田瑞军副教授南方科技大学化学系生物质谱与蛋白质组学课
- 格式:ppt
- 大小:213.50 KB
- 文档页数:3

Methods HEstaining,Massonstaining,immunohisto chemistry,qPCR,WesternblotandotherexperimentalmethodswereusedtoverifytheeffectofHDonintesti nalfibrosisinmiceandthepotentialmechanism.Re sults Invivoexperimentsshowedthatcomparedwiththenormalgroup,theDSS inducedintestinalfibrosismodelgrouphadshortenedcolon,increasedcolonhis topathologicalscore,increasedcollagenvolumefrac tion,andsignificantlyincreasedcollagenⅠexpres sion.Aftertreatmentwith4,10,and25mg·kg-1HD,comparedwiththemodelgroup,thecolonshorten ingofmicewasalleviated,andthehistopathologicalscore,collagenvolumefractionandtheexpressionofcollagenⅠdecreasedinadose dependentmanner.Comparedwiththemodelgroup,incolontissueandIEC 6cells,HDcoulddose dependentlyinhibittheproteinandmRNAlevelexpressionofEMT relatedfac torssuchasα SMA,Vimentin,N cadherin,andin creasetheexpressionofE cadherin.Conclusion HDcanimproveexperimentalintestinalfibrosisbyinhibi tingEMT.Keywords:p hydroxybenzaldehyde;intestinalfibro sis;inflammatoryboweldisease;collagendeposition;epithelial mesenchymaltransition;cadherin switch网络出版时间:2023-03-2015:54:20 网络出版地址:https://kns.cnki.net/kcms/detail/34.1086.R.20230319.2353.032.html◇肝脏药理学◇FATP5基因沉默抑制油酸诱导的HepG2细胞脂肪化及炎症作用王天琦1,孙文龙2,杨馥源3,李敬达4(1.长江大学农学院,湖北荆州 434000;2.山东理工大学生命科学学院,山东淄博 255000;3.长江大学医学院,湖北荆州 434000;4.长江大学生命科学学院,湖北荆州 434000)doi:10.12360/CPB202202066文献标志码:A文章编号:1001-1978(2023)04-0692-08中国图书分类号:R342 2;R344 7;R364 5;R392 11;R575 5摘要:目的 明确FATP5作为脂肪酸转运蛋白家族成员,其基因沉默对肝脏细胞脂肪化及炎症作用的影响,并初步探究该影响可能的机制。

科研Mol.Plant:拟南芥蛋白质组图谱的重塑与蛋白质在发育和免疫中共同调节编译:东方不赢,编辑:Emma、江舜尧。
原创微文,欢迎转发转载。
导读蛋白质组重塑是一种基础适应性反应。
复合体蛋白质和功能性蛋白质通常是共表达的。
研究者使用深度采样策略,将拟南芥组织核心蛋白质组定义为每个组织中约10,000个蛋白质,且在整个植物生命周期中量化近16,000个蛋白质(单细胞的拷贝数)。
整体翻译后修饰蛋白质组学中发现了氨基酸的交换现象,说明了真核生物中可能存在翻译的不忠。
蛋白质丰度的相关性分析揭示了光合作用、种子发育以及衰老和脱落调节中,存在可能组织/年龄特异性转录信号传导模块。
另外,本研究数据表明RD26和其他NAC转录因子具有在种子发育中与耐干旱相关的潜在功能,富含半胱氨酸的受体激酶CRK可以作为衰老中ROS传感器。
核糖体生物发生因子(RBF)复合体的所有组分均以组织、年龄特异的方式共表达,表明拟南芥中这些研究较少的复合体蛋白质组装时功能混杂。
本研究用flg22处理了拟南芥幼苗16小时,分析鉴定了基础免疫反应中蛋白质组结构的特征。
在flg22处理后1、3和16小时,结合平行反应监测(PRM)靶向蛋白质组学,植物激素、氨基酸组分分析和转录本测量获取了一个全面的结果。
研究者发现, JA和JA-Ile水平抑制是通过MYC2(茉莉酸敏感物质1)控制下的IAR3(IAA-ALA抗体3)和JOX2(茉莉酸诱导的加氧酶2)的解共轭和羟基化作用。
这个JA触发的免疫反应调节机制从未被报道,尚未得到充分研究。
本研究生成的数据集广泛覆盖了不同条件中的拟南芥蛋白质组,为相关研究提供了丰富资源。
论文ID原名:Reshaping of the Arabidopsis thaliana proteome landscapeandco-regulation of proteins in development and immunity译名:拟南芥蛋白质组图谱的重塑与蛋白质在发育和免疫中共同调节期刊:Molecular PlantIF:12.084发表时间:2020.09通讯作者:Wolfgang Hoehenwarter通讯作者单位:莱布尼茨植物生物化学研究所实验设计实验结果1. 深度蛋白质组学研究方法因为要对拟南几种组织(组织分别为根,叶,茎生叶,茎,花和角果/种子以及7天至93天衰老的整株植物幼苗)的不同生命阶段进行蛋白质组学测量,及测量用肽flg22处理PTI诱导的蛋白质组。

• 1196 •现物医学 Progress in Modern Biomedicine VoL21 NO.6 MAR2021doi: 10.1324 l/ki.pmb.2021.06.044•专论与综达•呼吸中的疾病诊断标志物及检测技术*罗静怡w孙鹏博u丁艺佩w王阳阳u谢伟东U3A(1清华大学深圳国际研究生院健康科学与技术重点实验室广东深圳518055;2清华大学深圳国际研究生院化学基因组学国家重点实验室广东深圳518055;3清华大学深圳国际研究生院国际开放创新教育中心广东深圳518055)摘要:人类呼出气体中的各种化合物能提供各种疾病和健康状况的重要信息。
近年来,由于红外、电化学、化学发光等新技术的重大突破和质谱仪的使用,使得在极低浓度下精确测量呼出的挥发性有机化合物(V O C s)和气溶胶颗粒成为可能,呼吸检测领域因而取得了重大进展:,呼吸检测因其可以作为一种实时、快速和无创的方法来评估和监测各种疾病与健康状况信息,在科学研究、临床运用中引起了广泛关注。
本综述主要概述呼出气体成分分析方法及在疾病诊断中的研究与应用情况,旨在为将来疾病的实时、快速和无创诊断提供一种新的策略关键词:呼吸气体分析;无创检测技术;疾病诊断中图分类号:Q47;R322.3;R446文献标识码:A文章编号:1673-6273(2021)06-丨196-05Exhaled Breath Markers for Disease Diagnosis and Detection Technologies* LUO Jing-yiu, SUNPeng-bou, DING Yi-peiIJ f WANG Yang-yang1-3, XIE Wei-dong1^(1 Key Lab in Health Science and Technology, Tsinghua Shenzhen Internationa! Graduate School, Tsinghua University, Shenzhen,Guangdong, 518055, China; 2 State Key Laboratory o f C hemical Oncogenomics, Tsinghua Shenzhen International Graduate School, Tsinghua University, Shenzhen, Guangdong, 518055, China: 3 Open FIESTA Center, Tsinghua Shenzhen International Graduate School,Tsinghua University, Shenzhen, Guangdong, 518055, China)ABSTRACT:V a rio u s c o m p o u n d s in h u m an e x h a le d breath can p ro v id e im p ortan t in fo rm a tio n o n v a rio u s d is e a s e s an d h ealth c o n d itio n s. M ajor b reak th rou gh s in n e w t e c h n o lo g ie s su ch a s in frared, e le ctro ch em istry, an d c h e m ilu m in e s c e n c e an d the u se o f m a ss sp ec tro meters h a v e m a d e it p o ss ib le to a c c u r a te ly m ea su re e x h a le d v o la tile o rg a n ic c o m p o u n d s (V O C s) a n d a e r o so l p a rticles at ex tr e m e ly lo w co n cen tra tio n s. S ig n ific a n t p ro g ress h a s b e e n m a d e in th e fie ld o f d etectio n. R esp ir a to ry te stin g h a s attracted w id esp re a d a tten tio n in s c ie n tific resea rch and c lin ic a l a p p lic a tio n s b e c a u se that ca n b e u se d as a r ea l-tim e, fast an d n o n-in v a s iv e m e th o d to ev a lu a te an d m o n ito r info rm a tio n o n v a rio u s d ise a s e s an d h ea lth c o n d itio n s. T h is r e v ie w m a in ly su m m a r iz e s th e a n a ly sis m e th o d s o f e x h a le d breath a n d th eir ap-p lic a tio n in d ise a s e d ia g n o sis, a n d a im s to p ro v id e a n e w stra teg y for the rea l-tim e, rapid an d n o n-in v a s iv e d ia g n o sis o f d is e a s e s in th e future.Key words:E x h a led breath a n a ly sis; N o n-in v a s iv e d e te c tio n te c h n o lo g y; D is e a s e d ia g n o sisChinese Library Classification(CLC):Q47; R322.3; R446 Document code:AArticle ID: 1673-6273(2021 )06-1196-05刖目我们呼出的气体是一个复杂的基质,它由成千上万个的分子构成了一个独特的生物标志,像血液、指纹一样携带着一些与每个人健康有关的信息。

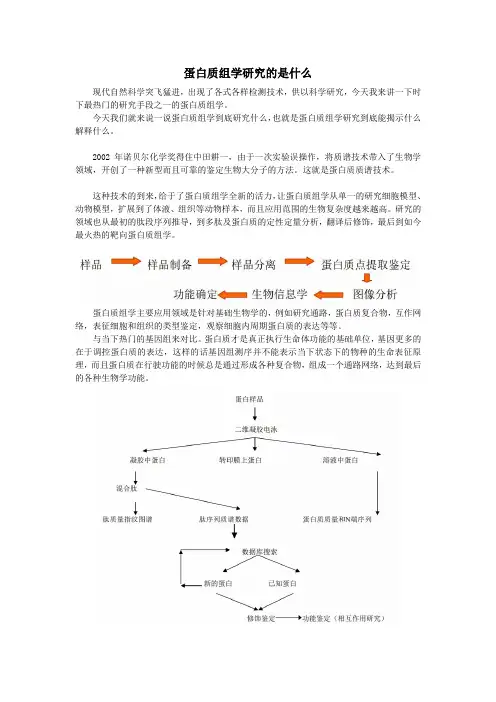
蛋白质组学研究的是什么现代自然科学突飞猛进,出现了各式各样检测技术,供以科学研究,今天我来讲一下时下最热门的研究手段之一的蛋白质组学。
今天我们就来说一说蛋白质组学到底研究什么,也就是蛋白质组学研究到底能揭示什么解释什么。
2002年诺贝尔化学奖得住中田耕一,由于一次实验误操作,将质谱技术带入了生物学领域,开创了一种新型而且可靠的鉴定生物大分子的方法。
这就是蛋白质质谱技术。
这种技术的到来,给于了蛋白质组学全新的活力,让蛋白质组学从单一的研究细胞模型、动物模型,扩展到了体液、组织等动物样本,而且应用范围的生物复杂度越来越高。
研究的领域也从最初的肽段序列推导,到多肽及蛋白质的定性定量分析,翻译后修饰,最后到如今最火热的靶向蛋白质组学。
蛋白质组学主要应用领域是针对基础生物学的,例如研究通路,蛋白质复合物,互作网络,表征细胞和组织的类型鉴定,观察细胞内周期蛋白质的表达等等。
与当下热门的基因组来对比。
蛋白质才是真正执行生命体功能的基础单位,基因更多的在于调控蛋白质的表达,这样的话基因组测序并不能表示当下状态下的物种的生命表征原理,而且蛋白质在行驶功能的时候总是通过形成各种复合物,组成一个通路网络,达到最后的各种生物学功能。
蛋白质组学的研究给我们提供了一个平台,可以从蛋白质系统的层面去考察、思考。
例如:蛋白-蛋白互作,蛋白质的细胞定位,蛋白质的翻译后修饰(磷酸化、甲基化等),以及研究信号通路及代谢通路的调控。
现今最为火热的靶向蛋白质组学更是提供了一个药物研究、医学研究的手段。
以肝毒性为例子,蛋白质组学就能够为药物研发的前期肝毒性评估提供重要的研究手段。
传统蛋白质组学主要是用来鉴定新的未知的生物大分子,但是这种传统的方法在临床药物研究中就出现了重复性低,精准度低的问题。
这时候如果我们在分析之前就明确我们具体要分析的物质是什么的话,这个问题就可以迎刃而解了,这样靶向蛋白质组学就兴起了,我们不用一次性鉴定成千上万个蛋白,只需要从其中挑出几十种甚至十几种研究相关的蛋白质,而且这些蛋白往往浓度较低,用传统的方法十分容易遗漏。

㊀南京农业大学学报㊀2020ꎬ43(4):740-747http://nauxb.njau.edu.cn㊀JournalofNanjingAgriculturalUniversityDOI:10.7685/jnau.201907009收稿日期:2019-07-06基金项目:国家自然科学基金项目(31772648)作者简介:李若楠ꎬ硕士研究生ꎮ∗通信作者:李春梅ꎬ教授ꎬ博士ꎬ主要从事饲料卫生㊁畜禽环境生理与营养调控研究ꎬE ̄mail:chunmeili@njau.edu.cnꎮ李若楠ꎬ康瑞芬ꎬ沈丹ꎬ等.谷氨酰胺对呕吐毒素诱导IPEC ̄J2细胞凋亡和炎症的影响[J].南京农业大学学报ꎬ2020ꎬ43(4):740-747.LIRuonanꎬKANGRuifenꎬSHENDanꎬetal.EffectsofglutamineondeoxynivalenolinducedapoptosisandinflammationofIPEC ̄J2cells[J].JournalofNanjingAgriculturalUniversityꎬ2020ꎬ43(4):740-747.谷氨酰胺对呕吐毒素诱导IPEC ̄J2细胞凋亡和炎症的影响李若楠ꎬ康瑞芬ꎬ沈丹ꎬ戴鹏远ꎬ唐倩ꎬ李春梅∗(南京农业大学动物科技学院ꎬ江苏南京210095)摘要:[目的]本试验以猪空肠上皮细胞(IPEC ̄J2)为模型ꎬ探讨谷氨酰胺(Gln)对呕吐毒素(DON)诱导的IPEC ̄J2细胞凋亡和炎症的影响ꎮ[方法]通过MTT方法测定细胞活力来选择适宜的Gln浓度ꎬ以不添加Gln和DON的细胞为空白对照组ꎬ试验组分别为0.75mmol L-1Gln组ꎬ2.0μg mL-1DON组和2.0μg mL-1DON+0.75mmol L-1Gln组ꎬ处理24h后测定各组细胞凋亡率㊁活性氧自由基(ROS)及细胞凋亡和炎症相关基因和蛋白的表达水平ꎮ[结果]与对照组相比ꎬDON处理24h细胞凋亡比例和ROS含量(ROS荧光密度/细胞总数)显著升高(P<0.01)ꎻ与DON组相比ꎬDON+Gln组显著降低细胞的凋亡比例和ROS含量(P<0.01)ꎮ与对照组相比ꎬDON组上调IPEC ̄J2细胞白细胞介素1β(IL ̄1β)㊁白细胞介素6(IL ̄6)㊁环氧合酶2(COX ̄2)及肿瘤坏死因子α(TNF ̄α)炎症相关基因和Caspase ̄3及Caspase ̄8凋亡相关基因的表达水平ꎻ与DON组相比ꎬDON+Gln组下调IPEC ̄J2细胞IL ̄1β㊁COX ̄2㊁Caspase ̄3㊁Caspase ̄8㊁BAK和Bcl ̄2基因的表达水平ꎮ蛋白免疫荧光结果显示ꎬ与对照组相比ꎬDON组上调IPEC ̄J2细胞Caspase ̄3ꎬ核转录因子κB(NF ̄κB)和磷酸化核转录因子κB(phospho ̄NF ̄κB)蛋白的表达ꎬ而添加Gln后(DON+Gln组)下调了相关蛋白的表达ꎮ[结论]Gln通过清除DON诱导的IPEC ̄J2细胞中过量的ROS及调节炎症和凋亡相关基因及蛋白的表达量来缓解由DON引起的肠道上皮细胞损伤ꎮ关键词:谷氨酰胺ꎻ呕吐毒素ꎻ猪空肠上皮细胞ꎻ炎症ꎻ凋亡中图分类号:S828.9㊀㊀㊀㊀文献标志码:A㊀㊀㊀㊀文章编号:1000-2030(2020)04-0740-08EffectsofglutamineondeoxynivalenolinducedapoptosisandinflammationofIPEC ̄J2cellsLIRuonanꎬKANGRuifenꎬSHENDanꎬDAIPengyuanꎬTANGQianꎬLIChunmei∗(CollegeofAnimalScienceandTechnologyꎬNanjingAgriculturalUniversityꎬNanjing210095ꎬChina)Abstract:[Objectives]Inthisstudyꎬintestinalporcineepithelialcells(IPEC ̄J2cells)wereusedasamodeltoinvestigatetheeffectsofglutamine(Gln)ondeoxynivalenol(DON)inducedapoptosisandinflammationofIPEC ̄J2cells.[Methods]TheappropriateGlnconcentrationwasselectedbyMTTcellviabilityassayꎬandthecellswithoutGlnandDONwereusedasthecontrolgroup.Thetreatedgroupswere0.75mmol L-1Glnꎬ2.0μg mL-1DONand2.0μg mL-1DON+0.75mmol L-1Glnꎬrespectively.After24hꎬtheapoptosisꎬreactiveoxygenspecies(ROS)ꎬsomegenesandproteinsassociatedwithapoptosisandinflammationweremeasured.[Results]ComparedwiththecontrolgroupꎬthepercentageofapoptosisandROScontent(ROSfluorescencedensity/celltotal)significantlyincreasedafterDONtreatmentfor24h(P<0.01).ComparedwiththeDONgroupꎬDON+GlnsignificantlydecreasedthepercentageofapoptosisandROScontent(P<0.01).ComparedwiththecontrolgroupꎬDONup ̄regulatedtheexpressionofgenesinvolvedininflammationandapoptosisꎬsuchasinterleukin ̄1β(IL ̄1β)ꎬinterleukin ̄6(IL ̄6)ꎬcyclooxygenase2(COX ̄2)ꎬtumornecrosisfactoralpha(TNF ̄α)ꎬCaspase ̄3ꎬCaspase ̄8ꎻDON+Glndown ̄regulatedtheexpressionofIL ̄1βꎬCOX ̄2ꎬCaspase ̄3ꎬCaspase ̄8ꎬBAKandBcl ̄2genescomparedwithDONgroup.ProteinimmunofluorescencerevealedthatcomparedwiththecontrolgroupꎬDONup ̄regulatedtheexpressionofCaspase ̄3ꎬnuclearfactorkappaB(NF ̄κB)andphosphorylatednuclearfactorkappaB(phospho ̄NF ̄κB)proteinꎬandGlndown ̄regulatetheexpressionofrelatedproteins.[Conclusions]GlnalleviatedthedamageofIPEC ̄J2cellscausedbyDONbydecreasingROSproductionandregulatingtheexpressionofgenesandproteininvolvedininflammationandapoptosis.Keywords:glutamineꎻdeoxynivalenolꎻintestinalporcineepithelialcell(IPEC ̄J2cell)ꎻinflammationꎻapoptosis147㊀第4期李若楠ꎬ等:谷氨酰胺对呕吐毒素诱导IPEC ̄J2细胞凋亡和炎症的影响㊀㊀呕吐毒素又称脱氧雪腐镰刀菌烯醇(deoxynivalenolꎬDON)ꎬ是由镰刀菌侵染农作物产生的一种B族单端孢霉烯族霉菌毒素ꎬ广泛存在于农作物中ꎬ严重威胁动物和人类的健康[1]ꎮ全球范围内ꎬ呕吐毒素的平均检出率高达59%ꎬ欧洲和亚洲的检出率最高[2]ꎮ龚阿琼等[3]对2017年我国多个地区的饲料原料进行了检测ꎬ经统计分析发现DON在大多数饲料原料中检出率均为100%ꎮDON广泛存在于饲料和人类食品中ꎬ对人和动物的健康造成危害ꎮDON可引起动物呕吐㊁拒食㊁腹泻㊁生长迟缓㊁肠道出血㊁免疫系统紊乱和神经系统疾病[4]ꎮ肠道作为营养物质消化和吸收的主要场所ꎬ亦是抵抗外源毒素和致病性病原微生物入侵的天然屏障[5]ꎮ肠道黏膜上皮是DON主要的靶器官ꎬ经口摄入的DON通过被动扩散进入肠上皮细胞后ꎬ抑制肠上皮细胞的蛋白质合成ꎬ破坏紧密连接蛋白的结构ꎬ损伤肠道屏障的完整性ꎬ增加肠上皮的通透性ꎬ降低营养物质的吸收和转运效率[6]ꎮ另外ꎬ也有研究报道ꎬDON影响细胞增殖ꎬ诱导细胞发生程序性死亡ꎬ改变相关基因的表达[7]ꎮ本课题组前期研究发现ꎬDON可诱发IPEC ̄J2细胞产生过量的ROSꎬ降低细胞的存活率ꎬ诱导细胞凋亡和炎症相关基因及蛋白表达水平发生改变[8]ꎮ谷氨酰胺(glutamineꎬGln)作为一种功能性氨基酸ꎬ可以促进肠道黏膜再生ꎬ增强肠黏膜免疫力ꎬ减少氧化损伤ꎬ刺激肠上皮细胞增殖ꎬ增强肠屏障功能ꎬ维持小肠消化功能和黏膜完整[9]ꎮ在调节基因表达㊁细胞信号㊁抗氧化反应㊁神经传递和免疫等方面发挥着重要作用[10]ꎮ已有研究指出ꎬ仔猪饲料中添加Gln有利于维持仔猪小肠的完整性ꎬ促进小肠黏膜屏障的再生[11]ꎬ但有关Gln是否能缓解毒素造成的肠道损伤尚未见报道ꎮ因此ꎬ本试验以IPEC ̄J2细胞为模型ꎬ探讨Gln对DON诱导的IPEC ̄J2细胞凋亡和炎症反应的影响ꎬ旨在阐明Gln在调节DON诱导的肠上皮细胞凋亡和炎症反应中可能发挥的作用ꎮ1㊀材料与方法1.1㊀试验材料猪空肠上皮细胞(intestinalporcineepithelialcellsꎬIPEC ̄J2)购自北京北纳创联生物有限公司ꎻ呕吐毒素购自美国Sigma公司ꎻ胎牛血清(FBS)㊁抗生素-抗真菌剂㊁DMEM高糖培养基均购自Gibco公司ꎻ荧光定量PCR(qPCR)试剂盒㊁反转录试剂盒购自TaKaRa公司ꎻL-谷氨酰胺(纯度>99.0%)㊁活性氧自由基(ROS)水平检测试剂盒㊁MTT检测试剂盒及AnnexinV ̄FITC细胞凋亡检测试剂盒均购自南京建成生物工程研究所ꎻcleaved ̄Caspase3抗体㊁NF ̄κB(P65)抗体购自美国Abcam公司ꎮ1.2㊀细胞的复苏与培养将冻存在液氮的细胞取出ꎬ于37ħ水浴锅解冻ꎬ转移至15mL离心管ꎬ1000r min-1离心5minꎻ弃上清液后ꎬ加入4mLDMEM高糖培养基(含10%FBS)ꎬ重悬并混匀细胞沉淀ꎬ接种至细胞培养瓶ꎬ放入37ħ㊁含5%CO2的细胞培养箱中培养ꎮ显微镜下观察细胞生长状况ꎬ视实际情况更换细胞培养液ꎬ待细胞融合达到80%时进行传代ꎮ弃上清液ꎬPBS清洗3次后ꎬ加入1mL胰酶并消化1.5minꎬ加入1mL培养基终止消化ꎻ将贴壁的细胞轻轻吹打下来后ꎬ将传代细胞与培养基一起移入新的15mL离心管中ꎬ1000r min-1离心5minꎻ弃上清液后ꎬ加入完全培养基ꎬ重悬混匀后ꎬ接种至细胞培养瓶ꎬ放入细胞培养箱培养ꎬ用于试验处理或继续传代ꎮ1.3㊀细胞活力测定采用MTT方法测定细胞活力ꎮIPEC ̄J2细胞以1ˑ105mL-1细胞浓度接种到96孔板ꎮ当96孔板内的细胞融合度达到80%时ꎬ分别用相同体积的DMEM(CK)㊁Gln(0.25㊁0.75和1.25mmol L-1)㊁DON(2.0μg mL-1)㊁DON+Gln(2.0μg mL-1+0.25mmol L-1㊁2.0μg mL-1+0.75mmol L-1㊁2.0μg mL-1+1.25mmol L-1)处理细胞24hꎬ参照MTT检测试剂盒说明书检测细胞活力ꎮ1.4㊀试验设计与处理以IPEC ̄J2细胞为模型ꎬ试验分为4个组ꎬ分别为细胞对照组(CK)㊁Gln组(0.75mmol L-1Gln)㊁DON组(2.0μg mL-1DON)㊁DON+Gln组(2.0μg mL-1DON+0.75mmol L-1Gln)ꎮDON溶于无水乙醇并配制成1mg mL-1的储存液于-20ħ保存ꎬ用基础培养基稀释至2.0μg mL-1并于4ħ保存ꎮL-谷氨酰胺(纯度>99.0%)用培养基配制成10mmol L-1的储存液于4ħ保存ꎮ将消化后的细胞以均等浓度分别接种到96孔板或6孔板ꎬ当细胞融合度达到80%时ꎬGln㊁DON和DON+Gln组分别用含有DON或(和)Gln的DMEM高糖培养基替代DMEM高糖培养基ꎬ继续培养(4㊁8和24h)进行相关试验ꎮ247南㊀京㊀农㊀业㊀大㊀学㊀学㊀报第43卷1.5㊀细胞ROS水平测定将盖玻片消毒后放入12孔板中ꎬ将100μL细胞悬液按105mL-1的密度接种于12孔板ꎬ4~6h后显微镜下观察细胞是否贴壁ꎬ贴壁后每孔加1mL完全培养基ꎮ细胞融合度达到80%时ꎬ弃掉培养液ꎬ用相同体积的DMEM(CK)㊁Gln(0.75mmol L-1)㊁DON(2.0μg mL-1)㊁DON+Gln(2.0μg mL-1DON+0.75mmol L-1Gln)处理细胞24h后ꎬ参照ROS检测试剂盒说明书检测ROS水平ꎮ操作结束后用激光共聚焦显微镜观察爬片并拍照ꎮ1.6㊀细胞凋亡率测定IPEC ̄J2细胞以2ˑ105mL-1浓度接入6孔板ꎬ当细胞融合度达到80%时ꎬ用不含乙二胺四乙酸(EDTA)的胰酶进行消化ꎬ参照AnnexinV ̄FITC细胞凋亡试剂盒说明书ꎬ用流式细胞仪检测细胞凋亡率ꎮ1.7㊀细胞凋亡和炎症相关基因表达水平的测定细胞以2ˑ105mL-1浓度接入6孔板ꎬ当细胞融合度达到80%时ꎬ提取细胞RNAꎻ测定细胞RNA浓度后ꎬ按照TaKaRa反转录试剂盒说明书对RNA进行反转录ꎬ获得cDNAꎻ测得cDNA浓度后ꎬ将各样品cDNA浓度稀释到100ng μL-1ꎬ-20ħ保存或用于后续试验ꎮ按照TaKaRaSYBR®PremixExTaq试剂盒说明书加入各反应试剂ꎬ在RT ̄qPCR仪上检测细胞凋亡和炎症相关基因的表达ꎮ设定反应程序如下:95ħ30sꎻ95ħ5sꎬ60ħ34sꎬ40个循环ꎻ95ħ15sꎬ60ħ1minꎬ95ħ15sꎮ采用2-ΔΔCT法计算各基因相对表达水平ꎮ表1㊀IPEC ̄J2细胞凋亡和炎症因子相关基因的引物序列Table1㊀Thesequencesofgenesofprimersrelatedapoptosisandinflammatorycytokines基因名称Genename引物序列Sequenceofprimerpairs(5ᶄң3ᶄ)产物长度/bp㊀Productsize㊀㊀β ̄actin㊀㊀CTGGAACGGTGAAGGTGA/TTTGGAAAGGCAGGGACT218㊀㊀IL ̄1β㊀㊀㊀㊀GAGCTGAAGGCTCTCCACCTC/ATCGCTGTCATCTCCTTGCAC223㊀㊀IL ̄6㊀㊀㊀㊀TTCACCTCTCCGGACAAAAC/TCTGCCAGTACCTCCTTGCT122㊀㊀TNF ̄α㊀㊀TTCCAGCTGGCCCCTTGAGC/GAGGGCATTGGCATACCCAC146㊀㊀COX ̄2㊀㊀㊀㊀AGAAGCGAGGACCAGCTTTC/AAAGCGGAGGTGTTCAGGAG215㊀㊀Caspase ̄3㊀㊀㊀㊀GGAATGGCATGTCGATCTGGT/ACTGTCCGTCTCAATCCCAC351㊀㊀Caspase ̄8㊀㊀㊀㊀TCTGCGGACTGGATGTGATT/TCTGAGGTTGCTGGTCACAC165㊀㊀BAK㊀㊀CCCTGTACGTCTACCAGAGG/AACCACAGCCAGAACTAGCA179㊀㊀Bcl ̄2㊀㊀㊀㊀GCTACTTACTGCCAAAGGGA/TTCAGGCGGAGCTGTAAGAG1611.8㊀细胞凋亡和炎症相关蛋白表达的测定利用免疫荧光法测定细胞凋亡和炎症相关蛋白的表达情况ꎮ用相同体积的DMEM(CK)㊁Gln(DMEM+0.75mmol L-1Gln)㊁DON(DMEM+2.0μg mL-1DON)㊁DON+Gln(DMEM+2.0μg mL-1DON+0.75mmol L-1Gln)处理细胞24h后ꎬ将培养基移除ꎬ每孔加入PBS(37ħ预热)1mLꎬ放在摇床上清洗(3次ꎬ每次5min)ꎻ移除PBSꎬ加4%多聚甲醛(1mL)ꎬ室温固定1hꎻ移除多聚甲醛ꎬPBS摇床清洗后ꎬ加入100μLTritonX-100(0.5%)通透10minꎬPBS清洗ꎻ加入1mL1%牛血清白蛋白(BSA)ꎬ室温封闭1hꎻ盖玻片上滴加50μL一抗ꎬ37ħ于湿盒中避光孵育1hꎻPBS清洗后ꎬ滴加50μLAlexaFluor488标记的二抗ꎬ于湿盒中37ħ避光孵育1hꎻ移除二抗ꎬPBS摇床清洗后滴加50μL4ᶄꎬ6-二脒基-2-苯基吲哚(DAPI)ꎬ于湿盒中室温避光孵育10minꎬPBS清洗ꎻ载玻片上滴加荧光抗淬灭剂ꎬ将细胞爬片倒扣在载玻片上封片ꎮ1.9㊀数据统计与分析试验数据用平均数ʃ标准差( xʃSD)表示ꎬ采用SPSS20.0软件进行单因素方差分析(One ̄wayANOVA)ꎬ并用LSD法进行多重比较ꎬ试验数据分析采用GraphPadPrism5软件完成ꎮ2㊀结果与分析2.1㊀Gln对DON诱导IPEC ̄J2细胞活力的影响由图1可知:与对照组相比ꎬ0.75mmol L-1Gln组和1.25mmol L-1Gln组细胞的存活率显著升高(P<0.01)ꎬDON组(2.0μg mL-1)和DON+0.25mmol L-1Gln组细胞的存活率显著降低(P<0.01)ꎬ而DON+0.75mmol L-1Gln组和DON+1.25mmol L-1Gln组细胞的存活率无显著差异ꎮ与DON组相比ꎬDON+0.75mmol L-1Gln和DON+1.25mmol L-1Gln组细胞存活率显著上升(P<0.01)ꎮDON+0.75mmol L-1Gln组㊀第4期李若楠ꎬ等:谷氨酰胺对呕吐毒素诱导IPEC ̄J2细胞凋亡和炎症的影响细胞活力与对照组相近ꎬ因此ꎬ我们选取0.75mmol L-1Gln作为试验剂量ꎮ图1㊀谷氨酰胺(Gln)对呕吐毒素(DONꎬ2.0μg mL-1)诱导IPEC ̄J2细胞活力的影响Fig 1㊀Effectsofglutamine(Gln)onthecellviabilityinIPEC ̄J2inducedbydeoxynivalenol(DON)1)CK:对照组Blankcontrolꎻ0.25㊁0.75㊁1.25Gln表示0.25㊁0.75㊁1.25mmol L-1Glnꎮ0.25ꎬ0.75ꎬ1.25Glnindicate0.25ꎬ0.75ꎬ1.25mmol L-1Gln.2)不同大写字母表示处理间差异显著(P<0.01)ꎮ下同ꎮDifferentcapitallettersabovethebarindicatesignificantdifferenceat0.01level.Thesameasfollows.2.2㊀Gln对DON诱导细胞ROS水平的影响如图2所示:DON(2.0μg mL-1)处理IPEC ̄J2细胞24h后ꎬ与对照组相比ꎬDON组ROS的荧光值显著升高(P<0.01)ꎻ而Gln(0.75mmol L-1)处理IPEC ̄J2细胞24h后ꎬ其ROS的荧光值无显著变化ꎮ与DON组相比ꎬDON+Gln组细胞ROS的荧光值显著降低(P<0.01)ꎮ图2㊀Gln对DON诱导IPEC ̄J2细胞ROS水平的影响(ˑ400)Fig 2㊀EffectsofGlnonROSproductioninducedbyDONinIPEC ̄J2cells㊀㊀CK:对照组BlankcontrolꎻGln:0.75mmol L-1GlnꎻDON:2.0μg mL-1DONꎻDON+Gln:2.0μg mL-1DON+0.75mmol L-1Gln.下同ꎮThesameasfollows.2.3㊀Gln对DON诱导细胞凋亡的影响如图3所示:DON(2.0μg mL-1)处理IPEC ̄J2细胞24h后ꎬ早期凋亡和晚期凋亡细胞的比例显著增加(P<0.01)ꎻ用Gln(0.75mmol L-1)单独处理细胞时ꎬ会显著降低早期凋亡细胞的比例(P<0.01)ꎻDON和Gln同时处理细胞时ꎬ与DON组相比ꎬ早期凋亡和晚期凋亡细胞的比例显著性降低(P<0.01)ꎮ2.4㊀Gln对DON诱导细胞凋亡和细胞炎症相关基因表达的影响2.4.1㊀对细胞凋亡相关基因表达的影响㊀由图4可知:DON(2.0μg mL-1)或Gln(0.75mmol L-1)处理IPEC ̄J2细胞4hꎬ与对照组相比ꎬGln组Caspase ̄3基因表达水平显著升高(P<0.05)ꎬDON+Gln组Caspase ̄3347南㊀京㊀农㊀业㊀大㊀学㊀学㊀报第43卷和Caspase ̄8基因表达水平显著升高(P<0.05)ꎻ与DON组相比ꎬDON+Gln组Caspase ̄3和Caspase ̄8基因的表达水平显著升高(P<0.05)ꎮDON或Gln处理IPEC ̄J2细胞8hꎬ与对照组相比ꎬGln组Bcl ̄2基因的表达水平显著升高(P<0.05)ꎬDON+Gln组Caspase ̄8㊁BAK和Bcl ̄2基因的表达水平显著升高(P<0.05)ꎻ与DON组相比ꎬDON+Gln组Caspase ̄3基因的表达水平显著降低(P<0.05)ꎬBAK㊁Bcl ̄2㊁Caspase ̄8基因的表达水平显著升高(P<0.05)ꎮ24h时ꎬ各基因表达水平在各处理组间无显著性差异ꎮ图3㊀Gln对DON诱导IPEC ̄J2细胞凋亡的影响Fig 3㊀EffectsofGlnonapoptosisinducedbyDONinIPEC ̄J2cells图4㊀Gln对DON诱导IPEC ̄J2细胞凋亡相关基因表达的影响Fig 4㊀EffectsofGlnongenesexpressionsofapoptosisinIPEC ̄J2cellsinducedbyDON㊀㊀相同时间内ꎬ不同小写字母表示处理间差异显著(P<0.05)ꎮAtthesametimeꎬdifferentsmalllettersabovethebarindicatesignif ̄icantdifferenceat0.05level.2.4.2㊀对细胞炎症相关基因mRNA表达的影响㊀由图5可知:DON(2.0μg mL-1)或Gln(0.75mmol L-1)处理IPEC ̄J2细胞4hꎬ与对照组相比ꎬDON组IL ̄6㊁COX ̄2和TNF ̄αmRNA表达水平显著升高(P<0.01)ꎬDON+Gln组COX ̄2㊁IL ̄1β和TNF ̄αmRNA表达水平显著升高(P<0.01)ꎻ与DON组相比ꎬDON+Gln组的IL ̄1β和TNF ̄αmRNA表达水平显著升高(P<0.01)ꎬIL ̄6和COX ̄2mRNA表达水平显著降低(P<0.01)ꎮDON或Gln处理IPEC ̄J2细胞8hꎬ与对照组相比ꎬGln组IL ̄1βmRNA表达水平显著升高(P<0.01)ꎬDON+Gln组IL ̄6mRNA表达水平显著增加(P<0.01)ꎻ与DON组相比ꎬDON+Gln组的IL ̄6mRNA447㊀第4期李若楠ꎬ等:谷氨酰胺对呕吐毒素诱导IPEC ̄J2细胞凋亡和炎症的影响表达水平显著升高(P<0.01)ꎮ24h时ꎬ与对照组相比ꎬGln组IL ̄1βmRNA表达水平显著升高(P<0.01)ꎬDON+Gln组IL ̄6mRNA表达水平显著升高(P<0.01)ꎻ与DON组相比ꎬDON+Gln组IL ̄6mRNA表达水平显著升高(P<0.01)ꎮ图5㊀Gln对DON诱导IPEC ̄J2细胞炎症相关基因表达的影响Fig 5㊀EffectsofGlnongenesexpressionsofinflammationinIPEC ̄J2cellsinducedbyDON㊀㊀相同时间内ꎬ不同大写字母表示处理间差异显著(P<0.01)ꎮAtthesametimeꎬdifferentcapitallettersabovethebarindicatesignificantdifferenceat0.01level.2.5㊀Gln对DON诱导细胞凋亡和细胞炎症相关蛋白表达水平的影响由图6可知:DON(2.0μg mL-1)处理IPEC ̄J2细胞24h后ꎬIPEC ̄J2细胞的cleaved ̄Caspase3蛋白在细胞质内大量表达ꎮ与对照组相比ꎬGln(0.75mmol L-1)组cleaved ̄Caspase3蛋白的荧光强度无显著变化ꎬDON组cleaved ̄Caspase3蛋白的荧光强度极显著升高(P<0.01)ꎻ与DON组比ꎬDON+Gln组细胞的cleaved ̄Caspase3蛋白荧光强度有所降低但无统计学差异ꎻNF ̄κB蛋白在细胞质内被观察到ꎬ且DON处理后其蛋白的荧光强度与对照组相比显著升高ꎮ与对照组相比ꎬGln组NF ̄κB蛋白的荧光强度无显著变化ꎻ与DON组相比ꎬDON+Gln组细胞的NF ̄κB蛋白荧光强度减少但无显著差异ꎻ磷酸化NF ̄κB蛋白由细胞质转移至细胞核而大量表达ꎮ与对照组相比ꎬGln组和DON+Gln组细胞的磷酸化NF ̄κB蛋白荧光强度无显著变化ꎬDON组蛋白的荧光强度显著增加(P<0.05)ꎮ与DON组相比ꎬDON+Gln组细胞的磷酸化NF ̄κB蛋白荧光强度显著降低(P<0.05)ꎮ3㊀讨论氨基酸是生物体内重要的一类物质ꎬ参与机体中蛋白质的合成和各种生理活动ꎮGln作为哺乳动物体内的非必需氨基酸ꎬ可由体内其他氨基酸转化而来ꎬ其代谢后产生的ATP是葡萄糖的3倍ꎬ被认为是小肠的主要供能物质[12]ꎮGln具有抗氧化功能ꎬ可以促进谷胱甘肽过氧化物酶基因的表达ꎬ还可通过氧化供能调节信号传递过程中细胞凋亡酶的活性ꎬ抑制一氧化氮的合成速率ꎬ参与由TNF ̄α诱导的细胞毒性作用和抗氧化物质谷胱甘肽的合成[13]ꎮROS检测结果表明ꎬ添加Gln可以降低由DON引起的过量ROSꎬ这可能是由于谷胱甘肽促进谷胱甘肽过氧化物酶的合成ꎬ从而提高细胞抗氧化能力ꎬ清除部分氧自由基ꎬ从而降低ROS水平ꎮEvans等[14]研究发现ꎬ在HT ̄29结肠癌细胞中由TNF ̄α相关凋亡诱导配体(TRAIL)引起的细胞凋亡会提高细胞核的聚缩ꎬ激活Caspase ̄8和Caspase ̄3的表达ꎻ添加Gln可抑制由TRAIL引起的细胞凋亡ꎬ但其缓解由细胞因子升高引起的细胞凋亡机制与细胞GSH介导的抗氧化应激机制并不相同ꎮ本试验结果发现ꎬ添加Gln可降低由DON诱导的IPEC ̄J2细胞凋亡相关蛋白cleaved ̄Caspase3的表达ꎮ因此ꎬ我们猜想Gln对肠上皮细胞的保护作用可能与凋亡相关蛋白表达的改变相关ꎮGln作为供能物质ꎬ当机体经口或非经口途径摄入Gln后肠道的细胞因子水平会有所降低ꎮ在对人547647卷南㊀京㊀农㊀业㊀大㊀学㊀学㊀报第43图6㊀Gln对DON诱导IPEC ̄J2细胞cleaved ̄Caspase3㊁NF ̄κB和磷酸化NF ̄κB蛋白表达的影响Fig 6㊀EffectsofGlnontheproteinexpressionofcleaved ̄Caspase3ꎬNF ̄κBandphospho ̄NF ̄κBinducedbyDONinIPEC ̄J2cells㊀㊀上:染色图ꎬˑ400ꎻ下:统计图ꎮTop:Coloringchartꎬˑ400ꎻBottom:Statisticalchart.不同大小写字母分别表示在0.01和0.05水平上差异显著ꎮDifferentcapitalandsmalllettersabovethebarindicatesignificantdifferenceat0.01and0.05levels.体进行肠道活检时发现ꎬ采用Gln治疗的病人肠道促炎细胞因子IL ̄6和IL ̄8基因表达量显著降低ꎬ这主要是因为Gln抑制了NF ̄κB的激活[15]ꎮ脑部创伤大鼠模型注射Gln后发现ꎬGln会缓解大鼠肠道组织形态损伤并降低肠道内细胞因子的表达量ꎬ其缓解作用可能是通过降低肠道内NF ̄κB蛋白的表达量来实现的[16]ꎮ谷氨酰胺对急性肺损伤小鼠具有保护作用[17]ꎮ本试验结果表明ꎬGln处理24h可以降低IPEC ̄J2细胞促炎细胞因子IL ̄6和COX ̄2基因表达量ꎬ降低NF ̄κB和磷酸化NF ̄κB蛋白的表达量ꎬ提高细胞的抗炎能力ꎬ但在处理4h和8h时ꎬGln促进炎症因子IL ̄6和IL ̄1β基因的表达ꎮ因此ꎬ我们推测Gln缓解细胞炎症反应与处理时间有关ꎬ短时间内炎症因子的释放可能是细胞受到外界刺激启动防御机制的一种表现ꎬ随处理时间的延长才显现出Gln的缓解作用ꎬ炎症相关因子的表达量开始呈降低趋势ꎮTNF ̄α是巨噬细胞被激活后产生的细胞因子ꎬ具有细胞毒性ꎬ其毒性作用会抑制Gln的蛋白质合成功能[18]ꎮ研究发现ROS通过诱导细胞TNF ̄α的增加ꎬ促进肾小球上皮细胞凋亡ꎬ而加入过氧化物清除剂后ꎬ凋亡比例降低[19]ꎮ本试验结果显示ꎬ虽然Gln可以清除ROSꎬ但DON+Gln组TNF ̄α基因的表达量与DON组相比显著升高ꎬ这可能与Gln的处理时间㊁靶向性㊁特异性和不同抗氧化剂的过度补偿作用有关ꎮ在基因水平上ꎬGln促进IPEC ̄J2细胞内TNF ̄α基因的表达ꎬ进而促进Caspase ̄3及Caspase ̄8基因的表达ꎮ但蛋白表达结果表明Gln降低细胞凋亡和炎症相关蛋白的表达ꎬ因此ꎬ试验结果最终表现为细胞凋亡相关蛋白的表达量降低ꎬ细胞凋亡比例降低ꎮ本研究结果表明ꎬGln通过清除IPEC ̄J2细胞中过量的ROS㊁调节凋亡和炎症相关基因及蛋白的表达水平来缓解由DON引起的肠道上皮细胞损伤ꎬ该结果为Gln在动物生产中的应用提供了基础理论依据ꎮ参考文献References:[1]㊀LiRꎬLiYꎬSuYꎬetal.Short ̄termingestionofdeoxynivalenolinnaturallycontaminatedfeedalterspigletperformanceandguthormonesecretion[J].AnimalScienceJournalꎬ2018ꎬ89(8):1134-1143.[2]JiFꎬXuJꎬLiuXꎬetal.NaturaloccurrenceofdeoxynivalenolandzearalenoneinwheatfromJiangsuProvinceꎬChina[J].FoodChemistryꎬ2014ꎬ747㊀第4期李若楠ꎬ等:谷氨酰胺对呕吐毒素诱导IPEC ̄J2细胞凋亡和炎症的影响157:393-397.[3]龚阿琼ꎬ全明旭ꎬ周锋ꎬ等.2017年上半年我国饲料原料及配合饲料霉菌毒素检测分析[J].中国饲料ꎬ2017(20):41-43.GongAQꎬQuanMXꎬZhouFꎬetal.DetectionandanalysisofmycotoxininfeedingredientsandcompoundfeedinChinainthefirsthalfof2017[J].ChineseFeedꎬ2017(20):41-43(inChinesewithEnglishabstract).[4]WuQꎬKu㊅caKꎬHumpfHUꎬetal.Fateofdeoxynivalenolanddeoxynivalenol ̄3 ̄glucosideduringcereal ̄basedthermalfoodprocessing:areviewstudy[J].MycotoxinResearchꎬ2017ꎬ33(1):79-91.[5]Gonz lez ̄CastroAMꎬMartínezCꎬSalvo ̄RomeroEꎬetal.Mucosalpathobiologyandmolecularsignatureofepithelialbarrierdysfunctioninthesmallintestineinirritablebowelsyndrome[J].JournalofGastroenterologyandHepatologyꎬ2017ꎬ32(1):53-63.[6]AwadWAꎬZentekJ.Thefeedcontaminantdeoxynivalenolaffectstheintestinalbarrierpermeabilitythroughinhibitionofproteinsynthesis[J].ArchivesofToxicologyꎬ2015ꎬ89(6):961-965.[7]BracarenseAPꎬLucioliJꎬGrenierBꎬetal.Chronicingestionofdeoxynivalenolandfumonisinꎬaloneorininteractionꎬinducesmorphologicalandimmunologicalchangesintheintestineofpiglets[J].BritishJournalofNutritionꎬ2012ꎬ107(12):1776-1786.[8]KangRꎬLiRꎬDaiPꎬetal.DeoxynivalenolinducedapoptosisandinflammationofIPEC ̄J2cellsbypromotingROSproduction[J].EnvironmentalPollutionꎬ2019ꎬ251:689-698.[9]LiuYꎬXiongXꎬTanBEꎬetal.Nutritionalinterventionfortheintestinaldevelopmentandhealthofweanedpigs[J].FrontiersinVeterinaryScienceꎬ2019ꎬ6:46-59.[10]WangHꎬZhangCꎬWuGYꎬetal.Glutamineenhancestightjunctionproteinexpressionandmodulatescorticotropin ̄releasingfactorsignalinginthejejunumofweanlingpiglets[J].JournalofNutritionꎬ2015ꎬ145(1):25-31.[11]XingSꎬZhangBLꎬLinMꎬetal.Effectsofalanyl ̄glutaminesupplementationonthesmallintestinalmucosabarrierinweanedpigletschallengedwithlipopolysaccharide[J].CanadianJournalofAnimalScienceꎬ2017ꎬ98(1):144-155.[12]WangBꎬWuGꎬZhouZꎬetal.Glutamineandintestinalbarrierfunction[J].AminoAcidsꎬ2015ꎬ47(10):2143-2154.[13]DingZꎬLiWꎬHuangJꎬetal.Dietaryalanyl ̄glutamineandvitaminEsupplementscouldconsiderablypromotetheexpressionofGPxandPPARαgenesꎬantioxidationꎬfeedutilizationꎬgrowthꎬandimprovecompositionofjuvenilecobia[J].Aquacultureꎬ2017ꎬ470:95-102.[14]EvansMEꎬJonesDPꎬZieglerTR.Glutaminepreventscytokine ̄inducedapoptosisinhumancolonicepithelialcells[J].JournalofNutritionꎬ2003ꎬ133(10):3065-3071.[15]KimMHꎬKimH.Therolesofglutamineintheintestineanditsimplicationinintestinaldiseases[J].InternationalJournalofMolecularSciencesꎬ2017ꎬ18(5):1051-1066.[16]ChenGꎬShiJꎬQiMꎬetal.GlutaminedecreasesintestinalnuclearfactorkappaBactivityandpro ̄inflammatorycytokineexpressionaftertraumaticbraininjuryinrats[J].InflammationResearchꎬ2008ꎬ57(2):57-64.[17]黄杰ꎬ刘京ꎬ马娜娜ꎬ等.谷氨酰胺对急性肺损伤小鼠MUC5AC的调节及其对肺脏的保护作用[J].畜牧与兽医ꎬ2019ꎬ51(4):97-103.HuangJꎬLiuJꎬMaNNꎬetal.RegulativeandprotectiveeffectsofglutamineontheexpressionofMUC5AC[J].AnimalHusbandry&VeterinaryMedicineꎬ2019ꎬ51(4):97-103(inChinesewithEnglishabstract).[18]ZhouJꎬFanSꎬCaoYꎬetal.Tumornecrosisfactor ̄αsuppressestheproteinfractionalsynthesisrateofthesmallintestinestimulatedbyglutamineinrats[J].ExperimentalandTherapeuticMedicineꎬ2015ꎬ9(2):547-552.[19]SverrissonKꎬAxelssonJꎬRippeAꎬetal.Acutereactiveoxygenspecies(ROS) ̄dependenteffectsofIL ̄1βꎬTNF ̄αꎬandIL ̄6ontheglomerularfiltrationbarrier(GFB)invivo[J].AmericanJournalofPhysiologyRenalPhysiologyꎬ2015ꎬ309(9):F800-F806.责任编辑:周广礼。

细胞表面的聪明受体作者:王曼徐华强来源:科学杂志()录入:Admin 字体:构成生物体数以亿计的细胞并非孤立地存在,而是一直处于互相联系中,感知周围环境的变化,作出相应反应,因为它们表面有“聪明”的受体。
强光使人闭眼、花香使人愉悦、黑暗使人恐惧……,你有没有想过,人的大脑是如何感知外部环境变化并作出反应的?人的身体由数万亿个细胞组成并精密协调地工作,完成各种生理功能,这其中又是什么充当传感器来传递信息,使细胞感知周围环境。
这些问题一直是个谜,在20世纪的大部分时间里,人们不清楚充当传感器的这些物质是由什么组成的,以及它们如何工作。
两位医生出身的科学家经过不懈的努力,揭示了其中的奥秘:细胞表面存在着一类称为G 蛋白偶联受体(G-protein-coupled receptor,简称GPCR)的蛋白质,它将从外界接受的不同信号分子,通过自身构象的变化激活细胞异源三聚体的鸟苷酸结合蛋白(即G蛋白),后者将信号传至胞的效应分子引起反应。
这两位科学家是美国霍华德·休斯医学研究所和美国杜克大学医学中心医学教授莱夫科维茨(Robert Joseph Lefkowitz),斯坦福大学医学院分子与细胞生理学教授科比尔卡(Brian Kent Kobilka)。
因为他们突破性地揭示了G蛋白偶联受体的在工作机制,共同获得了2012年度诺贝尔化学奖。
莱夫科维茨1968年利用放射性追踪细胞受体,将碘同位素标记糖皮质激素和肾上腺素,证明了受体独立于腺苷酸环化酶(AC)的存在,随后分离纯化了β2肾上腺素能受体(β2-adrenergic receptor),证实了细胞表面受体的存在。
后加入的科比尔卡则将编码β-肾上腺素能受体的基因从人类基因组中分离出来,并创造性地获得了β-肾上腺素能受体被激素激活并向细胞发送信号的精确图像,弄清了它的三维结构及其信号转导机制。
什么是G 蛋白偶联受体生物体的基本单位是细胞,细胞由细胞膜及其的细胞质和细胞核组成,外界信号进入细胞首先要通过细胞膜,水和氧气等小分子物质能自由通过,而某些离子和大分子物质则不能自由通过。

基于蛋白质组学的植物耐盐机制和分子标记研究植物是我们生活中不可或缺的一部分。
而随着人类活动的不断增加,环境的污染和全球气候变化问题日益严峻,这也在很大程度上影响了植物的生长和发展。
其中,盐渍化是一个普遍存在的现象,特别是在世界上许多干旱的地区。
为了使植物适应这种环境,科学家们从不同的角度去研究植物的适应性。
其中,基于蛋白质组学的研究是当前研究植物耐盐机制和分子标记的重要手段之一。
一、植物耐盐机制研究植物的耐盐性主要是通过减轻和适应两种方式来完成的。
减轻盐胁迫是指通过减少盐分进入植物体内以及提高盐分排出量的策略。
适应盐胁迫则是指增强植物对盐分的耐受性,使植物在含盐环境中仍能正常生长发育。
具体来说,植物的耐盐性主要表现在以下几个方面:(1)盐胁迫信号传导(Na+、K+/Na+交换、Ca2+等):盐胁迫会影响植物细胞膜的离子通道活性,导致细胞膜的通透性发生变化。
其中,Na+、K+/Na+交换和Ca2+等重要离子在植物的耐盐过程中起到了重要的作用。
因此,它们的信号传导机制以及相关的激素调控机制在植物耐盐性的研究中是非常重要的。
(2)离子平衡:由于盐分会影响植物体内K+、Na+、Cl-等离子的浓度平衡,而这种失衡会影响植物的生长和发育。
因此,在植物耐盐性的研究中,研究植物细胞膜上的离子通道的调控机制和离子平衡机制非常重要。
(3)保护酶和蛋白质的积累:受盐分胁迫影响,植物体内会增加一些保护酶(如超氧化物歧化酶和过氧化物酶)和特定的蛋白质(如HSPs)的积累,防止盐分对植物生长和发育的影响。
(4)激素参与:除细胞内离子之外,激素在植物较强的耐盐机制中也发挥了重要作用。
其中,ABA作为最重要的激素之一,在植物耐盐性研究中扮演着重要的角色。
二、基于蛋白质组学的研究随着蛋白质组学技术的快速发展,科学家们利用蛋白质组学技术研究植物对盐胁迫的适应性和耐受性机制,已经成为当前植物生物技术和分子生物学研究的热点之一。
(1)蛋白质组成分析:通过蛋白质组成分析可以了解植物胁迫后产生的蛋白质变化。

前沿空间蛋⽩质组学:⼀种强⼤的细胞⽣物学发现⼯具真核细胞⾼度区室化,⽣物过程被分隔在不同的区室进⾏。
蛋⽩质功能与亚细胞定位密切相关,不同的区室提供不同的化学环境(例如pH和氧化还原条件)、不同的潜在作⽤配体或底物。
因此,对蛋⽩质亚细胞定位的严格控制是细胞⽣理学的重要调控内容。
⼤多数细胞⽣物学过程涉及蛋⽩质亚细胞定位的变化,例如转录因⼦在细胞核-胞浆的穿梭、细胞凋亡过程中线粒体蛋⽩的重新定位,以及细胞表⾯信号传导受体的内吞等。
相反,蛋⽩质的错误定位通常与细胞功能障碍和疾病相关,包括神经变性、癌症和代谢紊乱等。
以蛋⽩质空间定位为研究⽅向的空间蛋⽩质组现在已经⽤于揭⽰⼈类蛋⽩质组的复杂结构,如单细胞变异、动态蛋⽩质易位,相互作⽤⽹络改变,以及蛋⽩定位改变等。
⼀些研究者也已成功运⽤空间蛋⽩质组学来研究疾病,包括急性病毒感染、肝病等。
2019年5⽉,KTH 瑞典皇家理⼯学院的 Emma Lundberg 教授和德国马克斯-普朗克研究所的Georg H. H. Borner 教授,在国际著名期刊 Nature Reviews | Molecular CellBiology(IF=35.612)发表了题为《Spatial proteomics: a powerful discovery tool for cellbiology》的综述性⽂章,系统介绍了空间蛋⽩质组学的技术、未来发展的机遇与挑战。
下⾯⼩编为⼤家解读⼀下这篇综述。
01空间蛋⽩质组学研究⽅法⽬前三种互补的⽅法可⽤于空间蛋⽩质组学研究:细胞器分级的质谱分析(图1)、蛋⽩质与蛋⽩质互作⽹络分析(图2),以及基于蛋⽩质定位的蛋⽩质成像(图3)。
1)基于质谱的细胞器分级⽅法质谱可⽤于复杂混合物中蛋⽩质的定性和定量研究。
空间蛋⽩质组学可借助传统⽣物化学分析⽅法,如下图1a 所⽰,通过定制的亚细胞分级分离(如,梯度离⼼或差速离⼼)来富集⽬标细胞器(绿⾊)。
然后利⽤质谱⽅法只分析富集到的组分。

基于遗传定量分析的功能基因组学研究随着人类基因组计划的完成及高通量测序技术的发展,生物学科学家们开始借助大规模基因数据对整个物种的生命现象进行全方位的分析。
该过程中涌现出了很多基因组学或生物信息学相关研究领域,如蛋白组学、转录组学、代谢组学、表观遗传学、系统生物学等。
其中,功能基因组学作为一门结合基因组科学和生物学的交叉学科,旨在揭示基因组中的大量序列信息的为生物系统的生物学功能所贡献的意义,成为现代生物学研究中一颗强有力的研究利器。
功能基因组学是基于遗传定量分析的一项研究领域。
它可以为我们提供一种有效的方式,来对基因组中大量生物学信息的序列中的关键信息进行识别和解释。
其核心思想是通过在分子和细胞水平研究基因和其功能时,遗传定量分析能够给出关于基因的生物学功能的定量描述,甚至还能对复杂生物系统中的基因和调节网络进行定量建模和预测。
这些方法现在已经成为广大生物学家研究基因组中的序列信息和生物学功能的非常重要的工具之一。
功能基因组学的核心技术有两个: 定量遗传解析(Quantitative genetic analysis)和系统生物学(Systems biology)。
定量遗传解析是功能基因组学中最为重要和最基础的工具。
基于遗传学研究的定量遗传解析,在揭示基因功能,从而推断复杂生物学行为的分子机制方面有特殊的优势。
它允许我们对多基因性状(如复杂病和中等大小)进行研究,并探索遗传和环境因素之间的相互作用。
借助定量遗传解析,我们可以控制一些变量,通过它们之间的相互作用来解释个体群体表型的差异。
这些研究中,研究人员可以对大量植物或动物进行基因组分析,并用复杂分析方法解析群体遗传学数据,解决许多生物学上的问题。
系统生物学是功能基因组学最为新兴的研究领域之一。
随着现代生物学研究的深入和科技的发展,生物学家们开始用定量系统生物学方法来研究整体生物系统,从基因和蛋白质相互作用到整体生物系统的稳态行为。
系统生物学将已知的基因,代谢反应和蛋白相互作用等生物学系统看做是一个整体,从而研究它们之间的相互作用和动力学的机制,以此增进我们对生物系统的理解和控制。
MS(mass spectrum)质谱法:一种测量离子质荷比的分析方法tamdam-MS 串联质谱:将两个或更多质谱连接在一起。
MALDI(Matrix-Assisted laser Desrption Ionization)基质辅助激光解吸电离离子源:用激光照射样品与基质形成的光结晶薄膜、基质从激光中吸收能量传递到生物分子,而电解过程中将质子转移到生物分子或生物分子得到质子而使生物分子电离的过程ESI(Electron spray Ionization)点喷雾离子源:属于一种软电离源,能使大质量的有机分子生成多电荷的离子。
2D-DIGE(two dimension difference gel electrophoresis)一种新的荧光标记的定量蛋白质组学技术。
利用电荷和分子量的差异分离蛋白质混合物,并通过多通道激光扫描分析不同蛋白质的第二向SDS-PAGE凝胶图像。
zoom gel 变焦镜头凝胶成像:利用百变焦镜头的设备对凝胶进行拍摄成像。
two-in-one-gel 二合一凝胶:将相同PH范围胶条一起进行SDS-PAGE的技术SILAC(stable isotope labeling with amino acids in cell culture)细胞培养下稳定同位素标记技术):是在细胞培养过程中,利用稳定同位素标记的氨基酸结合质谱技术对蛋白表达进行定量分析的技术。
CDIT(charge detection ion trap)电荷检测离子阱:一种将离子通过电磁场限定在有限空间内的设备,用于多级质谱分析。
ICAT(Isotope Coded Affinity Taq)同位素亲和标签技术:用具有不同质量同位素亲和标签标记处于不同状态下的细胞中的半胱氨酸,利用串联质谱技术对混合样品进行质谱分析。
iTRAQ(isobaric taqs fur relative and absolute guentitation)同位素标记相对和绝对定量技术:该技术利用多种同位素试剂标记蛋白多肽N末端或赖氨酸侧链基团,经高精度质谱仪分析,可同时比较8种样品间蛋白表达量。
《基因组学与蛋白质组学》课程教学大纲学时:40学分:2.5理论学时:40实验学时:0面向专业:生物科学、生物技术课程代码:B7700005先开课程:生物化学、分子生物学课程性质:必修/选修执笔人:朱新产审定人:第一部分:理论教学部分一、课程的性质、目的和任务《基因组学与蛋白质组学》是随着生物化学、分子生物学、结构生物学、晶体学和计算机技术等的迅猛发展而诞生的,是融合了生物信息学、计算机辅助设计等多学科而发展起来的新兴研究领域。
是当今生命科学研究的热点与前沿领域。
由于基因组学与蛋白质组学学科的边缘性,所以本课程在介绍基因组学与蛋白质组学基本基本技术和原理的同时,兼顾学科发展动向,讲授基因组与蛋白组学中的热点和最新进展,旨在使学生了解现代基因组学与蛋白质组学理论的新进展并为相关学科提供知识和技术。
二、课程的目的与教学要求通过本课程的学习,使学生掌握基因组学与蛋白质组学的基本理论、基础知识、主要研究方法和技术以及生物信息学和现代生物技术在基因组学与蛋白质组学上的应用及典型研究实例,熟悉从事基因组学与蛋白质组学的重要方法和途径。
努力培养学生具有科学思维方式、启发学生科学思维能力和勇于探索,善于思考、分析问题的能力,激发学生的学习热情,并通过学习提高自学能力、独立思考能力以及科研实践能力,为将来从事蛋白质的研究奠定坚实的理论和实践基础。
三、教学内容与课时分配第一篇基因组学第一章绪论(1学时)第一节基因组学的研究对象与任务;第二节基因组学发展的历程;第三节基因组学的分子基础;第四节基因组学的应用前景。
本章重点:1. 基因组学的概念及主要任务;2. 基因组学的研究对象。
本章难点:1.基因组学的应用及发展趋势;2.基因组学与生物的遗传改良、人类健康及生物进化。
建议教学方法:课堂讲授和讨论思考题:查阅有关资料,了解基因组学的应用发展。
第二章人类基因组计划(1学时)第一节人类基因组计划的诞生;第二节人类基因组研究的竞赛;第三节人类基因组测序存在的缺口;第四节人类基因组中的非编码成分;第五节人类基因组的概观;第六节人类基因组多样性计划。
糖尿病肾病小鼠中前纤维蛋白1的表达和免疫细胞浸润分析*麦丽萍, 黄桂萍, 邓春玉, 郑丹琳, 李晓红, 何国东△[南方医科大学附属广东省人民医院(广东省医学科学院)医学研究部,广东 广州 510080][摘要] 目的:探讨前纤维蛋白1(profilin 1, PFN1)在糖尿病肾病小鼠中的表达及其与免疫细胞浸润的相关性。
方法:利用糖尿病肾病患者肾组织芯片转录组表达数据分析PFN1的表达水平和免疫细胞浸润情况,并通过小鼠动物实验进行验证。
将16只C57BL/6小鼠随机分为正常组和模型组,每组8只。
模型组采用腹腔注射链脲佐菌素法建立糖尿病肾病小鼠模型,造模成功后检测小鼠肾组织中CD11b 、F4/80、CC 趋化因子受体4(CC chemo‐kine receptor 4, CCR4)、白细胞介素1受体I 型(interleukin -1 receptor type I , IL -1R1)、B 细胞淋巴瘤2(B -cell lympho‐ma -2, Bcl -2)、Bcl -2相关X 蛋白(Bcl -2-associated X protein , Bax )和胱天蛋白酶3(caspase -3)蛋白的表达水平。
另外,在糖尿病肾病细胞模型中过表达PFN1,检测单核细胞趋化蛋白1(monocyte chemotactic protein -1, MCP -1)和cas‐pase -3蛋白的表达情况。
结果:糖尿病肾病患者肾组织芯片转录组表达数据集GSE30122中,PFN1基因表达量显著升高(P <0.01),且与巨噬细胞和T 细胞有显著相关性。
糖尿病肾病小鼠模型组肾脏组织可见明显病理改变,肾脏组织中PFN1表达量显著升高(P <0.01),免疫指标CD11b 、F4/80、CCR4、IL -1R1和凋亡相关蛋白Bax 、Bcl -2、cas‐pase -3表达升高(P <0.01)。
植物激酶底物的定量和定位研究引言:植物激酶底物的定量和定位研究是植物生物学的一个重要研究方向。
植物激酶是一类能够催化底物磷酸化的酶,对植物生长发育、逆境响应及信号传导起到重要作用。
植物激酶底物的定量和定位研究可以揭示植物激酶的底物特异性和调控机制,为进一步解析信号传递网络提供重要依据。
一、植物激酶底物定量的方法1.蛋白质质谱法蛋白质质谱法是目前最常用的植物激酶底物定量方法之一、该方法通过将激酶底物和激酶一起孵育,并利用质谱技术定量激酶底物的磷酸化水平。
这个方法的优势是可以同时定量多个激酶底物,且可以在不同激酶底物之间进行比较,并进一步揭示激酶抉择性的机制。
2.免疫印迹法免疫印迹法也是一种常用的植物激酶底物定量方法。
该方法通过利用特异性的抗体识别磷酸化的底物蛋白质,并利用相关的信号转导技术进行定量分析。
这个方法的优势是准确、灵敏,能够实时监测激酶底物的磷酸化水平,并定量比较不同样品的差异。
二、植物激酶底物定位的方法1.免疫染色法免疫染色法是一种常用的植物激酶底物定位方法。
该方法通过利用特异性的抗体结合磷酸化底物蛋白质,并利用荧光染料或酶标记技术进行标记,从而实现对激酶底物的定位。
这个方法的优势是可以在细胞和组织水平上直接观察激酶底物的位置和表达水平,进一步揭示激酶底物的细胞定位和分布情况。
2.荧光标记法荧光标记法是一种新兴的植物激酶底物定位方法。
该方法通过将荧光蛋白标记在激酶底物上,利用荧光显微镜技术实时观察激酶底物在细胞内的分布和迁移情况。
这个方法的优势是高分辨率、快速,能够实时监测激酶底物的动态变化,揭示激酶底物与信号通路的关系。
结论:植物激酶底物的定量和定位研究为解析激酶底物网络的调控机制和信号传递网络提供了重要手段。
目前,蛋白质质谱法和免疫印迹法是主要的定量方法,免疫染色法和荧光标记法是主要的定位方法。
这些方法的应用可以揭示激酶底物的动态调控和信号通路的空间分布,从而为进一步理解植物生长发育和逆境响应的分子机制提供重要线索和依据。