动功能,进一步探索完善与基地农户的有效利益联结机制,增强责任意识,强化系列服务,自觉维护农民利益,诚信守法经营,为推进我省农业现代化和社会主义新农村建设做出新的贡献。
各地、各有关部门要深入贯彻落实省委、省政府《关于进一步推进农业产业化经营的意见》(吉发〔2005〕27号),进一步优化发展环境,加大扶持力度,完善政策措施,形成整体合力。
要注重引导并发挥好龙头企业在开拓市场、引导生产、深化加工、创新科技、强化服务等方面的带动作用,提升农业和农村经济的整体质量和水平。

关于编制《吉林省科学技术奖励推荐工作手册》的说明为做好本年度吉林省科学技术奖励推荐工作,帮助科技管理人员、科技人员了解吉林省科学技术奖推荐书填写、推荐书附件材料及推荐系统使用方法等相关要求,正确填报吉林省科学技术奖推荐书材料,我们编制了《吉林省科学技术奖励推荐工作手册》。
本手册主要内容包括:吉林省科学技术奖励年度工作日程、吉林省科学技术奖推荐书及填写说明、吉林省科学技术奖励推荐系统使用说明、吉林省科学技术奖评价指标以及吉林省科学技术奖学科(专业)评审组评审范围等内容。
《吉林省科学技术奖励推荐工作手册》是我办编制的第三部手册,如有不当之处,希望各级领导、专家和科技奖励管理人员提出宝贵意见,以便我们今后不断改进和完善相关工作,进一步提高服务质量和管理水平。
吉林省科学技术奖励工作办公室2010年3月吉林省科学技术奖励年度工作日程2月中旬—3月末提交吉林省科技奖项目材料,包括电子版材料和书面材料(逾期不予受理)4月30日前受理项目在省科技厅网站公布5—7月吉林省科学技术特殊贡献奖、科技进步奖学科(专业)评审组网络评审8月吉林省科技厅网站公布初评结果,组织异议项目处理和项目考察9月复审委员会会议10月吉林日报及吉林省科技厅网站公示年度拟获奖项目11月吉林省科学技术奖励委员会会议12月拟授奖项目、授奖人报省政府批准;证书及奖金发放吉林省科学技术特殊贡献奖推荐书( 年度)一、被推荐人基本情况吉林省科学技术奖励工作办公室制二、工作简历三、被推荐人的主要科学技术成就和贡献四、主要的科学发现、技术发明或技术创新要点五、被推荐人发表论文、专著及被引用情况六、被推荐人曾获奖励情况(专家推荐不填此栏)十一、附件1.公开发表的主要论文及专著2.他人引用的论文、专著3.技术鉴定证书及知识产权证明4.被推荐人近期标准照片及工作照片各一张5.其他《吉林省科学技术特殊贡献奖推荐书》填写说明《吉林省科学技术特殊贡献奖推荐书》要严格按规定格式打印或铅印,大小为大十六开本(高297毫米,宽210毫米)竖装。

吉林省农业委员会关于发布吉林省第八届优质食味水
稻品种鉴评结果的通知
文章属性
•【制定机关】吉林省农业委员会
•【公布日期】2017.05.16
•【字号】
•【施行日期】2017.05.16
•【效力等级】地方规范性文件
•【时效性】现行有效
•【主题分类】农业管理综合规定
正文
吉林省农业委员会关于发布吉林省第八届优质食味水稻品种
鉴评结果的通知
各有关单位:
为选育优质水稻品种,助力农业结构调整和现代农业建设,省农委于2017年3月31日组织召开了第八届优质食味水稻品种鉴评会。
此项鉴评活动从年初开始组织,经品种初选、田间鉴评、成分分析、品质鉴评等评鉴环节,从2013年以来全省新审定的117个品种中最后筛选出5个品种进入食味鉴评阶段。
通过食味品评专家打分评鉴,省农委研究决定,参加最后食味鉴评的5个品种全部入选,并设立第八届优质食味水稻鉴评活动一、二、三等奖。
一等奖:吉粳515 (省农科院选育)
二等奖:吉农大538 (吉林农大选育)
通禾66 (通化农科院选育)
三等奖:吉粳528 (省农科院选育)
吉农大138 (吉林农大选育)
希望获奖单位再接再厉,攻坚克难,再创水稻优质食味品种培育工作新佳绩,
同时要加大成果转化力度,使之尽快形成生产力,为农民增收、农业增效和推进全省农业供给侧改革做出更大贡献。
吉林省农业委员会
2017年5月16日。


㊀山东农业科学㊀2022ꎬ54(12):157~162ShandongAgriculturalSciences㊀DOI:10.14083/j.issn.1001-4942.2022.12.024收稿日期:2022-01-11ꎻ修回日期:2022-11-28基金项目:吉林省科技发展计划项目(20200402081NC)ꎻ吉林省农业科技创新工程项目(CXGC2017TD012)作者简介:崔龙(1978 )ꎬ男ꎬ助理研究员ꎬ主要从事核果类果树栽培与育种研究ꎮE-mail:331070480@qq.com通信作者:陈蕾(1982 )ꎬ女ꎬ硕士ꎬ副研究员ꎬ主要从事核果类果树遗传育种与生物技术研究ꎮE-mail:chenleisofia@126.com我国甜樱桃需冷量估算模型应用与效果研究进展崔龙1ꎬ王雪松1ꎬ隋松兵2ꎬ姜源3ꎬ张艳波1ꎬ王薇4ꎬ陈蕾1(1.吉林省农业科学院果树研究所ꎬ吉林公主岭㊀136100ꎻ2.吉林吉科生物高技术有限公司ꎬ吉林公主岭㊀136100ꎻ3.吉林市丰满区吉字号李子家庭农场ꎬ吉林吉林㊀132013ꎻ4.吉林农业大学ꎬ吉林长春㊀130118)㊀㊀摘要:需冷量是甜樱桃引种栽培㊁设施栽培和预测气候变化的关键依据ꎮ本文通过总结ɤ7.2ħ模型㊁0~7.2ħ模型㊁Utah模型和动力学模型在我国甜樱桃需冷量估算上的应用研究ꎬ得出ɤ7.2ħ模型因数值不稳定逐渐被替代ꎬ0~7.2ħ模型因便于操作㊁数值年际间差异小㊁较为稳定而在我国广泛应用ꎻUtah模型和动力学模型虽然考虑到高温和低温对低温累积的影响ꎬ但由于计算复杂并没有得到广泛应用ꎮ同时ꎬ基于中国知网文献数据对不同模型㊁不同地理位置㊁不同品种的需冷量进行比较分析ꎬ阐述需冷量的影响因素ꎬ并对今后研究方向进行展望ꎬ以期为甜樱桃适地适栽及品种选育提供理论依据ꎮ关键词:甜樱桃ꎻ需冷量ꎻ估算模型ꎻ应用效果中图分类号:S662.5㊀㊀文献标识号:A㊀㊀文章编号:1001-4942(2022)12-0157-06ApplicationandEffectAnalysisofChillingRequirementEstimatingModelforSweetCherryinChinaCuiLong1ꎬWangXuesong1ꎬSuiSongbing2ꎬJiangYuan3ꎬZhangYanbo1ꎬWangWei4ꎬChenLei1(1.InstituteofPomologyꎬJilinAcademyofAgriculturalSciencesꎬGongzhuling136100ꎬChinaꎻ2.JilinJikeBiologicalHighTechnologyCo.ꎬLtd.ꎬGongzhuling136100ꎬChinaꎻ3.JiziPlumFamilyFarminFengmanDistrictofJilinCityꎬJilin132013ꎬChinaꎻ4.JilinAgriculturalUniversityꎬChangchun130118ꎬChina)Abstract㊀Chillingrequirementisthekeybasisforsweetcherryintroductionꎬprotectedcultivationandclimatechangeprediction.Bysummarizingtheapplicationofɤ7.2ħmodelꎬ0~7.2ħmodelꎬUtahmodelandKineticmodelintheestimationofchillingrequirementforsweetcherryinChinaꎬitwasconcludedthatɤ7.2ħmodelwasgraduallyreplacedduetonumericalinstabilityꎬwhile0~7.2ħmodelwaswidelyusedinChinabecauseofeasyoperationꎬsmallinterannualdifferenceandstability.AlthoughUtahmodelandKineticmodelconsideredtheinfluenceofhightemperatureandlowtemperatureonchillingrequirementꎬtheywerenotwidelyusedbecauseoftheircomplexcalculation.BasedontheliteraturedataofCNKIꎬthechillingrequire ̄mentobtainedbydifferentmodelsofdifferentvarietiesfromdifferentgeographicallocationswereanalyzedandcomparedꎬandtheinfluencingfactorsandresearchprospectswereexpoundedtoprovidetheoreticalbasesforcultivationatsuitabletimeandplaceandvarietybreedingofsweetcherry.Keywords㊀SweetcherryꎻChillingrequirementꎻEstimatingmodelꎻApplicationeffect㊀㊀甜樱桃(PrunusaviumL.)素有 春果第一枝的美誉ꎬ因风味独特㊁营养丰富㊁经济效益高而深受消费者和栽培者欢迎ꎬ其产区在近20年迅速从山东㊁辽宁等传统种植区扩展至河南㊁山西㊁陕西㊁甘肃㊁江苏等陇海铁路沿线省份以及云㊁贵㊁川等冷凉高地[1]ꎮ随着人民生活水平的提高和果品淡季供应的超高效益ꎬ我国甜樱桃设施栽培迅猛发展ꎬ已形成具有一定规模的甜樱桃特色产业ꎬ在种植业结构调整和果农致富中发挥了重要作用ꎮ据不完全统计ꎬ我国甜樱桃设施栽培面积约有1.3万hm2[2]ꎬ其中ꎬ以塑料大棚种植为主的产地在山东省的临朐和烟台㊁陕西省的澄城等地ꎬ而日光温室种植以辽宁省的大连㊁朝阳等地为主ꎬ黑龙江㊁内蒙古㊁吉林等省(自治区)也均有分布[3]ꎮ需冷量即植物休眠期对低温量的要求ꎬ是在长期自然演变过程中形成的ꎬ已成为预测气候变化㊁确定设施栽培控温时机和因地制宜选择适宜品种的关键依据ꎮ满足需冷量ꎬ保证植物顺利通过自然休眠ꎬ是落叶果树设施栽培成功的关键条件之一[3]ꎮ在传统设施栽培管理条件下ꎬ若对需冷量掌控不准ꎬ就无法准确把握休眠期长短和揭苫升温时间ꎬ而且若需冷量得不到满足ꎬ易造成树体萌芽不一致㊁花器官发育不良㊁坐果率低和果实成熟期延后等问题ꎮ因此ꎬ准确估算甜樱桃自然休眠的需冷量对设施栽培及科学管理具有重要意义ꎮ前人已建立了多种落叶果树需冷量估算模型[4-7]ꎬ为探索适合甜樱桃需冷量估算且应用效果相对稳定的模型ꎬ本文基于中国知网的文献数据ꎬ对甜樱桃需冷量估算模型的应用研究及效果进行了综述分析ꎬ以期为甜樱桃引种栽培和品种选育提供理论依据ꎮ1㊀主要需冷量估算模型介绍目前ꎬ应用于落叶果树需冷量的估算模型主要有ɤ7.2ħ模型[4]㊁0~7.2ħ模型[5]㊁Utah模型[6]和动力学模型[7]ꎮ1.1㊀ɤ7.2ħ模型ɤ7.2ħ模型是1950年WeibergerJ.H.在估算桃需冷量时首次提出[4]ꎮ该估算模型以秋季连续3d日平均温度低于7.2ħ的日期为有效低温累计的起点ꎬ直至自然休眠解除时经历的7.2ħ以下低温的小时数作为需冷量ꎮ1.2㊀0~7.2ħ模型0~7.2ħ模型是基于ɤ7.2ħ模型演变而来的ꎬ以日均温稳定低于7.2ħ的日期为有效低温累积起点ꎬ直至自然休眠解除时经历的0~7.2ħ(不包括0ħ)低温的小时数即为需冷量值ꎮ1.3㊀Utah模型和正Utah模型Utah模型是由美国犹它州立大学的Richard ̄son提出的来计算需冷量的 冷温单位模型 ꎮ在概念上与0~7.2ħ低温模型相似ꎬ只是对低温转换的范围划分更细ꎮ该模型将破眠效率最高的最适冷温1个小时定义为一个冷温单位ꎬ而偏离适期适温且使破眠效率下降甚至具有负作用的温度其冷温单位小于1或为负值ꎮ由于Utah模型适用于冬季温度较低的年份或地区ꎬ但并不适用于暖温带和亚热带气候以及需冷量较低的品种ꎬ为此ꎬ有研究者忽略高温对低温累积的负效应ꎬ又提出了正Utah模型[8]ꎮ两种模型的温度与冷温单位转化关系见表1ꎮ㊀㊀表1㊀㊀Utah和正Utah模型的温度与冷温单位转化关系Utah模型温度(ħ)冷温单位(C.U)正Utah模型温度(ħ)冷温单位(C.U)Tɤ1.40Tɤ1.401.4<Tɤ2.40.51.4<Tɤ2.40.52.4<Tɤ9.11.02.4<Tɤ9.11.09.1<Tɤ12.40.59.1<Tɤ12.40.512.4<Tɤ15.90Tȡ12.4015.9<Tɤ18.0-0.5T>18.0-1.01.4㊀动力学模型动力学模型(dynamicmodel)是Erez在1990年就高温对低温抵消效应的作用机制而提出的[7]ꎬ该模型以低温累计过程中某种中间产物需要形成/降解达到一定产量的 次数 总和来衡量需冷量的高低[C P(ChillPortion)]ꎬ在低温下ꎬ当果树体内中间产物合成/降解的循环次数要大于这个 次数总和 时ꎬ休眠结束ꎮ2㊀甜樱桃需冷量估算模型应用研究进展目前ꎬ上述需冷量估算模型在桃[7-12]㊁枣[13-16]㊁石榴[16]㊁无花果[16]㊁杏[17-19]㊁李[20]㊁葡萄[21ꎬ22]㊁蓝莓[23]㊁猕猴桃[24ꎬ25]㊁越橘[26]㊁梨[27-29]㊁树莓[30]㊁核桃[31]㊁开心果[32]上的应用研究均有报道ꎬ但在甜樱桃上的应用研究相对较少ꎬ使用的模型也主要是ɤ7.2ħ模型㊁0~7.2ħ模型和Utah模型ꎬ而有关动力学模型在甜樱桃需冷量研究上的851㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第54卷㊀文献报道还很少[33]ꎮ2.1㊀甜樱桃需冷量估算模型应用概述Alburquerque等[34]研究认为用Utah模型和动力学模型估算西班牙7个主栽甜樱桃品种需冷量的效果优于ɤ7.2ħ模型ꎮ刘晓娟[35]研究表明ꎬ0~7.2ħ模型和0~9.8ħ模型能够较好地反映天水地区乌克兰系列5个甜樱桃品种的需冷量ꎮ宋永宏等[36]对不同需冷量模型在9个甜樱桃主栽品种上的应用研究表明ꎬ0~7.2ħ模型估算的需冷量数值年际间差异最小ꎬUtah模型估算的数值年际间差异次之ꎬɤ7.2ħ模型估算的数值年际间差异最大ꎮ刘聪利等[37]对66个甜樱桃品种的研究认为ꎬ0~7.2ħ模型广泛适合甜樱桃需冷量的测定ꎬ而Utah模型不能有效预测冬季比较温暖或比较寒冷地区甜樱桃的需冷量ꎬ数值往往较小或是负值ꎮ郭天发等[12]认为动力学模型与0~7.2ħ模型和Utah模型估算的需冷量变化趋势相同ꎬ用来评价郑州地区桃需冷量可行ꎮ孙菀霞等[38]对长三角地区低温特征及其对甜樱桃需冷量影响的研究表明ꎬ应用0~7.2ħ模型㊁Utah模型和动力学模型估计的低温累积结果存在较大差异ꎮ肖钧等[39]应用0~7.2ħ模型㊁Utah模型和动力学模型对六盘水市樱桃种植区需冷量进行测定ꎬ结果表明各个模型年度间需冷量积累变异系数最小的为动力学模型ꎬUtah模型和0~7.2ħ模型次之ꎮ综上可见ꎬ现有研究主要集中在应用需冷量模型测定不同品种的需冷量上ꎬ但针对同一品种应用不同估算模型研究其在不同生态区域需冷量变化的报道还很少ꎮ2.2㊀基于文献资源的樱桃需冷量估算模型应用效果影响因素分析从上述分析可见ꎬ评价的模型㊁生态区和气候等因素差异都可能影响研究结果ꎬ且由于估算方法的不同ꎬ目前应用的植物需冷量并不是一个常数ꎬ其数值因选用的估算模型㊁确定的休眠起始时间等的不同而变化ꎮ因此ꎬ我们基于近年来中国知网数据库中收录的甜樱桃需冷量相关文献ꎬ对影响甜樱桃需冷量估算模型应用效果的因素进行了分析总结ꎮ2.2.1㊀基因型㊀物种的需冷量主要是由其基因型决定的ꎮ刘聪利等[37]用0~7.2ħ模型对郑州地区66个樱桃品种进行需冷量估算ꎬ得出58个品种需冷量值介于573~716hꎬ5个品种需冷量值ȡ740hꎬ3个品种需冷量值ɤ549hꎮ陈晓丹等[40]对不同地区的64个樱桃品种进行需冷量估算ꎬ得出需冷量值介于400~800h的品种有42个ꎬ介于800~1000h的品种有14个ꎬ介于200~400h㊁1000~1200h及>1200h的品种13个ꎮ应用同一模型估算出的不同樱桃品种需冷量不同ꎬ说明基因型对需冷量值起决定作用ꎮ2.2.2㊀生态环境㊀甜樱桃品种的需冷量值除受基因型因素影响外ꎬ还与地域因素有关ꎮAl ̄burquerque等[34]发现不同地区的需冷量与海拔呈正相关性ꎮ谭钺[41]㊁李淑珍[42]等对我国不同地区不同果树需冷量及扣棚时间的研究表明ꎬ不同地区果树需冷量受当地气候背景影响而不同ꎬ扣棚时间不统一ꎮ我们基于文献数据的统计结果表明ꎬ应用Utah模型估算的同一樱桃品种在山西晋中和山东泰安两个生态区的需冷量差异较大ꎬ变异系数为9.73%~23.98%ꎬ那翁㊁红灯㊁大紫3个品种的变异系数均超过15%(表2)ꎻ应用0~7.2ħ模型估算的同一樱桃品种在3个地区的需冷量也存在较大差异ꎬ4个品种的变异系数均超过20%ꎬ尤其品种先锋的变异系数高达38.63%(表3)ꎮ可见ꎬ樱桃品种的需冷量受生态环境影响明显ꎬ可能是由植株对地区的生态适应性所致ꎮ㊀㊀表2㊀用Utah模型估算的不同生态区樱桃需冷量差异分析樱桃品种地区需冷量(C.U)标准偏差变异系数(%)参考文献早红宝石山西晋中76182.09.73[36]山东泰安925[43]那翁山西晋中748236.023.98[36]山东泰安1220[43]红灯山西晋中809195.519.46[36]山东泰安1200[43]大紫山西晋中761194.520.36[36]山东泰安1150[43]㊀㊀注:需冷量单位C.U指自然休眠结束时积累的冷温单位(chillunit)ꎮ2.2.3㊀模型选择㊀由于对落叶果树打破休眠进程中的生理和遗传特性机制研究尚不充分ꎬ现有951㊀第12期㊀㊀㊀㊀㊀崔龙ꎬ等:我国甜樱桃需冷量估算模型应用与效果研究进展的需冷量模型都是以田间经验观察为基础建立的ꎬ但并不通用ꎬ模型间的转换因子变化也很大[31ꎬ33]ꎮ目前ꎬ研究者还未找到统一有效的适合不同地区不同品种的需冷量估算模型ꎮ㊀㊀表3㊀用0~7.2ħ模型估算的不同生态区樱桃需冷量差异分析品种地区需冷量(h)标准差变异系数(%)参考文献河南郑州589283.038.63[37]先锋山西晋中481[36]四川绵阳1128[44]河南郑州516155.126.80[37]雷尼尔山西晋中428[36]四川绵阳792[44]河南郑州583202.329.81[37]红灯山西晋中493[36]四川绵阳960[44]河南郑州790132.321.11[37]大紫山西晋中466[36]四川绵阳624[44]㊀㊀表4是基于文献数据统计的应用ɤ7.2ħ模型㊁0~7.2ħ模型和Utah模型计算的不同生态区红灯和大紫两个樱桃品种需冷量值ꎬ同时运用隶属函数法[45]计算出不同模型和生态区域下樱桃需冷量的隶属函数值ꎬ以综合评判生态区域和估算模型对需冷量的影响ꎬ结果(表5)表明ꎬ3个需冷量估算模型中ꎬ0~7.2ħ模型的隶属函数值最高ꎬ说明用其估算需冷量较其他两种模型更为适宜ꎻ两个樱桃品种在不同生态区的需冷量隶属函数平均值为0.602427和0.504115ꎬ均高于不同模型需冷量估算值0.528252和0.449115ꎬ因此推断生态区域对樱桃需冷量的影响大于所用估算模型的影响ꎮ㊀㊀表4㊀不同地区不同估算模型对需冷量的影响品种地区Utah模(C.U)0~7.2ħ模型(h)ɤ7.2ħ模型(h)参考文献红灯河南郑州583[37]四川绵阳960[44]山西晋中8094931254[36]山东泰安1200[43]变异系数27.5236.51大紫河南郑州790[37]四川绵阳624[44]山西晋中761466918[36]山东泰安1150[43]变异系数28.7925.85㊀㊀表5㊀基于隶属函数法的不同地区和估算模型对甜樱桃需冷量的影响项目隶属函数值红灯大紫参考文献ɤ7.2ħ模型(h)00[36]0~7.2ħ模型(h)11[36]Utah模型(C.U)0.5847570.347345[36]平均0.5282520.449115河南郑州0.8072810[37]四川绵阳00.512346[44]山西晋中11[36]平均0.6024270.504115㊀㊀综合上述分析ꎬ0~7.2ħ模型在国内甜樱桃需冷量估算上应用较广ꎬ在同一地区估算的需冷量累积年际间变异小于ɤ7.2ħ模型ꎮUtah模型明确界定了高温对需冷量累积的负效应ꎬ但因其计算比较复杂ꎬ目前应用不多ꎮ动力学模型在甜樱桃需冷量累积估算上的应用研究文献较少[46]ꎬ还需对其稳定性及适应性进行进一步研究验证ꎮ3㊀展望由于甜樱桃深受消费者喜爱且经济价值较高ꎬ种植范围已经扩展到我国南方次适宜栽培区和北方寒冷地区ꎮ需冷量作为甜樱桃品种引进和推广的重要参考依据ꎬ是能否引种成功的关键因素之一ꎮ但因所选用的需冷量估算模型主要是物候学模型ꎬ而非生态生理学模型ꎬ准确性受生态环境影响较大[46]ꎮ虽然需冷量估算模型已较为丰富ꎬ但尚未有一种模型可以适用于任何地区和物种ꎮ0~7.2ħ模型忽略了低于0ħ和高于7.2ħ的温度相互抵消作用对休眠的影响ꎮUtah模型虽然应用较多ꎬ对低温转换的范围分的较细ꎬ但对低于0ħ和高于18ħ的相互抵消作用没有做出界定ꎮ因此在选用适宜需冷量估算模型时ꎬ必须结合该地区物候学数据ꎬ同一品种不同地区的需冷量结果不能通用ꎮ今后可在优化需冷量估算模型方面进行深入研究ꎬ以期开发出通用性良好的估算模型ꎮ同一樱桃品种只有一个准确的休眠时间ꎬ因各个模型的休眠起始时间统计方法不同ꎬ所估算出的需冷量数值也不同ꎮ在选用适宜估算模型的同时ꎬ对甜樱桃低温积累的起点和终点进行研究ꎬ如通过规范测定低温积累起点的材料选择㊁处理061㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第54卷㊀方法㊁打破休眠的分级标准等实验流程ꎬ以使测出的需冷量值可为相同气候区域引种试栽提供参考依据ꎮ另外ꎬ甜樱桃树体需冷量累积过程中相关的生物学机制研究还较少ꎬ通过研究环境因子对休眠期树体相关基因的表达㊁树体内部的生理代谢及外部信号等的影响ꎬ解析甜樱桃休眠的分子生物学及生理代谢机制ꎬ以期为选用和开发适宜不同地域及气候的估算模型提供数据支撑ꎮ参㊀考㊀文㊀献:[1]㊀潘凤荣ꎬ郑玮.中国甜樱桃设施栽培历程㊁存在问题及发展建议[J].落叶果树ꎬ2019ꎬ51(5):1-4.[2]㊀潘凤荣ꎬ唐福会.甜樱桃高产优质栽培[M].辽宁:辽宁科学技术出版社ꎬ2010.[3]㊀朱运钦ꎬ乔改梅ꎬ王子崇ꎬ等.15个葡萄品种需冷量的研究[J].中国果树ꎬ2008(6):16-18.[4]㊀WeibergerJH.Chillingrequirementsofpeachvarieties[J].Pro ̄ceedingsoftheAmericanSocietyforHorticulturalScienceꎬ1950ꎬ56:122-128.[5]㊀EggertFP.AstudyofrestinseveralvarietiesofappleandinotherfruitspeciesgrowninNewYorkState[J].ProceedingsoftheAmericanSocietyforHorticulturalScienceꎬ1951ꎬ57:169-178.[6]㊀RichardsonEAꎬSeeleySDꎬWalkerDR.Amodelforestima ̄tingthecompletionofrestfor Redhaven and Elbertapeachtrees[J].HorticulturalScienceꎬ1974ꎬ9(4):331-332.[7]㊀ErezAꎬFishmanSꎬLinsley ̄NoakesGCꎬetal.Thedynamicmodelforrestcompletioninpeachbuds[J].ActaHorticulturalScienceꎬ1990ꎬ276:165-174.[8]㊀Linsley ̄NoakesGCꎬAllanP.Comparisonoftwomodelsforthepredictionofrestcompletioninpeaches[J].Sci.Hortic.ꎬ1994ꎬ59(2):107-113.[9]㊀王力荣ꎬ朱更瑞ꎬ方伟超ꎬ等.桃品种需冷量评价模式的探讨[J].园艺学报ꎬ2003(4):379-383.[10]张明昊ꎬ严娟ꎬ蔡志翔ꎬ等.103份桃种质在南京地区的需冷量和需热量研究[J].果树学报ꎬ2021ꎬ38(1):29-39. [11]刘芸ꎬ姜新ꎬ陆玉英ꎬ等.桃树需冷量研究进展[J].中国农学通报ꎬ2019ꎬ35(21):61-65.[12]郭天发ꎬ陈昌文ꎬ李勇ꎬ等.桃新品种需冷量和需热量评价及动力学模型分析[J].新疆农业科学ꎬ2021ꎬ58(10):1821-1828.[13]梁文锋ꎬ徐崇志ꎬ王振磊ꎬ等.不同需冷量枣品种休眠期间的生理变化[J].干旱区资源与环境ꎬ2020ꎬ34(6):187-193.[14]韩蓓蓓ꎬ李东ꎬ雪婷ꎬ等.大荔冬枣的自然休眠和需冷量初探[J].陕西气象ꎬ2020(1):44-46.[15]张川疆ꎬ陈霞ꎬ林敏娟.不同枣品种需冷量研究[J].黑龙江农业科学ꎬ2019(8):88-92.[16]崔萍ꎬ曹尚银ꎬ姜建福.枣㊁石榴㊁无花果需冷量测定试验初报[J].经济林研究ꎬ2009ꎬ27(1):91-93.[17]逯昀ꎬ汤爱勤.豫东地区设施杏需冷量监测及扣棚升温时期研究[J].中国南方果树ꎬ2017ꎬ46(3):113-115. [18]刘桂彬.沧州地区杏树适宜需冷量模型比较[J].现代农业科技ꎬ2010(19):105ꎬ111.[19]RuizDꎬCampoyAJꎬEgeaJ.Chillingandheatrequirementsofapricotcultivarsforflowering[J].EnvironmentalandExperi ̄mentalBotanyꎬ2007ꎬ61(3):254-263.[20]高志红ꎬ张君毅ꎬ乔玉山ꎬ等.桃和李品种需冷量研究[J].中国果树ꎬ2004(3):20-23.[21]郭松涛ꎬ张亚红ꎬ李琴ꎬ等.宁夏地区设施葡萄需冷量和需热量研究[J].果树学报ꎬ2020ꎬ37(7):997-1007. [22]王海波ꎬ刘凤之ꎬ韩晓ꎬ等.葡萄需冷量和需热量估算模型及设施促早栽培品种筛选[J].农业工程学报ꎬ2017ꎬ33(17):187-193.[23]杨玉春ꎬ魏鑫ꎬ孙斌ꎬ等.蓝莓不同品种低温需冷量研究分析[J].农业科技通讯ꎬ2020(1):178-181.[24]赵婷婷.中华猕猴桃栽培品种需冷量研究[D].武汉:中国科学院研究生院(武汉植物园)ꎬ2016.[25]赵婷婷ꎬ韩飞ꎬ陈美艳ꎬ等.基于3种模型的猕猴桃重要栽培品种需冷量研究[J].中国果树ꎬ2018(6):36-39. [26]于斯嘉.越橘需冷量及休眠期生理生化特性的研究[D].长春:吉林农业大学ꎬ2014.[27]李先明ꎬ秦仲麒ꎬ涂俊凡ꎬ等.梨品种需冷量评价模式[J].西北农业学报ꎬ2013ꎬ22(5):68-71.[28]封雷.南京地区梨需冷量及设施栽培关键技术研究[D].南京:南京农业大学ꎬ2013.[29]韩浩章.苏南地区主要落叶果树的需冷量及休眠解除生理机制的研究[D].南京:南京农业大学ꎬ2004.[30]徐红艳.夏果型树莓的需冷量㊁破除休眠方法及促成栽培表现研究[D].南京:南京农业大学ꎬ2013.[31]LuedelingEꎬZhangMꎬMcGranahanGꎬetal.Validationofwinterchillmodelsusinghistoricrecordsofwalnutphenology[J].AgriculturalandForestMeteorologyꎬ2009ꎬ149(11):1854-1864.[32]ZhangJꎬTaylorC.Thedynamicmodelprovidesthebestde ̄scriptionofthechillprocesson Sirora pistachiotreesinAus ̄tralia[J].HortScienceꎬ2011ꎬ46(3):420-425.[33]KwonJꎬNamEꎬYunSꎬetal.Chillingandheatrequirementofpeachcultivarsandchangesinchillingaccumulationspec ̄trumsbasedon100 ̄yearrecordsinRepublicofKorea[J].Ag ̄riculturalandForestMeteorologyꎬ2020ꎬ288/289:108009. [34]AlburquerqueNꎬGarcía ̄MontielFꎬCarrilloAꎬetal.Chillingandheatrequirementsofsweetcherrycultivarsandtherelation ̄shipbetweenaltitudeandtheprobabilityofsatisfyingthechillrequirements[J].EnvironmentalandExperimentalBotanyꎬ2008ꎬ64(2):162-170.161㊀第12期㊀㊀㊀㊀㊀崔龙ꎬ等:我国甜樱桃需冷量估算模型应用与效果研究进展[35]刘晓娟.乌克兰大樱桃系列品种需冷量研究[J].甘肃农业ꎬ2009(12):104-105.[36]宋永宏ꎬ戴桂林ꎬ聂国伟ꎬ等.山西晋中甜樱桃主栽品种需冷量研究[J].中国果树ꎬ2016(6):12-15ꎬ20.[37]刘聪利ꎬ赵改荣ꎬ李明ꎬ等.66个甜樱桃品种需冷量的评价与聚类分析[J].果树学报ꎬ2017ꎬ34(4):464-472. [38]孙菀霞ꎬ刘勋菊ꎬ王丽ꎬ等.长三角地区低温特征及其对甜樱桃蓄冷量的影响[J].果树学报ꎬ2021ꎬ38(11):1900-1910.[39]肖钧ꎬ张敏ꎬ吴亚维ꎬ等.六盘水市甜樱桃主要种植区蓄冷量和蓄热量分析[J].黑龙江农业科学ꎬ2019(9):54-58. [40]陈晓丹ꎬ王磊ꎬ黄嘉赟ꎬ等.甜樱桃不同品种需冷量评估初探[J].中国南方果树ꎬ2017ꎬ46(3):109-112.[41]谭钺.设施桃低温需求量与需热量关系机制的初步研究[D].泰安:山东农业大学ꎬ2010.[42]李淑珍ꎬ赵文东ꎬ韩凤珠ꎬ等.不同地区设施果树的扣棚及升温时间[J].北方果树ꎬ2005(6):39-40.[43]陈晓丹.甜樱桃品种需冷量评估及休眠相关MADS-box基因研究[D].上海:上海交通大学ꎬ2017.[44]刘仁道ꎬ刘建军.甜樱桃不同品种需冷量研究[J].北方园艺ꎬ2009(2):84-85.[45]陈蕾ꎬ张艳波ꎬ崔龙ꎬ等.基于隶属函数法的李涂白防寒措施筛选[J].分子植物育种ꎬ2020ꎬ18(6):2070-2076. [46]HänninenH.Effectsofclimaticchangeontreesfromcoolandtemperateregionsanecophysiologicalapproachtomodelingofbudburstphenology[J].CanadianJournalofBotanyꎬ2011ꎬ73(2):183-199.261㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀山东农业科学㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀㊀第54卷㊀。

吉林省科学技术厅关于批准建设首批吉林省优质道地
药材科技示范基地的通知
文章属性
•【制定机关】吉林省科学技术厅
•【公布日期】2021.09.02
•【字号】吉科发医〔2021〕160号
•【施行日期】2021.09.02
•【效力等级】地方规范性文件
•【时效性】现行有效
•【主题分类】药政管理
正文
关于批准建设首批吉林省优质道地药材科技示范基地的通知
吉科发医〔2021〕160号各市(州)科技局:
为贯彻落实《全国中医药发展战略规划纲要(2016-2030年)》《全国道地药材生产基地建设规划(2018-2025年)》《中共吉林省委吉林省人民政府关于促进中医药传承创新高质量发展的实施意见》精神,进一步加快落实省政府关于“打造吉药整体道地药材形象,加强种植(养殖)示范基地建设和质量监管”要求,充分发挥科技支撑引领作用,加快我省道地药材高质量可持续发展,省科技厅启动了首批“吉林省优质道地药材科技示范基地”建设工作。
经各市(州)推荐、专家评审、现场考察,现批准吉林省北药药材加工有限公司等11个基地为首批建设的吉林省优质道地药材科技示范基地(名单见附件)。
各优质道地药材科技示范基地建设单位,要坚持保护与发展,做好优质道地药材种植养殖,围绕优质道地药材关键技术问题,创新一批适宜单元技术,集成示范推广中药材新品种、新技术,切实发挥好示范带动和辐射推广作用。
各市(州)科技管理部门要围绕中药材资源保护、品种培育繁育、规范化种植养殖等方面,加强
对优质道地药材科技示范基地建设政策引导和扶持,共同推进道地药材规范化、标准化、规模化与品牌化发展。
附件:首批吉林省优质道地药材科技示范基地名单
吉林省科学技术厅
2021年9月2日。
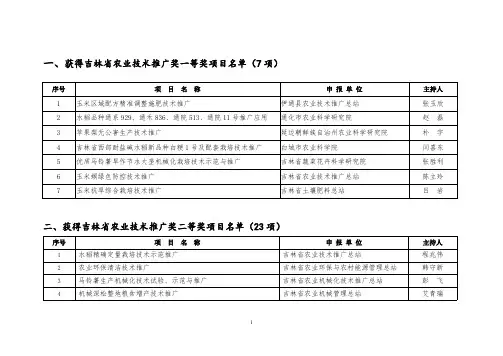
一、获得吉林省农业技术推广奖一等奖项目名单(7项)序号项目名称申报单位主持人1玉米区域配方精准调整施肥技术推广伊通县农业技术推广总站张玉欣2水稻品种通系929、通禾836、通院513、通院11号推广应用通化市农业科学研究院赵磊3苹果梨无公害生产技术推广延边朝鲜族自治州农业科学研究院朴宇4吉林省西部耐盐碱水稻新品种白粳1号及配套栽培技术推广白城市农业科学院闫喜东5优质马铃薯旱作节水大垄机械化栽培技术示范与推广吉林省蔬菜花卉科学研究院张胜利6玉米螟绿色防控技术推广吉林省农业技术推广总站陈立玲7玉米抗旱综合栽培技术推广吉林省土壤肥料总站吕岩二、获得吉林省农业技术推广奖二等奖项目名单(23项)序号项目名称申报单位主持人1水稻精确定量栽培技术示范推广吉林省农业技术推广总站程兆伟2农业环保清洁技术推广吉林省农业环保与农村能源管理总站韩守新3马铃薯生产机械化技术试验、示范与推广吉林省农业机械化技术推广总站彭飞4机械深松整地粮食增产技术推广吉林省农业机械管理总站艾青瑞5玉米高光效休耕轮作高产栽培技术推广九台市农业技术推广中心周世艳6农村户用沼气综合建设技术推广九台市农村能源环境保护办公室韩景林7德惠市郭家镇四千亩葡萄无公害标准化生产技术推广项目德惠市农产品质量监督检验管理中心李勇8双膜梁日光温室建设和后墙立体栽培技术组合长春市双阳区植物保护植物检疫站衣绍清9水稻机械化育插秧技术推广吉林市农业机械化技术推广中心洪杰10百万亩玉米田施用中微量元素技术推广应用磐石市农业技术推广站邹明辉11滑菇袋式代料栽培技术推广磐石市园艺特产技术推广站王丽清12水稻纹枯病综合防治技术推广吉林市植物保护植物检疫站孙丰年13机械深松蓄水与节水灌溉旱作栽培技术推广四平市农业机械化技术推广站曲甲民14玉米秸秆覆盖保护性耕作技术推广梨树县农业技术推广总站赵晓霞15双辽市花生大拢三行高产高效栽培技术推广双辽市农业技术推广中心张建华16玉米密植高产栽培技术研究与推广辽源市农业技术推广总站郭文景17通化市东昌区无公害地栽黑木耳栽培技术推广东昌区绿色食品办公室姜文辉18大棚黄瓜新品种优质高产栽培技术推广白城市洮北区蔬菜技术推广总站刘汉飞19向日葵杂交种及配套标准化栽培技术推广白城市种子管理站孙昕20西瓜套种杂交葵花高产栽培技术推广通榆县农民科技教育培训中心徐明慧21前郭县节水灌溉施肥技术推广前郭县农业技术推广中心张立明22宁江区测土配方施肥技术推广松原市宁江区农业技术推广中心王振23优质大豆“延农8号、11号”及配套栽培技术推广延边朝鲜族自治州农业科学研究院黄初女三、获得吉林省农业技术推广奖三等奖项目名单(26项)序号项目名称申报单位主持人1爆玉米高产高效综合技术推广长春净月经济开发区玉潭镇农业服务站刘玉敏2农安县玉米“一增四改”综合配套技术集成与推广农安县农业技术推广中心郝彦德3水稻生产全程机械化技术推广永吉县农业机械化技术推广站戚国富4玉米保护性耕作机械化技术推广磐石市农业机械技术推广站辛太国5综合农艺措施治理“三湖区”耕地面源污染技术推广桦甸市农业技术推广中心曹文明6防病除虫检测三位一体综合配套无公害蔬菜生产技术推广吉林市农业环保监测站李卫国7伊通县5000公顷无公害水稻栽培防治技术推广四平市农业技术推广中心郭星8板蓝根高效栽培示范双辽市农业技术推广中心刘书佳9东辽县水稻二化螟综合防治技术推广东辽县农业技术推广总站朱红军10东丰县6万公顷玉米增密高产技术推广东丰县农业技术推广总站孔庆涛11梅河口市沃土培肥工程推广梅河口市农业技术推广总站邹玉生12玉米高产节本增效综合配套栽培技术推广柳河县农业技术推广总站祝宏13等离子体种子处理技术推广通化县农业机械化技术推广站李威14玉米高产高效综合配套栽培技术推广抚松县农业技术推广总站刘美良155000亩晒红烟综合高产栽培技术推广临江市农业技术推广中心金勇男16龙胆草人工栽培技术推广靖宇县农业技术推广中心郑艳梅17花生新品种白院花3号丰产栽培技术推广吉林省白城市农业科学院王瑛霞18水稻综合节水高产配套栽培技术推广白城市洮北区水稻办公室李莉梅19花生标准化高产栽培技术推广白城市农业技术推广总站唐晓清20洮南市无公害红干椒标准化栽培技术推广洮南市农业技术推广中心杨瑞红21三十万亩白根萝卜优质高产综合配套栽培技术推广长岭县农业技术推广中心夏玉春22无公害番茄日光温室栽培技术推广松原市宁江区绿色食品开发办公室李国23水稻机械化收获推广延边州农机科技推广站杨占峰24玉米垄侧减免耕保墒栽培技术推广延边州农业技术推广站崔东成25沼液在水稻生产上应用技术推广和龙市农业技术推广中心崔日成26脱毒马铃薯标准化栽培技术推广敦化市农业技术推广中心吴桐。
一是提升了金融机构服务水平。
研发和试点推广“农地贷”产品,既是贯彻落实中央关于农村土地制度改革精神的具体行动,更是助推农业现代化、规模化发展的责任所在。
这项工作不仅促进农村土地有序流转,农业适度规模经营,而且破解了农村金融瓶颈,提升了农行支农水平,促进了农民增收致富。
二是降低了农村融资成本。
过去农民经常被“贷款难、担保难”所困扰。
一方面是缺钱发展生产,另一方面是无抵押担保,难以取得贷款。
而农村土地经营权、宅基地及房屋等农民的核心资产难以抵押贷款。
“农地贷”产品的创新推出,在办理抵押担保环节实行零费用,贷款利率实行保本微利,最大限度让利于民,而且还能享受政府财政贴息补助,极大地降低了种粮大户的融资成本。
三是提高了农村耕地效率。
通过“农地贷”产品的支持,农民可以规模经营、平整土地、打破户与户之间的土地隔界,扩大水田田块等措施,土地面积平均增加5%以上;实行规模化经营提高机械化作业率,粮食产量增加15%左右。
四是实现了农民增收致富。
一是散户增效。
延吉市土地流转价格从过去每公顷2000~3000元提高到近5000元,有的达到6000元,散户流出土地收益增加;同时从少量土地中解放出来,外出打工挣取劳务,收益大大增加;二是大户增收。
贷款为新型规模农业经营主体提供了稳定的、低成本的资金来源,促进了生产经营规模的逐步扩大,实现大型农机化集中连片作业,规模效应开始显现,盈利能力大幅提高。
五是促进了土地资源的有效转化。
农村土地经营权抵押贷款的推出,不仅解决了农户和新型农业经营主体贷款难、融资利率高的难题,而且促进了农村土地资源变资产、资产变资金的有效转化。
同时,通过银政等多方沟通和深度合作,为延边州申报开展农业部土地承包经营权流转规范化管理服务试点,积累了丰富的经验,提供了有力支撑。
5试点工作存在的主要问题一是法律法规修改跟进滞后于实际发展需要。
目前的法律法规对于“稳定承包权、放活经营权”政策体现不明显,仍是基于原来土地承包经营权一体为基础制定的,因此对于用土地经营权抵押贷款缺乏明确的法理依据。
吉林省科学技术厅关于2020年度吉林省科学技术奖励的决定文章属性•【制定机关】吉林省科学技术厅•【公布日期】2020.11.16•【字号】吉科发奖〔2020〕231号•【施行日期】2020.11.16•【效力等级】地方规范性文件•【时效性】现行有效•【主题分类】科技奖励正文吉林省科技厅关于2020年度吉林省科学技术奖励的决定各市(州)人民政府,长白山管委会,长春新区、中韩(长春)国际合作示范区管委会,各县(市)人民政府,各有关部门(单位):为深入贯彻落实习近平新时代中国特色社会主义思想,扎实推进创新驱动发展战略的实施,进一步调动和激发全省科技人员的创新创造活力,经省政府同意,决定对在吉林全面振兴全方位振兴过程中体现新担当、实现新突破、展现新作为的科学技术人员和组织给予表彰。
根据《吉林省科学技术奖励办法》规定,通过专家评审、复审和省科学技术奖励委员会全体会议审议,决定授予王家骐院士、王立春研究员“吉林省科学技术特殊贡献奖”。
授予《新型纳米孔和纳米结构材料的可控构筑及其在能源与环境中的应用》等11个项目吉林省自然科学奖一等奖;《云计算下大数据高效处理的关键技术研究与应用》等26个项目吉林省自然科学奖二等奖;《大豆高产高抗的生理机制解析》等24个项目吉林省自然科学奖三等奖。
授予《数控机床系列关键功能部件可靠性试验技术与装备》等4个项目吉林省技术发明奖一等奖;《快响应、精细化废气再循环调控关键技术》吉林省技术发明奖二等奖;《基于空间波导调制电子束倍增的瞬态真空光电子器件研究》等4个项目吉林省技术发明奖三等奖。
授予《远紫外高灵敏度成像技术及航天应用》等21个项目吉林省科学技术进步奖一等奖;《混合动力DCT及其平台产品研发与产业化》等83个项目吉林省科学技术进步奖二等奖;《生物可降解聚酯改性填料的制备技术》等102个项目吉林省科学技术进步奖三等奖。
希望获奖者珍惜荣誉,再接再厉,坚持以习近平总书记视察吉林重要讲话重要指示精神为指导,全面落实“四个面向”科技创新要求,大力弘扬科学家精神,创造更多世界领先的科技成果。
吉林省科学技术厅、吉林省财政厅关于对2022年度吉林省科技发展计划拟支持项目进行公示的通知
文章属性
•【制定机关】吉林省科学技术厅,吉林省财政厅
•【公布日期】2022.07.07
•【字号】
•【施行日期】2022.07.07
•【效力等级】地方规范性文件
•【时效性】现行有效
•【主题分类】科技计划
正文
关于对2022年度吉林省科技发展计划拟支持项目进行公示
的通知
各有关单位:
根据《吉林省科技发展计划项目立项管理办法》和《吉林省科技创新专项资金管理办法》以及省科技厅、省财政厅相关要求,现将2022年度吉林省科技发展计划拟支持项目向社会公示,公示期为7月8日至7月14日。
如有异议,可在公示期内提出书面意见,并注明真实姓名、工作单位和联系电话。
综合业务咨询电话:*************,纪检监督联系电话:*************,相关业务处室联系电话详见附件,来信请至长春市民康路522号209室,吉林省科学技术厅发展规划处,邮编130041。
附件(略):2022年度吉林省科技发展计划项目安排情况表
吉林省科学技术厅吉林省财政厅
2022年7月7日。
34APICULTURE OF CHINA蜜蜂遗传育种新技术(一)——蜜蜂转基因与克隆薛运波│文 李志勇│图育微课堂种1.蜜蜂转基因技术蜜蜂转基因育种(transgenic breeding)是通过基因工程技术将外源目的基因导入受体细胞,整合到受体细胞基因组中,并使外源基因得到表达和遗传,以此获得新品种。
该技术的根本意义在于它克服固有的生殖隔离,实现物种间遗传物质的交换,在改良蜜蜂抗病和经济性状以及生物反应器利用等方面展示了良好的应用前景。
自从1982年成功研究出首例转基因果蝇以来,转基因昆虫的研究便引起科学家们极大的兴趣。
目前,已经获得成功的转基因昆虫有黑腹果蝇、海地果蝇、地中海实蝇、家蚕、蚊子等。
鉴于蜜蜂特殊的社会行为和级型分化现象,有学者认为以下两种方法是蜜蜂转基因较好的途径:一是以蜜蜂人工授精技术为基础的精子介导转基因法,二是蜜蜂卵或幼虫的转基因操作与蜜蜂人工孵育技术相结合的方法。
(1)精子介导转基因法1971年,Brackett 等发现精子细胞具有自发地吸收外源DNA,并可在受精的过程中将外源DNA 携带进卵内。
用精子作为转移外源DNA 的载体,将DNA 溶液和动物精子共浴,精子能主动吸附外源DNA,利用这一点可以很容易地将外源DNA 导入精子细胞或结合于精子细胞表面。
再通过人工授精,将外源DNA 带入受精卵进而发育成个体(图1),从而生产转基因动物。
利用精子介导的转基因蜜蜂的研究也有成功报道。
1991年,Atkinson 等首次将蜜蜂精子与外源DNA 进行共培养,证实蜜蜂精子也能够吸收其他动物的DNA。
2000年,Robinson 等研究精子介导转染法用于蜜蜂转基因的可行性。
结果表明,蜜蜂精子能将外源线性质粒DNA 携带进入卵子,外源DNA 能在受体蜜蜂个体中存在数月之久,能在幼虫期进行表达,至少能稳定遗传三代。
尽管没有找到外源DNA 整合到蜜蜂基因组上的证据,但该研究证明蜜蜂精子同样具有携带外源DNA 进入卵子的能力。
附件:吉林省肉牛产业化发展重大科技专项申报指南实施“秸秆变肉”暨千万头肉牛建设工程,是我省打造万亿级农产品加工业和食品产业的必然要求,是构筑吉林农业发展新优势的必然选择,是促进农业高质高效、农民富裕富足的重要途径。
在现有资源条件下,我省肉牛生产水平进步明显,但仍存在新品种培育缓慢、秸秆等粗饲料资源利用不充分、肉牛产品多元化开发不足等技术性制约难题。
本专项旨在围绕肉牛种业创新、高效繁育、粗饲料开发利用、产品精深加工和疫病防治等关键共性技术开展攻关,为千万头肉牛工程建设提供有力的科技支撑。
专项遵循“链条部署,一体化实施”的原则,以课题为单元组织申报,鼓励产学研协同攻关,共设置7个课题,执行期3年。
预计资助总额度1500万元。
每个课题支持1-2项,资助额度不超过150万/项(分两批次拨付)。
其中,企业牵头承担的项目、市县所属单位申报的项目,资助比例最高不超过项目预算总额的50%;高校、科研院所等事业单位联合企业共同申报的项目,资助比例最高不超过项目预算总额的70%。
项目所需其余资金由项目单位自行筹措足额落实。
课题1:肉牛新品种(系)培育研究重点内容:以新品种(新品系、新种群)培育为需求导向,立足我省肉牛主导品种群体优势和地方品种资源特色优势,围绕地方品种肉质好、抗病抗逆性强等特点,以传统育种技术为基础,开展主导品种区域间联合育种、地方特色多品种联合育种,通过研发并熟化表型性状精准鉴定技术、分子标记辅助选择技术、全基因组选择技术、早期精确选种技术、多品种联合遗传评估等技术,系统评价挖掘地方品种牛优质、抗逆等特色性状关键基因和遗传基础,持续加强新品种和本地品种选育,不断提高肉牛群体的生产性能和核心种源自给率,开展优质、高产、抗逆等性状突出的特色新品种培育和种群扩繁。
考核指标:构建品种培育关键技术体系2-3个,培育优质、高产、抗逆性强等性状突出的优质肉牛品系(品种)2-3个,高档肉产量、日增重、饲料利用率等关键指标提高5-10%。