第六章线粒体
- 格式:ppt
- 大小:7.20 MB
- 文档页数:15

第六章线粒体一、名词解释1、ATP synthetase(ATPase) ATP合成酶2、biological oxidation 生物氧化3、chemiosmotic coupling hypothesis 化学渗透学说4、cristae 嵴5、elementary particle 基本颗粒6、leader peptide(sequence) 导肽7、mitochondria 线粒体8、oxidative phosphorylation 氧化磷酸化9、respiratory chain 呼吸链10、semiautonomous organelle 半自主性细胞器11、submitochondrial particle 亚线粒体小泡二、是非判断1、1890年Altmann在肝细胞内首次发现命名为bioblast,他认为细胞中这类颗粒与细菌相似,可能是共生于细胞内能够独立自主生活的有机体。
的功能是质子传递的通道并产生ATP。
2、F3、动物细胞所含的线粒体一般较植物细胞少。
4、动物细胞嵴排列成小管状,植物及低等动物排列成板层。
5、线粒体外膜可允许1万Dalton以下的分子进出。
6、线粒体疾病主要分为遗传性和获得性疾病两大类。
7、线粒体释放的cytC不参与细胞凋亡。
8、动物细胞线粒体内的核糖体小些约55S,植物细胞和酵母菌的大些接近78S。
期进行。
9、植物细胞MtDNA较动物为大,一般其复制是在G210、线粒体的遗传密码与核基因的遗传密码稍有不同,不同物种的线粒体遗传密码都是相同的。
11、导肽运送的蛋白质都是分泌性的。
12、线粒体内膜和外膜在化学组成上的区别是内膜的脂和蛋白质的比值不同,前者为1:1, 后者则为0.3:1。
13、导肽是引导新生肽进行转运和定位的一段序列,转运后不一定都要被切除。
14、线粒体基因表达过程中的密码系统与通用的密码系统完全一样。
15、线粒体一般以棒状和粒状为主,在一定条件下可以转变并且是可逆的。

第六章线粒体名词解释1、电子传递链electron-transport chain膜上一系列由电子载体组成的电子传递途径。
这些电子载体接受高能电子,并在传递过程中逐步降低电子的能量,最终将释放的能量用于合成ATP或以其他能量形式储存。
2、化学渗透学说chemiosmosis氧化磷酸化的耦联机制。
电子经电子传递链传递后,形成跨线粒体内膜的质子动力势,用以驱动ATP合成酶合成ATP。
3、结合变构模型binding change model利用质子动力势驱动ATP合成酶构象发生改变,将ADP和无机磷合成ATP的模型。
4、孔蛋白porin存在于线粒体和叶绿体外膜上的整合膜蛋白,形成非选择性的通道。
5、内共生学说endosysmbiont theory关于叶绿体和线粒体起源的假说,认为叶绿体和线粒体起源于被原始真核细胞吞噬的共生原核生物。
6、线粒体mitochondrion将储存在有机物中的能量通过氧化磷酸化过程形成ATP的细胞器。
线粒体是一种能量转换细胞器,还参与细胞凋亡等重要生理过程。
7、氧化磷酸化oxidative phosphorylation底物在氧化过程中产生高能电子,通过线粒体内膜电子传递链,将高能电子的能量释放出来转换成质子动力势进而合成ATP的过程。
8、ATP合酶ATP synthase位于线粒体内膜或叶绿体的类囊体膜上,通过氧化磷酸化或光合磷酸化催化ADP和无机磷合成ATP的酶,由F1头部和嵌入膜内的F0基部组成,也常见于细菌膜上。
9、线粒体膜间隙intermembrane space线粒体内膜和外膜之间的间隙,约6~8nm,其中充满无定形的液体,含有可溶性的酶、底物和辅助因子。
膜间隙的标志酶是腺苷酸激酶。
10、嵴cristae线粒体内膜向基质折褶形成的结构称作嵴(cristae), 嵴的形成使内膜的表面积大大增加。
11、电子载体electron carriers在电子传递过程中与释放的电子结合并将电子传递下去的物质称为电子载体。


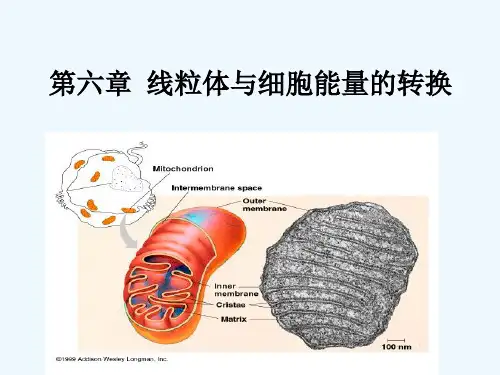


第六章线粒体与细胞的能量转换第一节线粒体的基本特征一、线粒体的形态、数量和结构(一)线粒体的形态、数量与细胞的类型和生理状态有关(细胞类型、生理状态、代谢需求)1.光镜下的线粒体成线状、粒状或杆状。
2.在低渗环境下,线粒体膨胀如泡状,在高渗环境下,线粒体又伸长为线状3.酸性时线粒体膨胀,碱性时线粒体为粒状(二)线粒体是由双层单位膜套叠而成的封闭性膜囊结构1.外膜是线粒体外层单位膜在组成上,外模的1/2为脂类,1/2位蛋白质,外膜上镶嵌的蛋白质包括多种转运蛋白,允许通过分子量在10000以下的物质(通透性大)2.内膜的内表面附着许多颗粒①内膜直接包围的空间称内腔,含有基质,也称基质腔;内膜与外膜之间的空间称为外腔,或膜间腔。
②嵴的形成大大扩大了内膜的面积,提高了内膜的代谢效率③内膜的化学组成中20%是脂类(心磷脂占20%),80%是蛋白质④内膜的通透性很小,但内膜有高度的选择通透性⑤基粒分为头部、柄部、基片三部分,由多种蛋白质亚基组成。
机理头部具有酶活性,能催化ADP磷酸化生成ATP,因此,基粒又称ATP合成酶或ATP合酶复合体3.内外膜相互接近所形成的转为接触点是物质转运到线粒体的临时性结构线粒体的内外膜上存在着一些内膜与外模相互接触的地方,在这些地方膜间隙变狭窄,称为转位接触点4.基质是氧化代谢的场所线粒体中催化三羧酸循环、脂肪酸氧化、氨基酸分解、蛋白质合成等有关的酶都在基质中,参与物质的代谢5.基粒的化学本质是ATP合成酶二、线粒体的化学组成1.线粒体的主要成分是蛋白质,且多数分布于内膜和基质,分为两类:可溶性蛋白和不可溶性蛋白或膜镶嵌酶蛋白(线粒体是细胞中含酶最多的细胞器)2.线粒体内外膜的标志酶分别是细胞色素氧化酶和单胺氧化酶等;基质和膜间腔的标志酶分别为苹果酸脱氢酶和腺苷酸激酶三.线粒体的遗传体系(一)线粒体DNA构成了线粒体基因组1.线粒体基因组序列(也称剑桥序列)共16569个碱基对,为一条裸露的,不与组蛋白结合的双链环状的DNA分子。

医学遗传学第六章线粒体遗传病线粒体基因组的结构特征:不与组蛋白结合,呈裸露闭环双链状。
分为重链H链和轻链L 链。
重链富含鸟嘌呤G。
轻链富含胞嘧啶C。
编码区:线粒体基因排列紧密,部分会重叠。
没有启动子和内含子。
有的基因没有完整的三连体终止密码。
基因间隔区短,只有87bp。
两条链都有编码功能。
非编码区(D环区/控制区):与线粒体复制及转录有关。
包括H链复制的起始点。
H、L链转录的启动子。
四个高度保守序列(可通过此结构判定物种)。
两个高变区。
编码区的保守序列包含37个基因:2个rRNA基因,22个tRNA基因以及是13个氧化磷酸化有关的蛋白质编码基因。
线粒体基因组的遗传特征:半自主性遗传密码和通用密码不同母系遗传:母亲将mtDNA传递给她的所有子女,但只有她的女儿们能将其mtDNA传递给下一代。
mtDNA的突变率极高:原因是--mtDNA的特殊结构(缺乏酪蛋白和其他DNA组结合蛋白的保护。
无DNA损伤的修复系统,没有内含子。
独特的复制方式和复制错配。
mtDNA处于高度氧化的环境。
(ROS和自由基导致氧化损伤或诱导突变。
)同质性与异质性:同质性:一个组织或细胞中所有的线粒体具有相同的基因组,全部为野生型或全部为突变型mtDNA。
异质性:一个细胞或组织具有突变型,也有野生型mtDNA,基金组不同。
异质性的发生率:中枢神经系统>肌肉组织>血液。
成人大于儿童。
阀值效应:突变的mtDNA达到一定的比例,超过阀值时能引起某些器官或组织功能异常,导致能量产生急剧下降。
对ATP依赖程度:中枢神经系统>骨骼肌>心脏>胰腺>肾脏>肝脏。
遗传瓶颈与复制分离:遗传瓶颈:卵母细胞中的线粒体数目从10万个锐减到少于100个的过程。
复制分离:细胞分裂时进行随机分配,导致mtDNA异质性变化的过程。
线粒体基因组突变类型:点突变:2/3的点突变发生在与线粒体内蛋白质翻译有关的tRNA和rRNA基因上,这些突变会导致tRNA和rRNA的结构异常,从而影响mtDNA编码的多肽链的翻译过程。


