多糖结构总结
- 格式:doc
- 大小:578.96 KB
- 文档页数:7

多糖的结构和功能的分子生物学研究多糖是一种高分子化合物,由不同的单糖分子通过碳-碳键或者碳-氧键连接而成。
多糖的结构不仅决定了它们的性质和功能,也影响了它们在生物系统中的作用和发挥。
多糖的结构研究一直是分子生物学研究的热点。
在多糖结构研究中,分子生物学的方法和技术得到了广泛的应用。
一、糖基化修饰的多糖结构多种生物大分子都会经历糖基化修饰,这是一种生物大分子表面化学修饰,涉及到蛋白质、核酸和多糖等。
糖基化修饰是多糖结构研究中一个重要的方向,它影响了多糖在细胞中的功能和分布,同时也对外界环境的变化有所响应。
以壳多糖为例,它是常见的一种多糖,存在于不同种类的细菌和真菌细胞壁中,同时也是常见的病原体。
壳多糖的结构研究发现,其糖基化修饰程度和方式的不同,可以影响到其生物活性和免疫学特性。
因此,对壳多糖的糖基化修饰的研究对于设计和生产新型抗生素和疫苗具有重要的意义。
二、多糖的三维结构解析在多糖结构研究中,三维结构的研究是另一个重要的方向。
与其他生物大分子相比,多糖较为复杂,不同的单糖子基、连接方式和伸展程度都决定了多糖的三维结构。
因此,研究多糖的三维结构就可以从原子层面了解多糖的性质和功能。
目前,多糖的三维结构研究主要通过核磁共振、X射线晶体学和电子显微镜等技术手段来完成。
例如,X射线晶体学可以解析多糖的晶体结构,提供高分辨率的空间信息。
电子显微镜则可以帮助研究人员获得多糖的三维形态,这有利于了解多糖在细胞和组织中的相互作用和变化。
三、多糖的生物学功能多糖在生物中具有多种生物学功能,例如参与免疫调节、细胞凝聚、防御外部信号等。
多糖功能的了解与其结构有着密切联系,因此研究多糖的生物学功能也是多糖结构研究的重要方向。
以纤维连接素为例,它是一种高分子化合物,存在于细胞外基质中,是细胞外支架的主要构成元素。
纤维连接素的结构研究表明,其结构的独特性决定了它对细胞外基质的组织和机械特性的影响。
同时,纤维连接素在胶原纤维和弹性纤维的修饰、不同细胞类型之间的相互作用等方面发挥着关键作用。

多糖的结构一、多糖的概念多糖是由许多单糖分子通过糖苷键连接而成的大分子化合物。
它们是生物体内重要的能量来源,也是构成细胞壁、细胞膜和组织结构的重要成分。
多糖可以分为两类,即多糖和寡糖,其中多糖由许多单糖分子组成,而寡糖则由较少的单糖分子组成。
二、多糖的结构多糖的结构非常多样,可以是直链状、分枝状或环状。
这些结构的差异主要取决于单糖分子之间的连接方式和连接位置。
1. 直链状多糖直链状多糖是指单糖分子通过糖苷键直接连接在一起,形成一条直线。
这种结构通常具有较高的溶解度和可溶性,因为这种结构可以使水分子更容易与多糖分子相互作用。
直链状多糖在生物体内起着能量储存和结构支持的作用。
2. 分枝状多糖分枝状多糖是指单糖分子通过糖苷键连接成主链,同时还有其他单糖分子通过糖苷键连接在主链上,形成分支结构。
这种结构使得多糖的空间结构更加复杂,增加了多糖的稳定性和生物活性。
分枝状多糖在生物体内具有重要的生物功能,例如细胞识别、细胞黏附和信号传导等。
3. 环状多糖环状多糖是指单糖分子通过糖苷键形成一个或多个环状结构。
这种结构使得多糖分子更加紧密和稳定。
环状多糖在生物体内广泛存在,例如淀粉和纤维素等。
它们在植物细胞壁中起着结构支持的作用。
三、多糖的功能多糖在生物体内具有多种功能,包括能量储存、结构支持、细胞识别、细胞黏附和信号传导等。
1. 能量储存多糖是生物体内重要的能量来源。
例如,淀粉是植物细胞中的能量储存物质,动物体内的糖原也是通过多糖形式储存的能量。
2. 结构支持多糖可以构成细胞壁、细胞膜和组织结构的重要成分,起到支持和保护细胞的作用。
例如,纤维素是植物细胞壁的主要组成部分,赋予植物细胞结构稳定性。
3. 细胞识别多糖具有特异的生物学活性,可以与细胞膜上的受体结合,以实现细胞间的相互识别。
这对于细胞的正常功能和生物体的正常发育至关重要。
4. 细胞黏附多糖可以通过与细胞表面的特定受体结合,促进细胞的黏附和聚集。
这对于细胞间的相互作用和组织形成至关重要。

多糖结构多糖(polysaccharide)是由多个单糖分子缩合、失水而成,是一类分子机构复杂且庞大的糖类物质。
凡符合高分子化合物概念的碳水化合物及其衍生物均称为多糖。
多糖多糖在自然界分布极广,亦很重要。
有的是构成动植物骨架结构的组成成分,如纤维素;有的是作为动植物储藏的养分,如糖原和淀粉;有的具有特殊的生物活性,像人体中的肝素有抗凝血作用,肺炎球菌细胞壁中的多糖有抗原作用。
多糖的结构单位是单糖,多糖相对分子质量从几万到几千万。
结构单位之间以苷键相连接,常见的苷键有α-1,4-、β-1,4-和α-1,6-苷键。
结构单位可以连成直链,也可以形成支链,直链一般以α-1,4-苷键(如淀粉)和β-1,4-苷键9如纤维素)连成;支链中链与链的连接点常是α-1,6-苷键。
由一种类型的单糖组成的有葡萄糖、甘露聚糖、半乳聚糖等,由二种以上的单糖组成的杂多糖(hetero polysaccharide)有氨基糖的葡糖胺葡聚糖等,在化学结构上实属多种多样。
就分子量而论,有从0.5万个分子组成的到超过106个的多糖。
比10个少的短链的称为寡糖。
不过,就糖链而论即使是寡糖,在寡糖上结合了蛋白质和脂类的,就整个分子而论,如果是属于高分子,则从广义上来看也属于多糖,因此特称为复合多糖(conjugated polysaccharide,complex poly-saccharide)或复合糖质(glycoconjugate)(糖蛋白、糖脂类、蛋白多糖)。
[1]临床作用免疫调节Hosono Akira等将双岐杆菌属细菌的细胞超声粉碎提取后,用超滤设备和阴离子交换树脂、凝胶色谱纯化出具有免疫增强活性的多糖。
Oka Shuichi等从紫苏(Perilla)中分离得到的多糖具有抗变态反应作用。
Fujimiy hjaki注射用黄芪多糖从蘑菇属(Agr/cus)植物的子实体中提取出的多糖具有免疫抑制作用,它能减少我们通常使用的免疫抑制剂的诸如细胞毒性、机体抗感染能力下降、对骨髓造血细胞的繁殖抑制等副作用,此多糖可以做成口服或注射用药物,也可制成一种功能性食品。
经过分级纯化的多糖在测定结构前须检查其纯度及测定分子量。
检查纯度最常用的判断方法:(1)用G C 、HPLC测定组成多糖的单糖的摩尔比是否恒定。
用不同的柱型测定结果更为可靠。
(2)电泳只出现一条带。
如可用聚丙烯酰胺凝胶电泳、乙酸纤维素薄膜电泳及玻璃纤维纸电泳。
对于中性多糖可采用高压电泳,以硼酸盐为缓冲液,可增大其迁移速度。
(3)凝胶柱层析图呈现对称的单峰。
若有“拖尾”现象,说明其均一性不够好。
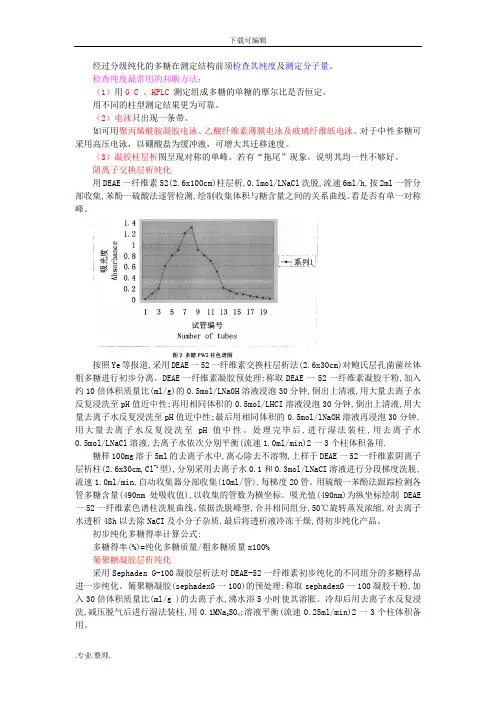
阴离子交换层析纯化用DEAE一纤维素52(2.6x100cm)柱层析,0.lmol/LNaCl洗脱,流速6ml/h,按2ml一管分部收集,苯酚一硫酸法逐管检测,绘制收集体积与糖含量之间的关系曲线。
看是否有单一对称峰。
按照Ye等报道,采用DEAE一52一纤维素交换柱层析法(2.6x30cm)对鲍氏层孔菌菌丝体粗多糖进行初步分离。
DEAE一纤维素凝胶预处理:称取DEAE一52一纤维素凝胶干粉,加入约10倍体积质量比(ml/g)的0.5mol/LNa0H溶液浸泡30分钟,倒出上清液,用大量去离子水反复浸洗至pH值近中性;再用相同体积的0.5mol/LHCI溶液浸泡30分钟,倒出上清液,用大量去离子水反复浸洗至pH值近中性;最后用相同体积的0.5mol/lNaOH溶液再浸泡30分钟,用大量去离子水反复浸洗至pH值中性。
处理完毕后,进行湿法装柱,用去离子水0.5mol/LNaCl溶液,去离子水依次分别平衡(流速1.0ml/min)2一3个柱体积备用.糖样100mg溶于5ml的去离子水中,离心除去不溶物,上样于DEAE一52一纤维素阴离子层析柱(2.6x30cm,Cl-1型),分别采用去离子水0.1和0.3mol/LNaCI溶液进行分段梯度洗脱,流速1.0ml/min,自动收集器分部收集(10ml/管),每梯度20管。
用硫酸一苯酚法跟踪检测各管多糖含量(490nm处吸收值),以收集的管数为横坐标。
吸光值(490nm)为纵坐标绘制DEAE 一52一纤维素色谱柱洗脱曲线。



多糖(polysaccharides)=聚糖(glycans)第一节序言多糖具有储存能量、结构支持、防御等功能;80 年代又发现其可控制细胞的分裂和分化,调节细胞的生长和衰老。
近年发现糖及其缀合物是细胞识别的主要标记物,在细胞间物质运输、信号传导、免疫功能调节等方面都有相当重要的作用。
第二节多糖及其分类与结构一、定义:十个以上单糖聚合而成的糖属于多糖,DP (degreepolymerization):10-105。
二、分类:分植物多糖和动物多糖,又从来源、功能和化学结构分类。
三、结构1.化学化学结构分类简单多糖结合多糖(蛋白多糖)均多糖(homosaccharides) 杂多糖(heterosaccharides) 直链多糖(liner PS) 支链多糖(branch PS)2.表示方法均多糖:glucan fructan xylan 杂多糖:galactomamnan glucomannan3.糖的组成Gal、glc、xyl、ara、rha、fru、fuc,rib(核糖)mannose(甘露糖)糖醛酸:如Glucuronic acid = glu A 去氧糖、氨基糖、糖醇、酰基糖、磺酰酯糖、磷酸基糖4. 四级结构一级:糖的组成;(种类,glc, xyl......)糖的构型(ɑ、ß、D、L)连接方式(连接位置、支链、直链)连接顺序二级:以氢键结合的聚合体(糖骨架间)三级:一级结构重复顺序(有规则)四级:糖链间以非共价键结合形成聚集体的立体结构可拉伸的带状结构皱纹型带状结构屈曲状螺旋结构曲屈线圈状结构第三节多糖的提取、纯化和分离方法一、提取方法1.易溶于热水的多糖:90-100℃水提三次,也有用盐水提; 浓缩后加EtOH沉淀。
2.难溶于水,可溶于稀碱液的多糖:0.5N NaOH提两次,酸中和沉淀。
酸性糖?3.糖复合物:与蛋白质形成的糖复合物,需断裂糖和蛋白质的结合,常用的断裂法有碱解法和酶解法。

一:多糖中的单糖组分分析一般对多糖进行完全水解,水解条件:封管0.5~3M硫酸或1~6M盐酸,80℃~100℃水解2.5~8h 即可。
或控制水解条件,进行逐步水解,如封管0.025M硫酸,100℃水解15min,30min,45min 等,水解液用碳酸钡或氢氧化钡中和,滤液浓缩后可用纸层析、薄层层析、气相层析或高压液相层析等鉴定。
二:相邻单糖基连接方式分析将甲基化多糖水解得到甲基化的单糖,而此单糖上甲基化之羟基所在的碳原子就是连接键所在。
高碘酸氧化是定量反应,Smith降解是将高碘酸氧化产物进行还原,酸水解或部分水解,从高碘酸的消耗量和不同产物的生成,便可进行糖苷键位置的判断-产物中若有一分子比例的甲酸生成而消耗两分子比例的高碘酸根时,表明多糖的非还原末端或非末端部分有1-6苷键相连的单糖基存在;产物中若有赤藓醇生成,则提示有1-4结合苷键;若有甘油生成,有1-6、1-2结合的苷键或有还原性末端葡萄糖基等;若产物中能检出单糖,如葡萄糖、半乳糖、甘露糖等,则有1-3苷键存在。
结合¹³C-NMR确定连接位置。
三:端基碳苷键构型分析1:酶解实验:不被淀粉酶水解的多糖,无α-苷键,与纤维素酶有作用者,存在β-苷键。
2;IR:α-型差向异构体的C-H键在844±8cm‾¹处有一个吸收峰;β-型的C-H键在891±7cm‾处有一个吸收峰。
但是,海藻糖、阿洛糖和异阿洛糖的α-型和β-型同时存在的情况下,就不能以次来判断。
3:¹H-NMR:端基碳的δ值大于5.00ppm者,糖苷键为α-型,小于5.00ppm者,则为β-型。
4;¹³C-NMR:α-型连接的C₁化学位移在97-101ppm,β-型的在103~105ppm。
对甘露聚糖不能用化学位移判断α-型或β-型。
可用裂分常数决定,一般¹Jc-h=170HZ,为α-型,160HZ 者为β-型。

糖类化学知识点总结糖类是一类重要的有机化合物,其化学结构和性质的研究对于生物学和食品工业具有重要的意义。
糖类包括单糖、双糖、多糖等多种类型,它们具有不同的分子结构和特性。
本文将对糖类的化学结构、命名方法、性质以及在生物体内和食品工业中的应用进行系统的总结和阐述。
一、单糖的化学结构和命名方法1. 单糖的分类单糖是由碳、氢、氧三种元素组成的糖类化合物,它们的分子结构中含有一个或多个羟基和一个或多个醛基或酮基。
根据它们的化学结构,单糖可分为醛糖和酮糖两类。
醛糖的分子中含有一个醛基,酮糖的分子中含有一个酮基。
2. 单糖的化学结构单糖的化学结构可以用希尔德-奥斯特公式来表示,其中n代表碳原子数,希尔德-奥斯特公式的结构为(CH2O)n。
单糖的分子结构包括直链结构和环状结构两种形式。
直链结构是单糖分子直接相连形成的链状结构,而环状结构是由直链结构转变而来的,其中含有环氧醇化合物。
3. 单糖的命名方法根据单糖分子中羟基的位置不同,可以分为各种不同的单糖,比如葡萄糖、果糖、半乳糖等,并且还可以根据立体构型的不同将它们分为L-型和D-型两种立体异构体。
二、双糖和多糖的化学结构和性质1. 双糖的化学结构和性质双糖是由两个单糖分子通过糖苷键连接而成的化合物,根据单糖分子的组成不同,双糖可分为蔗糖、麦芽糖、乳糖等多种类型。
双糖具有不同的甜度和溶解度,它们在食品工业中具有广泛的应用。
2. 多糖的化学结构和性质多糖是由多个单糖分子通过糖苷键连接而成的化合物,它们的分子结构复杂,包括淀粉、纤维素、半乳聚糖等多种类型。
多糖在生物体内具有重要的功能,如淀粉是植物体内储存能量的重要物质,而纤维素是植物细胞壁结构的主要组成部分。
三、糖的生物合成和降解1. 糖的生物合成糖类在生物体内是通过一系列酶促反应进行合成的,主要包括糖异生和糖原合成两个过程。
糖异生是指通过葡萄糖及其衍生物的代谢途径来合成其他单糖,而糖原合成是指通过多糖合成反应来合成淀粉和糖原。

多糖结构通式多糖是由许多单糖分子通过糖苷键连接而成的高分子化合物,是生物体内最常见的一种生物大分子化合物,包括淀粉、纤维素、糖原、聚果糖、琥珀酸、卡拉胶等等。
多糖在生物体内具有重要的生物学功能,例如作为能量的储存和释放、细胞骨架构建、免疫调节等。
多糖通常具有复杂的结构,其结构特点决定了其生物学功能。
下面我们就来详细介绍多糖的结构通式。
一、单糖单元结构多糖的结构由单糖单元组成,因此首先需要了解单糖的结构。
单糖是最基本的碳水化合物单体,是由一个或多个羟基(-OH)和一个碳羰基(C=O)组成的分子,通式为(CH2O)n。
在生物体内最常见的单糖有葡萄糖、果糖、半乳糖、甘露糖、半乳糖等。
葡萄糖的通式为C6H12O6,是一种六碳醛酮糖。
其结构中含有6个碳原子,12个氢原子,6个氧原子。
葡萄糖的化学式为C6H12O6,它存在于许多物质中,包括蜂蜜、蔬菜、水果和果汁中,也是淀粉、糖原、纤维素等多糖的基本单元。
二、多糖的通式结构多糖的通式结构通常由单糖单元通过不同的糖苷键连接而成。
糖苷键是单糖单元之间通过缺氧的碳原子和羟基结合形成的共价键。
根据糖苷键的不同连接方式,多糖可以分为直链多糖、支链多糖和分枝多糖。
1. 直链多糖的通式结构直链多糖的单糖单元通过直链方式逐个连接而成。
以淀粉为例,其结构通式如下:(葡萄糖)n(葡萄糖)n代表多个葡萄糖单元通过α-1,4- 糖苷键连接而成的直链多糖结构。
淀粉是植物细胞内的主要能量储备物质,其直链结构决定了其在体内被酶水解时的特定生物学功能。
2. 支链多糖的通式结构支链多糖的单糖单元通过主链和支链的方式连接而成。
以聚果糖为例,其结构通式如下:(葡萄糖)n-(葡萄糖)m(葡萄糖)n代表主链上多个葡萄糖单元的直链结构,(葡萄糖)m代表侧链上少量葡萄糖单元的支链结构。
聚果糖在植物体内具有结构支持和生长调节的重要功能。
3. 分枝多糖的通式结构分枝多糖具有着不同位置和数量的支链,其结构更为复杂。

多糖结构研究方法多糖及其复合物是来自于高等动、植物细胞膜和微生物细胞壁中的天然大分子物质之一,自然界含量丰富,与人类生活紧密相关,对维持生命活动起至关重要的作用。
多糖和核酸、蛋白质、脂类构成了最基本的4类生命物质。
由于多糖的生物活性与多糖的结构关系密切,因此清楚认识多糖的结构是进行多糖研究和利用的基础。
多糖结构比蛋白质和核酸的结构更加复杂,可以说是自然界中最复杂的生物大分子。
从化学观点来看,多糖结构解析最大的难点就在于其结构的复杂性。
糖的结构分类可沿用蛋白质和核酸的分类方法,即多糖的结构也可分为一级、二级、三级和四级结构.与蛋白质或核酸大分子相比,糖链的一级结构“含义”要十分丰富。
测定糖链的一级结构,要解决以下几个问题:(1)相对分子质量;(2)糖链的糖基组成,各种单糖组成的摩尔比;(3)有无糖醛酸及具体的糖醛酸类型和比例;(4)各单糖残基的D-或L.构型,毗喃环或呋喃环形式;(5)各个单糖残基之间的连接顺序;(6)每个糖苷键所取的a—或B.异头异构形式;(7)每个糖残基上羟基被取代情况:(8)糖链和非糖部分连接情况;(9)主链和支链连接位点:(10)糖残基可能连接硫酸酯基、乙酰基、磷酸基、甲基的类型等.多糖的二级结构是指多糖主链间以氢键为主要次级键而形成的有规则的构象,与分子主链的构象有关,不涉及侧链的空间排布;多糖的三级结构和四级结构是指以二级结构为基础,由于糖单位之间的非共价相互作用,导致二级结构在有序的空间里产生的有规则的构象四。
多糖结构的分析手段很多。
不仅有仪器分析法,如红外、核磁共振、质谱等,还有化学方法,如完全酸水解、部分酸水解、高碘酸氧化、Smith降解、甲基化反应等,以及生物学方法,如特异性糖苷酶酶切、免疫学方法等。
1质谱(MS)由于MS法在糖链结构分析中具有快速灵敏,样品用量少、结构信息直观的特点而得到越来越广泛的应用。
近年来各种软电离技术的诞生,如快原子轰击质谱(FAB-MS),电喷雾质谱(ESI-MS),基质辅助激光解析离子化质谱(MALDI —MS)等,使得糖结构分析的研究取得了日新月异的发展。
第一讲多糖概述多糖(Polysaccharides)是自然界含量最丰富的物质之一,广泛存在于动物细胞膜、植物和微生物细胞壁中,是与人类生活紧密相关的一类生物大分子,对维持生命活动起着重要的作用。
一、多糖的研究历程核酸、蛋白质和多糖并称为生命三大物质。
核酸、蛋白质研究进程显著快于多糖。
(说明原因)1.生物体内糖的研究历程:19世纪:认为多糖是能量物质,主要研究多糖的代谢与转化途径;20世纪:认为多糖是机体的结构物质,主要研究细胞外的基质成分;21世纪:认为多糖是机体的信息物质,主要研究细胞的信号转导。
2.国内外的研究现状及研究计划美国:1986年,“糖库计划”;2001年,“功能糖组学研究计划”,研究蛋白质/糖链—细胞通讯。
日本:1991年,“糖工程前沿计划”;2002年,“糖链功能1000”计划,研究糖链功能/糖类药物。
欧盟:1994年“欧洲糖类研究开发网络”;“欧盟第四、五、六框架”,研究糖链结构与功能。
中国:跟踪性研究和探索生研究;急需国家重大科技计划支持。
作为三种生物大分子之一,糖类的研究工作和蛋白质、核酸的研究工作相比,在我国还是一个薄弱环节。
现在国际上多糖研究以日本、美国、德国、加拿大处于领先地位,我国多糖的研究起步较晚,经过一个相对寂静的时期之后,自80年代各地的研究如雨后春笋般掀起。
3.糖链与生理病理的关系(1)与生理、病理的关系在正常生命过程中,多糖参与细胞分化、胚胎发育和免疫应答;在病理过程中,多糖参与癌变、感染过程。
(2)在分子内及分子间的作用分子内:影响蛋白质的折叠、半衰期,并对蛋白质有监护作用。
分子间:具有细胞识别、抗原性和信号转导作用。
二、多糖分类1.根据组成单糖的类别,多糖可分为:(1)均聚多糖(homopolysaccharides):指由同一种单糖组成,如淀粉(starch)、纤维素(cellulose)。
(2)杂多糖(heteropolysaccharides):杂多糖则是由多种单糖组成,如树胶(gums)、粘胶多糖(mucillages)、果胶(pectins)、半纤维素(hemicelluloses)等。
多糖结构里→的含义糖化学
多糖结构中的箭头(→)通常用于表示糖分子之间的连接方式或者糖链的结构。
在糖化学中,多糖是由多个单糖分子通过糖苷键连接而成的大分子化合物。
箭头可以用来描述不同单糖分子之间的连接方式,例如α-葡萄糖和β-葡萄糖通过1,4-糖苷键连接形成淀粉的结构可以用箭头来表示。
另外,箭头也可以用来表示多糖分子的空间结构,比如在多糖的分支结构中,箭头可以指示分支的位置和连接方式。
总之,在糖化学中,箭头在多糖结构中扮演着非常重要的角色,它可以帮助我们理解多糖分子的结构和性质,对于研究多糖的生物学功能和应用具有重要意义。
多糖结构分析多糖在生物学上的重要意义,尤其是在医药学上的重要意义决定了多糖研究的迅速发展,多糖构效关系的研究已成为多糖研究的热点。
但由于多糖结构的复杂性和多样性,其结构测定远远落后于蛋白质和核酸,本实验选择天然多糖(半乳葡萄甘露聚糖)作为实验材料,对其一级结构做初步的分析。
多糖一级结构的分析包括:纯度鉴定,分子量测定,单糖组成测定和糖链的序列测定。
糖链的序列测定包括:单糖残基在糖链中的次序,单糖残基间连键的位置,链的分支情况等诸多方面。
【实验目的】1.了解多糖结构分析的内容及方法。
2.了解多糖一级结构分析的基本原理。
3.掌握多糖一级结构分析的基本方法。
一、糖含量测定【实验原理】苯酚—硫酸试剂与游离的或寡糖、多糖中的己糖、糖醛酸起显色反应,己糖在490nm 处有最大吸收,吸收值与糖含量呈线性关系。
【实验材料】1. 实验器材721型分光光度计。
2. 实验试剂(1)98%的浓硫酸。
(2)80%苯酚:80g苯酚加20ml水使之溶解,可置冰箱中避光长期贮存。
(3)6%苯酚:临用前用80%苯酚配制。
(4)标准葡萄糖溶液(0.1 mg/ml):取100mg葡萄糖,用蒸馏水溶解,定容至1L。
(5)多糖样品:半乳葡萄甘露聚糖溶液(0.1 mg/ml)。
【实验操作】1. 制作标准曲线:取9支干燥试管,按下表操作横坐标为多糖微克数,纵坐标为光密度值,绘制标准曲线。
2. 样品含量测定:取样品液1.0ml,按上述步骤操作,测光密度。
3.计算:糖含量(%)=C /(C0× V)×100%C: 由标准曲线查得的糖微克数C0:样品溶液的浓度(0.1 mg/ml)V:测定时用的样品溶液体积(1.0ml)二、单糖组成分析【实验原理】多糖在浓硫酸中保温一定时间可完全水解为单糖,通过纸层析分离,特定试剂显色后与已知糖的标准混合物作对比,可以鉴定多糖水解产物中单糖的组成。
【实验材料】1. 实验器材水解管;滤纸;玻璃毛细管;层析缸;喷雾器。
多糖结构通式-回复题目:多糖结构通式的探索与解析引言:多糖是一类由大量单糖分子通过糖苷键连接而成的生物大分子。
在生物体内起着重要的结构和功能作用。
多糖具有丰富的结构多样性,其中常见的多糖有淀粉、纤维素、壳多糖等。
通过研究多糖的结构通式,我们可以更加深入地了解多糖的性质和功能,为生物大分子的研究提供重要的理论基础。
第一部分:多糖的基本概念及分类1. 多糖的定义和特性;2. 多糖的分类原则和流行分类法。
第二部分:多糖结构通式的构建1. 单糖的组成成分和结构特点;2. 糖苷键的形成机制;3. 多糖链的形成和结构。
第三部分:多糖结构通式的解析方法1. X射线晶体学;2. 核磁共振技术;3. 质谱和红外光谱技术。
第四部分:多糖结构通式的意义和应用1. 与生物体内其他组分的相互作用;2. 在药物载体和传递系统中的应用;3. 多糖的生物活性和生理功能。
结论:多糖结构通式的研究和解析对于理解多糖的性质和功能具有重要意义。
通过针对不同多糖结构的解析,我们可以更好地理解多糖的构建、功能以及与其他生物分子之间的相互作用。
未来,进一步的研究将进一步揭示多糖结构与功能之间的关系,为多糖类药物和生物医学领域的发展提供重要的理论基础和应用前景。
参考文献:1. Cantarel BL, Coutinho PM, Rancurel C, et al. The Carbohydrate-Active EnZymes database (CAZy): an expert resource for Glycogenomics[J]. Nucleic acids research, 2008, 37(Database issue): D233-D238.2. Varki A. Glycan-defined subclasses of cell surface molecules[J]. Trends in biochemical sciences, 1994, 19(6): 235-240.3. Stanley P, Schachter H, Taniguchi N. N-Glycans[M]. Cold SpringHarbor Laboratory Press, 2009.4. Jimenez-Barbero J, Gerothanassis I P, Tvaroska I. NMR in Glycoscience and Glycotechnology[M]. Royal Society of Chemistry, 2017.5. Yang B, Yu G, Zhao X. A perspective on the role of molecular dynamics in structural glycoscience[J]. Topics in Current Chemistry, 2018, 376(5): 41.。
多糖结构总结.
IR红外分析()1
的数据和图形对比可以看出,亚硒酸根[图1(a)]从硒化壳聚糖[图1(b)]与壳聚糖主要是由以下的光谱图形和光谱数据C的羟基上,主要连接在C的氨基本上和62-1变为C的氨基硒化后,NH的弯曲振动由1594.52cm变化得到证明:壳聚糖2-1为基的酰的干未基位C聚1523.29cm,壳糖氨上脱净乙基羰振动峰2
-1,而硒化壳聚糖C位上未脱干净的乙酰基1650.32cm的羰基振动峰为2-1,可能是受到C位的羟基上亚硒酸基的影响;同样由于硒化壳聚糖1632.88cm6C位氨基上和C位羟基上亚硒酸根的影响,壳聚糖C-O伸缩振动峰由62-1-1-1处观察到亚硒酸酯的800.00cm1090.41cmSe=O1079.45cm。
同时,在变为双键的振动峰。
上述红外分析结果表明:壳聚糖与亚硒酸可能是通过C位上的6酯化反应和C位上氨基的静电作用完成的。
(硒化壳聚糖的制备及其表征)
2
的对比中可以图4、从羧甲基壳聚糖与硒化羧甲基壳聚糖的红外光谱图图3主要由以下光谱图形C的羟基上。
看出, 亚硒酸根主要连接在C位的羧甲基和62-1反对称吸收峰在羧甲基壳聚糖: 1627cm-COOH处的和光谱数据变化得到证明-1
与亚1599cm-COOH, 这可能是羧甲基壳聚糖中的硒化羧甲基壳聚糖中红移至-1伸缩振动在硒化羧甲基壳处的C-O1119cm硒酸钠发生反应, 从而使键力削弱。
-1在硒化羧上的羟基也参与了硒化反应。
此外, 聚糖中红移至1064cm, 说明
C6-1(硒化羧806.125cm甲基壳聚糖的红外光谱中观测到位于双键振动峰。
的Se=O 甲基壳聚糖的合成及表征) 2.X-射线衍射,X射线衍射法是研究多糖的
结晶构型的有效方法。
多糖通常是不能结晶的但在适宜的条件下,它可以微晶态存在。
所以进行衍射分析的样品必须通过外界的诱导使其中相当部分呈现微晶态。
进行衍射的香菇多糖样品一般先制成碱进一步处理制备。
孙艳等将从香菇中分离
而得的多糖经,性溶液,然后在水中透析a=b=1.
晶格为六角形确定其立体结构为右手心三度螺旋衍射分析X2,,, 晶格常数
5nm, c =0. 6nm。
ZhangP等经X-衍射分析表明:天然香菇多糖具β三股绳
状螺旋型立体结构,但加入尿素或二甲亚砜后立体构型改变,转变为单绳螺旋结
构。
(香菇多糖结构分析和构效关系研究进展)
3.拉曼光谱法
拉曼光谱在检测多糖分子的振动相同原子的非极性键和异头物方面效果较好。
它侧重于探测多糖生物大分子的空间结构,如平铺折叠或螺旋状等。
研究
-1-1926cm954和有很强的拉曼吸收,此外在-D 表明,α螺旋直链淀粉在
865cm-1内对多糖的类500-1500cm有C-O-C 糖苷键的伸缩振动吸收,拉曼
光谱在处
型和糖苷的连接方式的检测灵敏,比红外光谱表现出了更高的分辨率,许多复杂-1区域内。
的拉曼吸收谱带都在低于600cm
2.1 Seleno-LP的拉曼光谱
-1-1附近的吸收峰亚硒酸酯中和Seleno-LP的激光拉曼光谱在
911cm699cmSe=O和Se-OH的伸缩振动,而LP在这两处均没有吸收峰。
这证实了seleno-LP中存在Se=O键。
(兰州百合多糖硒酸酯的合成及表征)
4.热分析 3.1 Seleno-LP的热分析
图3 为LP and seleno-12的差热图谱,曲线纵坐标为试样与参比物的温度差(AT)。
从图3可以看出,两种样品的差热图谱具有一定的相似性和相异性,二者表现出了3个明显的放热峰和2个明显的吸热峰。
差别在第一个峰,seleno-LP的温度要明显低于LP。
Seleno-LP的2个吸热峰峰顶温度和前2个放热峰都要低于LP的放热峰和吸热峰峰顶温度。
这与seleno-LP热稳定性低于LP的热稳定性相一致。
and ℃以前主要是LP seleno-LP的热失重图谱,在100为图4LP and
℃113seleno-LP中吸附水和结晶水的失重,两者差别不是很明显。
温度上升到开始分解,由于酯的量比较少,所以在很短的时间内完全分解。
8eleno-LP 时,引起的原因主要是多糖长都开始分解。
和seleno-LP当温度上升到246℃时,LP%和89.9℃时的失重百分数70085.9%,在链的断裂。
LP和
seleno-LP480℃和的起始分解温度是LP℃,%,且98.0seleno-LP的起始分解温度为113%,91.6 的差,而形成酯是其重要的原因。
的稳定性比℃,可以看出240seleno-LPLP
UV)5.紫外光谱
(
中,在的紫外光谱有所不同。
图1(B)LP从图1可以看出和Seleno-LP的吸收峰基本相吻A=335nmA=338nm处有一个强的吸收峰,与文献报道硒在合,紫外结果表明合成物中含有硒。
(兰州百合多糖硒酸酯的合成及表征)。