生化实验01-蛋白质含量的测定
- 格式:ppt
- 大小:640.50 KB
- 文档页数:14

蛋白质含量的测定:1. 样品与浓硫酸和催化剂一同加热消化,使蛋白质分解,其中碳和氢被氧化为二氧化碳和水逸出,而样品中的有机氮转化为氨与硫酸结合成硫酸铵。
然后加碱蒸馏,使氨蒸出,用硼酸吸收后再以标准盐酸或硫酸溶液滴定。
根据标准酸消耗量可计算出蛋白质的含量。
2NH3 + 4H3BO3=(NH4)2B4O7+5H2O(NH4)2B4O7+2HCl+5H2O=2NH4Cl+4H3BO3 F=5.71可知,蛋白质与氮的比例是5.71,每一摩尔氮对应一摩尔盐酸①凯氏定氮法原理:蛋白质平均含氮量为16%。
当样品与浓硫酸共热,蛋白氮转化为铵盐,在强碱性条件下将氨蒸出,用加有指示剂的硼酸吸收,最后用标准酸滴定硼酸,通过标准酸的用量即可求出蛋白质中的含氮量和蛋白质含量。
②双缩脲法原理:尿素在180℃下脱氨生成双缩脲,在碱性溶液中双缩脲可与Cu2+形成稳定的紫红色络合物。
蛋白质中的肽键实际上就是酰胺键,故多肽、蛋白质等都有双缩脲(biuret)反应,产生蓝色或紫色复合物。
比色定蛋白质含量。
缺点:灵敏度低,样品必须可溶,在大量糖类共存和含有脯氨酸的肽中显色不好。
其精确度较差(数mg),且会受样品中硫酸铵及Tris 的干扰,但准确度较高,不受蛋白质的种类影响。
③Folin酚法(Lowry)Folin酚法是biuret 法的延伸,所用试剂由试剂甲和乙两部分组成。
试剂甲相当于双缩脲试剂(碱性铜试剂),试剂乙中含有磷钼酸和磷钨酸。
在碱性条件下,蛋白质中的巯基和酚基等可将Cu2+还原成Cu+,Cu+能定量地与Folin-酚试剂反应生成蓝色物质,600nm比色测定蛋白质含量。
灵敏度较高(约0.1 mg),但较麻烦,也会受硫酸铵及硫醇化合物的干扰。
步骤中各项试剂的混合,要特别注意均匀澈底,否则会有大误差。
④紫外法280nm光吸收法:利用Tyr在280nm在吸收进行测定。
280nm-260nm的吸收差法:若样品液中有少量核酸共存按下式计算:蛋白质浓度(mg/ml)=1.24E280-0.74E260 (280 260为角标)⑤色素结合法(Bradford 法)直接测定法:利用蛋白质与色素分子(Coomassie Brilliant Blue G-250)结合物的光吸收用分光光度法进行测定。

蛋白质生化实验报告生殖免疫研究所薛樱子学号:1133111003实验一溶液中蛋白质浓度的测定一光吸收法(测量范围:0.1—2mg)1实验原理:由于蛋白质中存在着含有共轭双键的酪氨酸和色氨酸,它们具有吸收紫外光的性质,其吸收高峰在280nm波长处,且在此波长内吸收峰的光密度值OD280nm与其浓度成正比关系,故可作为蛋白质定量测定的依据。
纯蛋白的A280/A260为 1.8,纯核酸的A280/A260为2步骤:2.1)打开仪器的电源开关(接220V交流电),打开比色槽暗箱盖,选择光源,选档,选波长,用调零旋钮调暗电流至0 。
2.2)将空白对照样品和待测溶液装入石英比色杯2.3)将仪器的比色槽暗箱合上,比色槽处于蒸馏水(或校正缓冲液)校正位置,旋转光量调节器使电表指针正确处于0 。
2.4)拉出比色槽手柄拉杆使比色槽处于样品位置读数。
2. 5)在260nm和280nm分别读数(分别用缓冲液调0),根据上表查处相应蛋白浓度,根据喜事倍数计算原溶液蛋白浓度,根据体积计算总蛋白量。
3结果与分析:A280=0.571 A260=0.340根据公式:蛋白浓度=1.5 ×A280 —0.75 ×A260=1.5×0.571—0.75×0.340=0.6015也可以根据蛋白质和核算含量折算图表画出一条直线估算蛋白质的浓度。
结果说明我的蛋白样品浓度是0.6015mg/ml。
二Folin—酚法(测量范围:10-300ug/ml)1实验原理:在碱性条件下,蛋白质中的肽键与铜结合生成复合物。
Folin—酚试剂中的磷钼酸盐—磷钨酸盐被蛋白质中的酪氨酸和苯丙氨酸残基还原,产生深蓝色(钼兰和钨兰的混合物)。
在一定的条件下,蓝色深度与蛋白的量成正比。
2试剂:2.1)甲液:1,4%碳酸钠;2,0.2n氢氧化钠;3,1%硫酸铜;4,2%酒石酸钾钠。
1和2,4溶于500ml水中,然后和4以50:1混合。


〔Ⅱ〕实验部分实验一蛋白质含量测定法本实验的目的是学会各种蛋白质含量的测定方法。
了解各种测定方法的基本原理和优缺点。
蛋白质含量测定法,是生物化学研究中最常用、最基本的分析方法之一。
目前常用的有四种古老的经典方法,即定氮法,双缩尿法(Biuret法)、Folin-酚试剂法(Lowry 法)和紫外吸收法。
另外还有一种近十年才普遍使用起来的新的测定法,即考马斯亮蓝法(Bradford法)。
其中Bradford法和Lowry法灵敏度最高,比紫外吸收法灵敏10~20倍,比Biuret法灵敏100倍以上。
定氮法虽然比较复杂,但较准确,往往以定氮法测定的蛋白质作为其他方法的标准蛋白质。
值得注意的是,这后四种方法并不能在任何条件下适用于任何形式的蛋白质,因为一种蛋白质溶液用这四种方法测定,有可能得出四种不同的结果。
每种测定法都不是完美无缺的,都有其优缺点。
在选择方法时应考虑:①实验对测定所要求的灵敏度和精确度;②蛋白质的性质;③溶液中存在的干扰物质;④测定所要花费的时间。
考马斯亮蓝法(Bradford法),由于其突出的优点,正得到越来越广泛的应用。
显色后会色浅,则必须提高碳酸钠—氢氧化钠溶液的浓度1~2倍。
进行测定时,加F olin—酚试剂时要特别小心,因为该试剂仅在酸性pH条件下稳定,但上述还原反应只在pH=10的情况下发生,故当Folin一酚试剂加到碱性的铜—蛋白质溶液中时,必须立即混匀,以便在磷钼酸—磷钨酸试剂被破坏之前,还原反应即能发生。
五、考马斯亮兰法(Bradford法)(一)实验原理双缩脲法(Biuret法)和Folin—酚试剂法(Lowry法)的明显缺点和许多限制,促使科学家们去寻找更好的蛋白质溶液测定的方法。
1976年由Bradford建立的考马斯亮兰法(Bradford法),是根据蛋白质与染料相结合的原理设计的。
这种蛋白质测定法具有超过其他几种方法的突出优点,因而正在得到广泛的应用。
这一方法是目前灵敏度最高的蛋白质测定法。


生化实验报告引言:生化实验是现代生物科学研究中不可或缺的一部分。
通过生化实验,可以深入了解生物体的分子组成和功能,揭示生物体的生理过程和相关疾病的发生机制。
本报告将就我们进行的一系列生化实验结果进行总结和分析。
实验一:蛋白质浓度测定实验蛋白质是生物体的重要组成部分,也是许多生理过程的关键调节因子。
通过本实验,我们采用比色法测定了不同样本中的蛋白质含量,并得出以下结论:1. 样本A的蛋白质浓度明显高于样本B,表明样本A可能含有更多的蛋白质。
2. 样本C的蛋白质浓度最低,可能是由于样本C中存在其他干扰物质,影响了测定结果。
实验二:酶活性测定实验酶是生物体内参与代谢反应的重要分子,其活性的变化可以反映生物体的生理状态。
本实验中,我们通过测定不同条件下酶的活性,得出以下结论:1. 酶活性随着温度的升高呈先增加后下降的趋势,说明酶在适宜温度下具有最高的活性。
2. pH值的变化对酶活性也有一定影响,酶活性在酸性和碱性条件下均降低,表明酶对于特定pH值有一定的适应性。
3. 离子浓度的改变可以显著影响酶的活性,高浓度的阳离子对酶活性的抑制效果较大。
实验三:DNA提取实验DNA是生物体内存储遗传信息的重要分子,在基因研究和生物技术中应用广泛。
本实验中,我们采用了几种常见的DNA提取方法,得出以下结论:1. 不同提取方法对于不同样本的DNA提取效果有所差异,需要根据实际情况选择合适的方法。
2. 样本中其他杂质的存在会干扰DNA的提取,需要进行适当的纯化步骤以提高提取纯度。
3. 所得到的DNA质量可以通过比色法或荧光法进行检测,得出相对准确的结果。
实验四:酸碱滴定实验酸碱滴定是一种常用的分析方法,在酸碱度测定和沉淀反应等方面有广泛应用。
本实验中,我们进行了一系列的酸碱滴定实验,得出以下结论:1. 强酸和强碱的滴定过程呈现出明显的酸碱中和点,通过计算可以得到溶液中目标物质的浓度。
2. 在滴定过程中,我们需要通过指示剂的颜色变化来判断滴定终点,需注意颜色的变化程度和持续时间。

实验一蛋白质分子量的测定—凝胶层析法一、原理凝胶层析法也称分子筛层析法,是利用具有一定孔径大小的多孔凝胶作固定相的层析技术。
当混合物随流动相经过凝胶层析柱时,其中各组分按其分子大小不同而被分离的技术。
该法设备简单、操作方便、重复性好、样品回收率高。
凝胶是一种不带电的具有三维空间的多孔网状结构、呈珠状颗粒的物质,每个颗粒的细微结构及筛孔的直径均匀一致,像筛子,小的分子可以进入凝胶网孔,而大的分子则排阻于颗粒之外。
当含有分子大小不一的蛋白质混合物样品加到用此类凝胶颗粒装填而成的层析柱上时,这些物质即随洗脱液的流动而发生移动。
大分子物质沿凝胶颗粒间隙随洗脱液移动,流程短,移动速率快,先被洗出层析柱;而小分子物质可通过凝胶网孔进入颗粒内部,然后再扩散出来,故流程长,移动速度慢,最后被洗出层析柱,从而使样品中不同大小的分子彼此获得分离。
若分子大小介于上述完全排阻或完全渗入凝胶的物质,则居二者之间从柱中流出。
总之,各种不同相对分子质量的蛋白质分子,最终由于它们被排阻和扩散的程度不同,在凝胶柱中所经过的路程和时间也不同,从而彼此可以分离开来。
将凝胶装在柱后,柱床体积称为“总体积”,以Vt表示。
实质上Vt是由Vo,Vi与Vg三部分组成,Vo称为“孔隙体积”或“外水体积”,即存在于柱床内凝胶颗粒外面空隙之间的水相体积,相应于一般层析法中柱内流动相的体积;Vi为内体积,即凝胶颗粒内部所含水相的体积。
Vg为凝胶本身的体积。
洗脱体积(Ve)与Vo与Vi之间的关系可用下式表示:Ve=Vo+KdVi。
式中Ve为洗脱体积,自加入样品时算起,到组分最大浓度(峰)出现时所流出的体积;Kd为样品组分在二相间的分配系数,也可以说Kd是分子量不同的溶质在凝胶内部与外部的分配系数。
它只与被分离的物质分子的大小和凝胶颗粒孔隙的大小分布有关,而与柱的长度粗细无关,也就是说它对每一物质为常数,与柱的物理条件无关。
Kd可通过实验求得,上式可以改写为:Kd=(Ve-Vo)/Vi。

蛋白质含量的测试方法蛋白质是生物体生命活动所必需的重要营养物质之一、了解食物中蛋白质的含量对于饮食调控和营养评估非常重要。
蛋白质含量的测试方法可以根据不同的样品性质和需要,选择合适的方法进行定量分析。
以下将介绍几种常见的蛋白质含量测试方法。
一、低里氏试剂法低里氏试剂法是目前最常用的蛋白质含量测试方法之一、该方法利用氢氧化钠(NaOH)/硫酸铜(CuSO4)/低里氏试剂进行蛋白质的量化分析。
具体操作步骤如下:1.将待测样品溶解于含有氢氧化钠的试液中,加入硫酸铜和低里氏试剂。
2.进行加热反应,使还原蛋白质与低里氏试剂发生比色反应。
3.通过比色计(常见的是分光光度计)测定试液的吸光度,并与标准曲线对照,计算出样品中蛋白质的含量。
二、布拉得福法布拉得福法是另一种常用的蛋白质定量方法。
该方法利用显色剂最鲁丁(Coomassie Brilliant Blue G-250)与蛋白质分子之间的相互作用来定量测定蛋白质含量。
具体操作步骤如下:1.将待测样品与显色剂最鲁丁充分混合,并保持一定时间使其反应发生。
2.使用比色计测定混合液的吸光度,通过与标准曲线对照,计算出蛋白质的含量。
布拉得福法相对于低里氏试剂法更为敏感,对大多数蛋白质都有较好的定量效果。
三、生物素结合法生物素结合法是一种利用亲和力层析技术测定蛋白质含量的方法。
该方法基于生物素和亲和层析树脂之间的结合作用,通过测定结合蛋白质与酶标记物之间的信号强度,来定量测定蛋白质的含量。
具体操作步骤如下:1.将待测样品与含有生物素键合的亲和层析树脂充分混合,并进行孵育反应。
2.经过层析分离后,将酶标记物加入,通过测定酶标记物与生物素之间的信号强度,计算出蛋白质的含量。
生物素结合法一般适用于高通量的蛋白质含量测定,具有灵敏度高、准确度高的特点。
四、生物学方法在蛋白质含量测定中,生物学方法也具有一定的重要性。
例如,利用生物学方法如酶活性测定、氨基酸组分测定等,可以间接推测蛋白质的含量。

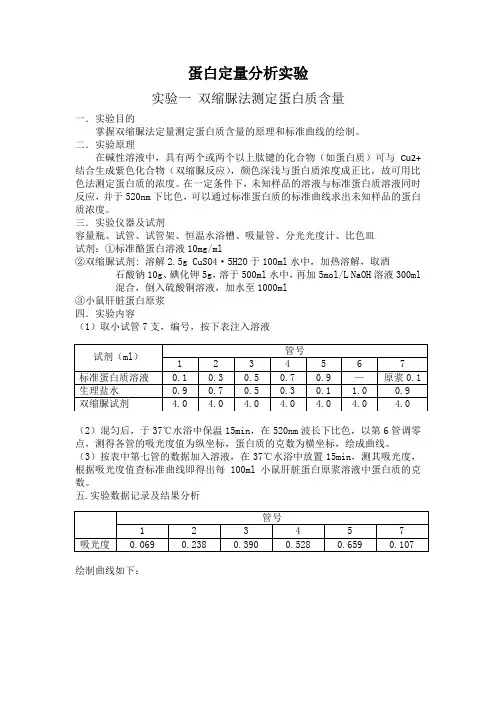
蛋白定量分析实验实验一双缩脲法测定蛋白质含量一.实验目的掌握双缩脲法定量测定蛋白质含量的原理和标准曲线的绘制。
二.实验原理在碱性溶液中,具有两个或两个以上肽键的化合物(如蛋白质)可与Cu2+结合生成紫色化合物(双缩脲反应),颜色深浅与蛋白质浓度成正比,故可用比色法测定蛋白质的浓度。
在一定条件下,未知样品的溶液与标准蛋白质溶液同时反应,并于520nm下比色,可以通过标准蛋白质的标准曲线求出未知样品的蛋白质浓度。
三.实验仪器及试剂容量瓶、试管、试管架、恒温水浴槽、吸量管、分光光度计、比色皿试剂:①标准酪蛋白溶液10mg/ml②双缩脲试剂: 溶解2.5g CuSO4·5H2O于100ml水中,加热溶解,取酒石酸钠10g、碘化钾5g,溶于500ml水中,再加5mol/L NaOH溶液300ml混合,倒入硫酸铜溶液,加水至1000ml③小鼠肝脏蛋白原浆四.实验内容(1)取小试管7支,编号,按下表注入溶液(2)混匀后,于37℃水浴中保温15min,在520nm波长下比色,以第6管调零点,测得各管的吸光度值为纵坐标,蛋白质的克数为横坐标,绘成曲线。
(3)按表中第七管的数据加入溶液,在37℃水浴中放置15min,测其吸光度,根据吸光度值查标准曲线即得出每100ml小鼠肝脏蛋白原浆溶液中蛋白质的克数。
五.实验数据记录及结果分析绘制曲线如下:由图可知,当吸光度为0.107时,相对应的蛋白质克数为1.38 mg,则100ml 小鼠肝脏蛋白原浆液中蛋白质克数为1.38 g。
实验二考马斯亮蓝法测定蛋白质含量一.实验目的掌握考马斯亮蓝法定量测定蛋白质含量的原理和方法;熟悉紫外分光光度计的使用。
二.实验原理考马斯亮蓝在游离状态下呈红色,当它与蛋白质结合后变为蓝色,在一定蛋白质浓度范围内,结合物在595 nm波长下有最大吸收峰,测其光的吸收量即可得结合蛋白质的量。
三.实验仪器及试剂仪器:分光光度计,试管,吸量管,比色皿试剂:①考马斯亮蓝试剂: 考马斯亮蓝G-250 100mg溶于50 ml 95%乙醇中,加入100ml 85%磷酸,用蒸馏水稀释至1000 ml。

蛋白质含量测定方法
首先,我们来介绍最常用的蛋白质含量测定方法之一——比色法。
比色法是通过蛋白质与某种试剂发生化学反应产生有色产物,
然后利用分光光度计测定产物的吸光度来计算蛋白质含量的方法。
常用的试剂包括布拉德福试剂、伯里氏试剂等。
比色法操作简便,
结果准确,适用于多种类型的蛋白质样品。
其次,还有一种常用的蛋白质含量测定方法是BCA法。
BCA法
是利用蛋白质与BCA试剂在碱性条件下发生还原反应,生成紫色螯
合物,然后利用分光光度计测定其吸光度来计算蛋白质含量的方法。
BCA法对于含有还原剂、胶体物质和表面活性剂的样品具有较好的
适用性,同时也具有较高的灵敏度和线性范围。
另外,还有一种常用的蛋白质含量测定方法是Lowry法。
Lowry
法是通过蛋白质与碱性铜离子和菲罗啉在碱性条件下发生络合反应,生成蓝色络合物,然后利用分光光度计测定其吸光度来计算蛋白质
含量的方法。
Lowry法对于各种类型的蛋白质样品均有较好的适用性,但操作相对复杂,需要较长的实验时间。
除了上述介绍的常用方法外,还有其他一些蛋白质含量测定方
法,如紫外吸收法、荧光法等。
这些方法各有特点,适用于不同类
型的蛋白质样品,实验人员可以根据实际情况选择合适的方法进行
测定。
总的来说,蛋白质含量的准确测定对于科学研究和实验室工作
至关重要。
在选择测定方法时,需要考虑样品的性质、实验条件、
仪器设备等因素,并且在操作过程中要严格按照方法要求进行操作,确保测定结果的准确性和可靠性。
希望本文介绍的方法能为您的实
验工作提供一些帮助,祝您实验顺利!。
实验一:蛋白质含量测定方法的研究---非蛋白物质的可能干扰及其干扰程度的研究分析摘要:本次实验通过分别利用三种方法(Folin-酚法,考马斯亮蓝(G-250),紫外法)平行测定纯蛋白溶液和生物材料中蛋白的含量,分别计算和比较每种材料通过三种方法测定结果的差异性。
再利用同时进行的11组实验的数据进行整合分析。
最后通过比较两种待测液在三种方法测定下的差异性来分析非蛋白质干扰因素对三种方法的影响及影响程度。
关键字:比色法,蛋白质含量测定,非蛋白质干扰因素,差异比较与分析1.研究背景及目的实验室测定蛋白质含量的方法多种,且依照不同的应用方向,依照不同的原理有多种蛋白质含量的测定方式。
目前蛋白质含量测定的主要应用方法是比色法,但是由于实验操作以及设备存在误差,所以蛋白质溶液的真实浓度难以知晓,因此不能直接恒量各种方法在不同测定环境中的差异性。
同时在实际的测量中,由于生物样品的复杂性,目标溶液中往往存在着诸多非蛋白干扰因素。
因此需要实验者通过多种方法,有一定的数据支持,从可依靠的数据的线性变化中寻求合理的数理参照,从而对非蛋白干扰因素的影响程度做以分析和比较。
2.原理本次实验选取三种实验方法——Folin-酚法,考马斯亮蓝(G-250)法和紫外法:Folin-酚法的原理利用是蛋白质中含有酚基的氨基酸数与蛋白质浓度呈正比,从而对特殊残基的测定来确定蛋白质含量;考马斯亮蓝法的原理是在一定蛋白质浓度范围内,蛋白质与色素结合物在595nm波长下光的吸收与浓度呈正比,故可用于蛋白质的测定;紫外法的原理是:蛋白质在紫外光下,吸收高峰在280nm左右,在此波长范围内蛋白质溶液的光密度值与浓度呈正比。
本次实验测定的对象为纯蛋白溶液(BSA,牛血清白蛋白)和生物材料(绿豆芽下胚轴蛋白的提取液),分别作为无非蛋白质干扰和非蛋白质干扰的两个实验组。
通过分别对两种溶液利用三种方法进行测定。
用数理处理和分析来衡量三种方法在测定中的波动,比较两待测液即纯蛋白和生物组织提取液之间的波动情况的差异。
实验名称:生化蛋白测定实验实验日期:2023年X月X日实验目的:1. 掌握蛋白质定量分析的基本原理和方法。
2. 熟悉分光光度计的使用和操作。
3. 通过实验,学习如何测定蛋白质浓度。
实验原理:蛋白质的定量分析是生物化学研究中不可或缺的环节。
本实验采用双缩脲法测定蛋白质含量。
该方法基于蛋白质中的肽键与铜离子在碱性条件下形成紫色络合物,其颜色深浅与蛋白质含量成正比。
通过比色法测定吸光度,从而计算出蛋白质的浓度。
实验材料与试剂:1. 实验材料:动物组织样品(如肝脏、肌肉等)。
2. 试剂:- 双缩脲试剂:A液(含碱性酒石酸铜溶液)、B液(含硫酸铜溶液)。
- 标准蛋白质溶液(已知浓度)。
- 0.1mol/L氢氧化钠溶液。
- 0.05mol/L硫酸铜溶液。
- 0.1mol/L盐酸溶液。
- 0.1mol/L磷酸盐缓冲液(pH 7.0)。
实验仪器:1. 分析天平。
2. 分光光度计。
3. 烧杯。
4. 试管。
5. 移液器。
6. 混匀器。
实验步骤:1. 样品处理:称取适量动物组织样品,用0.1mol/L氢氧化钠溶液溶解,制成蛋白质溶液。
2. 标准曲线绘制:取不同浓度的标准蛋白质溶液,加入双缩脲试剂A液,混匀,室温放置10分钟。
加入B液,混匀,在540nm波长下测定吸光度。
以蛋白质浓度为横坐标,吸光度为纵坐标,绘制标准曲线。
3. 样品测定:取待测样品,按照上述步骤进行处理,测定吸光度。
4. 计算蛋白质浓度:根据标准曲线,计算待测样品中蛋白质的浓度。
实验结果:1. 标准曲线绘制:根据实验数据,绘制标准曲线,线性回归方程为y=0.0035x+0.0146,相关系数R²=0.998。
2. 样品测定:待测样品的吸光度为0.65。
3. 蛋白质浓度计算:根据标准曲线,待测样品中蛋白质浓度为0.3mg/mL。
讨论与分析:本实验采用双缩脲法成功测定了待测样品中的蛋白质含量。
实验过程中,应注意以下几点:1. 样品处理过程中,应尽量减少蛋白质的降解。