Developing a head for energy sensing AMP-activated
- 格式:pdf
- 大小:343.24 KB
- 文档页数:9
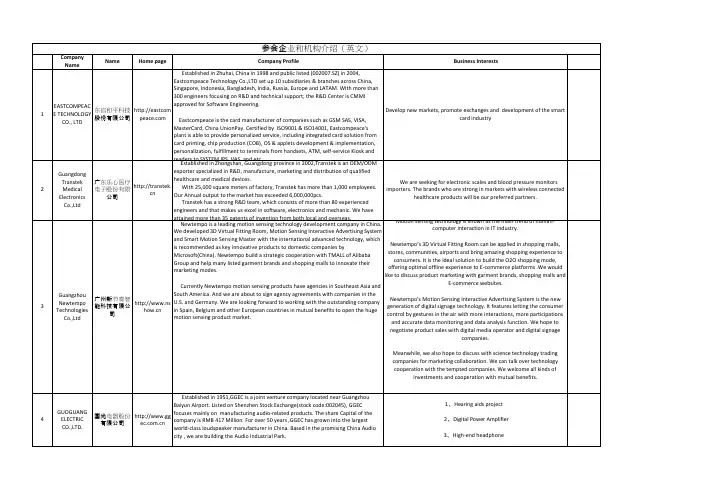
CompanyNameName Home page Company Profile Business Interests1EASTCOMPEACE TECHNOLOGYCO., LTD东信和平科技股份有限公司http://eastcomEstablished in Zhuhai, China in 1998 and public listed (002007.SZ) in 2004,Eastcompeace Technology Co.,LTD set up 10 subsidiaries & branches across China,Singapore, Indonesia, Bangladesh, India, Russia, Europe and LATAM. WIth more than300 engineers focusing on R&D and technical support; the R&D Center is CMMIapproved for Software Engineering.Eastcompeace is the card manufacturer of companies such as GSM SAS, VISA,MasterCard, China UnionPay. Certified by ISO9001 & ISO14001, Eastcompeace'splant is able to provide personalized service, including integrated card solution fromcard printing, chip production (COB), OS & applets development & implementation,personalization, fulfillment to terminals from handsets, ATM, self-service Kiosk andreaders to SYSTEM IPS, VAS, and etc.Develop new markets, promote exchanges and development of the smartcard industry2GuangdongTranstekMedicalElectronicsCo.,Ltd广东乐心医疗电子股份有限公司http://transtek.cnEstablished in Zhongshan, Guangdong province in 2002,Transtek is an OEM/ODMexporter specialized in R&D, manufacture, marketing and distribution of qualifiedhealthcare and medical devices.With 25,000 square meters of factory, Transtek has more than 1,000 employees.Our Annual output to the market has exceeded 6,000,000pcs.Transtek has a strong R&D team, which consists of more than 80 experiencedengineers and that makes us excel in software, electronics and mechanic. We haveattained more than 35 patents of invention from both local and overseas.We are seeking for electronic scales and blood pressure monitorsimporters. The brands who are strong in markets with wireless connectedhealthcare products will be our preferred partners.3GuangzhouNewtempoTechnologiesCo.,Ltd广州新节奏智能科技有限公司http://www.nsNewtempo is a leading motion sensing technology development company in China.We developed 3D Virtual Fitting Room, Motion Sensing Interactive Advertising Systemand Smart Motion Sensing Master with the international advanced technology, whichis recommended as key innovative products to domestic companies byMicrosoft(China). Newtempo build a strategic cooperation with TMALL of AlibabaGroup and help many listed garment brands and shopping malls to innovate theirmarketing modes.Currently Newtempo motion sensing products have agencies in Southeast Asia andSouth America. And we are about to sign agency agreements with companies in theU.S. and Germany. We are looking forward to working with the outstanding companyin Spain, Belgium and other European countries in mutual benefits to open the hugemotion sensing product market.Motion sensing technology is known as the main trend of human-computer interaction in IT industry.Newtempo’s 3D Virtual Fitting Room can be applied in shopping malls,stores, communities, airports and bring amazing shopping experience toconsumers. It is the ideal solution to build the O2O shopping mode,offering optimal offline experience to E-commerce platforms .We wouldlike to discuss product marketing with garment brands, shopping malls andE-commerce websites.Newtempo’s Motion Sensing Interactive Advertising System is the newgeneration of digital signage technology. It features letting the consumercontrol by gestures in the air with more interactions, more participationsand accurate data monitoring and data analysis function. We hope tonegotiate product sales with digital media operator and digital signagecompanies.Meanwhile, we also hope to discuss with science technology tradingcompanies for marketing collaboration. We can talk over technologycooperation with the tempted companies. We welcome all kinds ofinvestments and cooperation with mutual benefits.4GUOGUANGELECTRICCO.,LTD.国光电器股份有限公司http://www.ggEstablished in 1951,GGEC is a joint venture company located near GuangzhouBaiyun Airport. Listed on Shenzhen Stock Exchange(stock code:002045), GGECfocuses mainly on manufacturing audio-related products. The share Capital of thecompany is RMB 417 Million. For over 50 years ,GGEC has grown into the largestworld-class loudspeaker manufacturer in China. Based in the promising China Audiocity , we are building the Audio Industrial Park.1、H earing aids project2、D igital Power Amplifier3、H igh-end headphone参会企业和机构介绍(英文)5ShenzhenKeDao DigitalTechnologyEngineeringCo., Ltd深圳市科道数字技术工程有限公司http://www.i-Shenzhen KeDao Digital Technology Engineering Co., Ltd. was founded in 2004.Over the past ten years, we focused on the intelligent control products’ R&D,production and sales. I-tone is known as a state-level Hi-tech enterprise withcomplete independent intellectual property rights. Now, we have three series ofproducts: the Intelligent Building, Intelligent Office and Smart Home. Above all, theSmart Home series product occupies more than 30% of the domestic smart homemarket. After years of technology break through and market testing, we became theChina's top brand in the smart home. I-tone’s products guarantee the most comfortand secure experience for home & property via our humanized design and privacyassurance. Nowadays, I-tone has launched "Life Square ", "Life Cube" products withinsmart home series. We also offer the "intelligent building management system”,which provides a clear marketing direction for Chinese intelligent building andintelligent household industry. I-tone has huge technical reserves and elite marketingteam. In the future, I- tone will continuously maintain the enterprising, unremittingexploration and developing new generation of products each year to market in orderto meet the needs of all kinds of consumer groups and establish the real Chineseintelligent community network.1、Seeking overseas agent for our intelligent home product2、to cooperate with abroad advanced intelligent control technology6CANCELLEDZhuhai FounderPCBDevelopmentLimited.珠海方正印刷电路板发展有限公司www.founderpFounder PCB, established in 1986 in Zhuhai, is one of the leading PCBmanufacturers in China with 6 factories (Labeled F1-F6) and 1 PCB R&D Institution,with annual production capacity of over 15 million square feet. We are dedicated toprovide High-Tech PCB products which included HDI PCBs, Multilayer PCBs (up to 40layers), High Speed Backplanes, Line Card PCBs, Gold Finger PCBs and IC Substrate.We provide QTA/NPI services that can then transition to volume manufacturing.Exploring perspective European market expansion opportunities,technological cooperation and foreign investment intention.7Zhuhai SeineTechnologyCo., Ltd.珠海赛纳打印科技股份有限公司www.pantum.comZhuhai Seine Technology Co., Ltd is developed from Zhuhai Ninestar TechnologyCo., Ltd., which was founded in 2000. Seine is currently one of the world’s biggestgeneral printer consumable manufacturer and professional in research, developmentand manufacture. Its products have been selling to over 100 countries and areasworldwide. Also, Seine is the only laser printer manufacturer that has completelyproprietary technology in China by now, which fills the laser printer technology blankin China. With the investment in 2007, Legend Capital became a strategic partner ofSeine. In both 2011 and 2012, Seine ranked 4th in Forbes China Best SMEs.Learn about the economics development situations and requirements,especially the distribution and sales conditions of the printer consumablesmanufacturers and printer manufacturers; Expect to negotiate with localgovernmental agencies and famous local technological companies.8SHENZHEN BAKBATTERYCO.,LTD深圳市比克电池有限公司http://www.baFounded in August 2001, China BAK Battery, Inc. is listed on NASDAQ withregistered capital of 87 million US dollars. China BAK Battery, Inc. is a leading globalmanufacturer of lithium-based battery cells, with production capacity of 1.5 millionpieces per day and annual sales revenues of RMB 1.5 Billion. Our main productsinclude cylindrical, prismatic and polymer battery cells, which are the principalcomponent of rechargeable batteries commonly used in cellular phones, notebookcomputers and portable consumer electronics such as digital media devices, portablemedia players, portable audio players, portable gaming devices, PDAs, EVs ,solar,wind energy, base station and etc.. We supply the security and efficient solution forpower source to so many famous brands such as HP, Dell, lenovo, ZTE, TCL, Huawei,Chery, Nissan, Yutong, FAW,etc.The Company emphasizes the introducing of talents and technology. There arethree research centers located respectively in Shenzhen, Tianjin, and Vancouver,Canada, and a national postdoctoral work station in Shenzhen. We have the mostadvanced equipments to achieve more than 30 types of simulation experiments forreliability and safety of the products and use analytical data by 12 experimentalstations and 80 acquisition channels to produce new products and develop newmaterials. The quality management system in the company strictly abides to thestandards of ISO9000 and ISO14000 and our products are CE, UN and UL certified.The sales market includes Europe, North America, South America, Southeast Asia,Taiwan and Korea as well as China and major customers include HP, Dell, and Lenovo,ZTE, TCL, Chery, Taiwan Yulong and Yutong.Communication with a true heart and always be the best is our business purpose.We wish to use truly communication, uninterrupted innovative products andenhanced service to achieve a win-win scheme with all customers.Cooperation with the main and new energy, manufacturing, electricvehicle related industries and enterprises to carry out technicalcooperation.9Shenzhen GasCorporationLtd.深圳市燃气集团股份有限公司http://www.szgFounded in 1982, Shenzhen Gas Corporation was restructured as an enterprisewith Chinese and foreign joint investment via international bidding in 2004,integrated into a corporation in 2007 with 30-year franchising rights for pipeline gas,and listed on Shanghai Stock Exchange in 2009. Specialized in the urban pipeline gassupply, LP-gas wholesale, bottled LP-gas retailing and gas investment, it is the leadingenterprise engaged in gas supply in Shenzhen.Communicate with the foreign advanced enterprises on the gas safetymanagement experience and new technology, seek for technicalinnovation and cooperation; learn about the latest information of theinternational new energy market, and accelerate the steps of utilizing anddeveloping new energy.10Shenzhen GasInvestmentCompany深圳市燃气投资有限公司http://www.szgEstablished in 2005, Shenzhen gas Investment Corporation is a wholly-ownedsubsidiary of Shenzhen Gas Corporation with registered capital of RMB 1 Billion. It isengaged in investment, development and management for gas industry projects outof Shenzhen.After 9 years of development, Shenzhen Gas Investment Corporation has totallyinvested in 33 Independent Corporate Enterprises in 7 provinces (AutonomousRegions)include Jiangxi, Anhui, Shandong, Jiangsu, Guangdong and Inner Mongolia.So far, Shenzhen Gas Investment Corporation owns RMB 3.41 Billion of total assets,RMB 1.44 Billion of net assets and 400,000 of gas users.Communicate with the foreign advanced enterprises on how to improvethe technology and expand the market for the comprehensive utilizationof gas, including LPG and LNG, introduce advanced managementexperience, and seek for technical innovation and cooperation.11ShenzhenHazardousWasteTreatmentStation Co.,Ltd.深圳市危险废物处理站有限公司http://www.sz/Sponsored by Shenzhen Municipal Government, Shenzhen Hazardous WasteTreatment Station Co., Ltd. (SHWTS) was established in April 1988. It is the firstprofessional institution specializing in treatment and disposal of hazardous wastes inChina. In 2006, SHWTS was transformed into a state-owned enterprise, which is nowco-managed by Shenzhen Investment Holdings Co., Ltd. and Human Settlements andEnvironment Commission of Shenzhen.We are capable of collecting and treating 32 out of the 49 categories of wasteslisted in the“National Catalog of Hazardous Wastes.” Our treatment capacity is340,000 tons per year.In the last two decades and more, we have collected and treated more than 1.8million tons of various kinds of hazardous wastes, which took up about 80% of thetotal industrial waste produced in the Shenzhen city, and thus achieved goodenvironmental, social and economic benefit.To learn about new technology and equipment of hazardous wastetreatment12CANCELLEDZhuhaiGuangtongAutomobileCo., Ltd珠海广通汽车有限公司http://yintonggZhuhai Guangtong Automobile Co.,Ltd., a wholly-owned subsidiary of YinlongEnergy(the original Yintong Energy), was founded on Aug.30th, 1999. It is anenterprise specialized in producing passenger vehicles (including pure electric vehicle,LNG vehicle).Guangtong Automobile has annual production capability of 10,000 units ofvehicles. It covers a land area of 260,000 square meters, 1768 employees for which157 staff focusing on the research and development of the passenger vehicle, 86 forquality control, 1300 production workers, and 140 for customer services.Guangtong Automobile is focusing on the topics of R&D andtechnology support based on lithium battery material, li-battery, EV andcore components, peak load and frequency regulation of smart powergrids, or any potential cooperation project, product marketing andpromotion activities.13Zhuhai greengold energycontroltechnologyco.,LTD珠海绿金能控科技有限公司http://www.lvjiZhuhai Green Gold Energy Control is an integrated energy control solutionprovider that always delicate to the operation of the online shopping mall for energysaving, the management and control for the city energy, the overall energy savingsolution for the commercial buildings, city lighting, industry energy conservation,intelligent home development as well as others energy saving services.We have very strong R&D team, who acquires key technologies with independentintellectual property rights. With strategies including the integration of solutiondesign, energy efficiency evaluation, energy audit, clean production, technologydevelopment, project investment and financing, project construction, the solid basisof our patented technology of independent intellectual property rights, completesupply chain resource, the satisfactoryservice system, GreenGold has the strong confidence to offer our customer variouscomprehensive energy saving services.1、LED lighting products trade and technical cooperation,2、LED drive power trade and technical cooperation,3、Building energy-saving products New material technology transfer andcooperation,4、Intelligent Home Furnishing product technology transfer andcooperation.14CANCELLEDZHUHAIYINLONGENERGYCO.,LTD珠海银隆新能源有限公司http://www.yinSince its industrialization investment in 2009, the Yinlong Energy strives to build anew energy & closed-loop industrial chain, which focuses on the research,development, production and sales of lithium battery material supply, lithiumbattery. power assembly of electric vehicles, complete vehicles, as well as smartgrid peak and frequency regulation system.We improve the company's comprehensive strength by means of M&A and aim tobuild a leading enterprise with complete new energy industry chain. Through theinvestment and acquisition in 2010 of 53.6% of the stocks of the U.S. NASDAQ-listedAltair Nanotechnologies Inc. (Stock code ALTI), we became its largest shareholder andsuccessfully entered into international capital market. At the same time we masteredthe top production technology of lithium titanate material in the world, and becamethe leader of green energy solutions in the world and the excellent example ofascending the global stage as a China's new energy brand.Yinlong is focusing on the R&D and technology support based on lithiumbattery material, li-battery, EV and core components, peak load andfrequency regulation of smart power grids, or any potential cooperationproject, product marketing and promotion activities.15GuangdongGemsInvestment Co.,Ltd广东金石投资有限公司http://Guangdong Gems Investment Co., Ltd, was established in November 2009 with aseparate legal entity with registered capital of 10 million yuan.Guangdong Gems Investment Co., Ltd is always adhere to the "cooperation,sharing, win-win" business philosophy and "customer first, integrity management,standardized management, coordination and development" business purposes.Through the establishment of standardized operating procedures, improvement ofthe investment risk control system and hold responsible to customer, GuangdongGems Investment form a good management style and is considered as a trustworthycompany within customers and society. Meanwhile, the company is also adhere tothe "professional, professional, elite, discreetness" administration philosophy toattract a team with extensive management experience in finance, management, legal,financial and other sectors of the economy of elite talent team.To seek appropriate investment in the project of helps to expand thecompany's production scale and improve the market share of thecompany's products, improve the comprehensive competitiveness of thecompany, in line with the company long-term plan. Through this activity,Gems investment is expecting to gain insight of foreign cutting-edgetechnology and management experience.16Shenzhen HTIGroup Co.,Ltd深圳市高新投集团有限公司.cnShenzhen HTI Group Co., Ltd. (referred as HTI), established on December 29th1994, is one of the oldest professional companies to provide services in financingguarantee, venture capital and surety bonds in China. HTI has been rated AA in capitalmarket and AAA as a credit guarantee company. The registered capital of HTI is 2.2billion RMB, and the total assets of HTI is 4.1 billion RMB.HTI has greatly supported the growth and development of Huawei, ZTE, Kingdee,Han's Laser, Clou, BYD, O-film and many other high-tech companies through itsguarantee services. Among those companies, 71 companies are listed on capitalmarket home and abroad. By doing so, HTI has earned enormous social and economicbenefits.The surety bonds services of HTI are conducted through its nationwide networkwhich includes seven branches(located in Beijing, Guangzhou, Chengdu, Hangzhou,Xi'an, Hunan and Tianjin) and twenty-five offices. HTI has become the largest suretybonds company in China.To seek cooperation and would like to communicate with local investors.17SHENZHENSMALL&MEDIUMENTERPRISESCREDITFINANCINGGUARANTEEGROUPCO.,LTD深圳市中小企业信用融资担保集团有限公司http://www.szcWith 18 Billion RMB registered capital, Shenzhen Small&Medium EnterprisesCredit Financing Guarantee Group Co. Ltd(CGC) was established on Oct 28.1999,which owns the largest registered capital and serves most customers among all thecredit financing institutions in Shenzhen at present. Besides, our group is the earlistto ranked the highest class AAA of Credit Quality Rating and class AA of CapitalMarket Rating among peers. With branches in districts of Nanshan,Futian,Baoan andLonghua in Shenzhen City, Hangzhou City and Chengdu City, CGC is not merelycovered the company’s service within the whole city of Shenzhen, but also radiates toother provinces. As a catalyst in the small and medium enterprises development, CGCprovides them with integrated financial services, such as loan guarantee, entrustedloans, industrial technology capital guarantee, collective notes guarantee, venturecapital, letter of guarantee, microcredit and pawning.1,Pursuing venture capital projects: Based on the years of experiences inventure capital, CGC is positively seeking the overseas venture capitalprojects for equity investment2,Introducing foreign capital of equity investment: The high-techenterprises of Shenzhen are strongly supported by Shenzhen government.CGC is considering to introduce overseas venture capital to Shenzhen andinvesting into high-tech enterprises to help them go public in future.3,Introducing fund into guarantee industry: As a sunrise industry in ournation, guarantee industry has high development potential. Our group ,one of the leaders in guarantee industry, is attempting to introduceforeign funds into guarantee industry, and in some cases, we could beequity partners.4,Providing Intermediary service in investment and financing fields: As aspecialized agency in investment and financing field ,our group possesses alarge number of superior enterprises, projects ,government resources andnumerous partners.61 enterprises of our clients has gone public since CGCwas set up in 1999. Also, over 200 clients is now in the phase of pre-IPO.Since that our group has a mass of enterprise resources appropriate forequity investment, we could recommend these qualitative projects toforeign investing institutions as an intermediary service.18Zhuhai greengoldinverstmentmanagementco.,LTD珠海绿金投资管理有限公司http://Zhuhai Green Gold Investment Management Limited, registered in 2013,concentrated in investment management, consulting, fiduciary asset managementservices.Green Gold investments has a highly specialized personel finance and researchgroup consisted of 25 people. More than half of our members obtained a masterdegree or above while most of our invertment research staff have been in investmentand financial field for more than 10 years. With our highly specialize team, GreenGold obtained an excellent historical investment performance. Green Gold Investments established a mature and efficient investment decisionmaking, risk control, research support, operational security, marketing and customerservice system for corporate and personal investment management.1、Consulting and management of energy-saving projects;2、Energy-saving project investment and financing cooperation;3、Intelligent home and building energy technology transfer tocooperation;4、Led lighting products and drive power trade and technical cooperation.19CANCELLEDAdministrativeCommittee ofZhongshanNational TorchHi-TechIndustrialDevelopmentZone中山火炬高技术产业开发区管理委员会http://www.zstZhongshan Torch Hi-tech Industrial Development Zone (hereinafter referred to asZhongshan Torch Zone) is a nation-level new hi-tech industrial development zone,which was jointly established by the Ministry of Science and Technology of thePeople’s Republic of China, the People’s Government of Guangdong Province andZhongshan Municipal Government in 1990. Zhongshan Torch Zone, situated on thewest bank of the Pearl River Estuary, is adjacent to Shenzhen and Hong Kong on theopposite sides of the sea. The city of Zhongshan located in the junction of Beijing-Zhuhai Expressway, Seaside Expressway, Zhongshan-Jiangmen Expressway and thePearl River Delta light railway, which is currently under construction. At the sametime, it possesses Zhongshan Port, which ranks top 10 among the ports nationwide interms of container-handling capacity. These super conditions of hinge traffic on thewest bank of the Pearl River Estuary push Zhongshan Torch Zone to the grandcompetition stage of economic circle surrounding Lingdingyang Sea.With a planned area of 13.5 square kilometers, National Health TechnologyIndustrial Base was jointly established by the Ministry of Science and Technology ofthe People’s Republic of China, the People’s Government of Guangdong Province andZhongshan Municipal Government in 1994.As the first domestic health technology industrial base established subject tointernational criteria, the Base is mainly engaged in research and development, clinictrial, production and sale of traditional Chinese and western medicines, bio-engineering, medical instruments, healthy foods and cosmetics. Furthermore, it offersa series of professional services including land purchase, factory building lease, designand construction, company registration, medicine proof-testing and application, legalconsultation, market exploitation, human resource recruitment, tax programming,financing and GMP authentication. Up to present, over 100 enterprises have settleddown in the Base, covering production, research, management and sale that haveformed a perfect industrial chain and industry system of pharmaceuticals, which haveattracted a lot of talents, capital and technology related to health industry. The basehas also exploited parks like South China City of Modern Traditional Chinese MedicineWe are seeking entrepreneurs with investment intention, Particularly inhealth technology industry.20HuaguandaEngineeringCo.Ltd华冠达工程有限公司http://www.ch/Founded in 1998, Huaguanda Engineering Co.Ltd possessed the registered capitalof RMB 102 Million. At present, the company has obtained the first class qualificationof construction for overall contracting, takes special class “AAA” reputation aroundChina and passed the ISO9001 Quality System Certification.The company employs over 600 employees, many of them held middle and seniortechnical title.We’ve set up an advanced system of enterprise management.The company has achieved great success in business for “fulfill keep the terms ofcontracts and his words”.The company will try its best to provide the high quality project and service for allcircles of the world with faithful service and hard work.Energy conservation and environmental protection technicalcommunication about the construction industry.21ShenzhenInvestmentHoldingsIndustrial ParkDevelopmentAnd OperationCompany Ltd.深圳市投控产业园区开发运营有限公司http://www.sihShenzhen Investment Holdings Industrial Park Development and OperationCompany is a wholly owned subsidiary of Shenzhen Investment Holdings Company,which is a municipal state-owned assets management company.The main responsibilities of the company are the administration of industrial parkdevelopment & construction projects, sales, leasing and operation management. Thecore business included industrial estate construction, development, operationmanagement and technologial financial services.Shenzhen Investment Holdings Industrial Park Development andOperation Company is to carry out the construction and operationmanagement of six Shenzhen municipal investment and financing projects.Currentlly, Shenzhen Software Industry Base and Shenzhen Bay EcologicalPark are inviting high-tech, strategic cooperation projects, new energy andother related enterprises and research institutions.22ShenzhenKenuo BridgeTechnology Co.Ltd.深圳科诺桥科技有限公司http//www.szkShenzhen Kenuo Bridge Technology Co. Ltd., is located in the electronics industrydeveloped Shenzhen City Guangdong province. The company possess 4000 m2purification workshop, a registered capital of RMB 20 Million,150 people work in thefirst stage of the project with 30 of them specialized in engineering technique,advanced production and testing equipment, perfect production and qualitymanagement system. Traditionally adhering to the "Shenzhen speed" spirit, wepromote pragmatic and efficient production.Through excellent talents and advancedtechnology, the company is com mitted to the research, development, productionand sales of electromagnetic shielding materials, services in flexible circuit boardindustry. The company has the ability to produce 100000 m2 of electromagneticshielding film per month. In addition, we provide conductive adhesive film, pure filmand other products to the customer.Under the company principle of "professional production with high quality ofcustomer service" we share the benifit of advanced production technique with ourcustomers.Seek ambitious cooperator that can help Kenuo Bridge todevelop into a world class circuit board material manufacturewithin 5 years.Develop conductive adhersive film, pure filmand non adhesive FPC when the supply of electromagneticshielding film is stable.23SHENZHENTONGCHANGROUPCO.,LTD深圳市通产集团有限公司http://www.sztShenzhen Tongchan Group Co.,Ltd. , founded in 2000 with a total assets near 4billion RMB, has more than 6 thousand employees. It owns Shenzhen Beauty StarCo.,Ltd, Zhaoqing Tongchan Glass Technology Co.,Ltd, Sichuan Tongchan HuajingGlass Co.,Ltd, Shenzhen Huajing Glass Bottle Co.Ltd, Shenzhen 863 MaterialTechnology R&D Center etc., as the wholly owned or holding enterprises. It also holdsthe share of several high-quality companies and some listed corporations. Thecompany’s main business are high-end cosmetic plastic packaging and glasspackaging. The major customers of plastic packaging are P&G, Shiseido, Unilever,Avon, L’Oreal, Arrow while the main customer of glass packaging are Carlsberg,Heineken, Tsingtao, Pearl River, Haitian and the other well-known beermanufacturers. Some products are also exported to South-East Asia.To invite buyer from AB InBev Group's headquater in Belgium toparticipate the meeting, the main topic is as following:1. To develop long-term cooperation relationship between Tong chanGroup and AB InBev (China) market;2. To learn more about the glass bottle's development trend;3. To discuss about AB InBev's global demand on the glass bottle。

R2Auto-Tuning Fiber-Optic SensorE3X-NHigh Performance DC Amplifier with Pushbutton Sensitivity Adjustment H Maintenance is made easier withpushbutton sensitivity adjustmentH Sensing distance is up to 100%longerthan standard amplifiersH Choose either single channel or fourchannel modelsH Compact housing:32mm and 10mmhousing widthsH Four amplifiers in a single housingsaves space and wiringH Four fiber-optic cables can be mounteddirectly next to each other without mutual interferenceOrdering InformationJ AMPLIFIERSTypeSingle channel Four channel Off-delay timer None Provided Provided Remote teach input None Provided Provided Part numberNPN outputE3X-NT11E3X-NT21E3X-NM11PNP outputE3X-NT41E3X-NT51E3X-NM41J FIBER-OPTIC CABLESPlease see the E32Fiber-Optic Cables section.J ACCESSORIESDescriptionPart number Replacement protective cover for E3X-NT jj E39-G8Replacement protective cover for E3X-NM jjE39-G9E3X-NE3X-N3SpecificationsDescription General-purpose Multi-function Four-channel Part number NPN E3X-NT11E3X-NT21E3X-NM11PNPE3X-NT41E3X-NT51E3X-NM41Supply voltage 12to 24VDC ±10%,10%ripple max.Current consumption 50mA max.150mA max.Required fiber-optic cables Please see the E32Fiber-Optic Cables section.Light source Pulse modulated red LED (680nm)Operation mode Light-ON,Dark-ON (switch selectable)SensitivityPushbutton setting (see operation section)Mutual interference protection Provided Remote teach option —Provided Control outputDC solid stateType NPN open collectorPNP open collector p Max.load 100mA,30VDC max.Max.ON-state voltage drop1VDC max.at 100mAResponse time 500µs max.at rated detection distance Timing functions —OFF:delay,40ms,selectable ON or OFFCircuit protectionOutput short circuit protection,DC power reverse polarity protection Teaching confirmation function Indicators (red/green LED)and buzzerIndicators Operation (orange LED)and output stability (green LED)Materials Case PBT plastic Cover PolycarbonateMounting DIN-rail track,or on flat surface through holes in bracket (provided)Pre-leaded 3conductor cable,4conductor cable,7conductor cable,Connections Pre leaded 2m (6.5ft)2m (6.5ft)2m (6.5ft)Weight100g (3.5oz.)with 2m cableEnclosure ratings UL —gNEMA —IECIP 50(with cover on)Approvals UL —ppCSA —Ambient temperature Operating:--25°C to 55°C (--13°F to 131°F)with no ice buildup pStorage:--40°C to 70°C (--40°F to 158°F)E3X-NE3X-N4J OUTPUT CIRCUIT DIAGRAMSE3X-NT11E3X-NT21E3X-NM1112to 24VDC12to 24VDC12to 24VDCinputControl outputwire color Channel no.1234Black White Grey OrangeE3X-NT41E3X-NT5112to 24VDC12to 24VDCinputE3X-NE3X-N5NomenclatureE3X-NT11(NPN)E3X-NT21(NPN)E3X-NT41(PNP)E3X-NT51(PNP)E3X-NM11(NPN)E3X-NM41(PNP)Operation indicator Stability indicator (green)Teaching indicator (red/green)Teaching Mode selectorTimer switch (see Operation mode Four operation indicators (orange)Four stabilityindicators Channel selectorTeachingindicatorOperationselectorsTeaching Mode selectorNote:The E3X-NT11or E3X-NT41donot have a timer function.OperationJ CHOOSING THE RIGHT TEACHING METHODRefer to the following table to select the most suitable sensitivity setting method.Teaching method Maximum sensitivity setting No-object teaching With/Without-object teaching Typical applicationDetection of the existence of objects that interrupt light perfectlyDetection of objects with no background objectsIf teaching is impossible by stopping the movement of sensing objectsTo detect bright or dark objectsby teaching only with backgroundobjectsDetection of a slight difference in reflectionColor discriminationBackground objects withunstable reflectionDetection of object surface irregularitiesElimination of background object influenceNote:1.None of the four channels has any output when the E3X-NM (four channels)is in teaching mode (i.e.,all the four channels willbe in teaching mode).2.If the set distance is very short (i.e.,0to 12mm for the E32-TC200and 0to 4mm for the E32-DC200),no-object teaching isnot possible due to excessive light,in which case,perform with/without-object teaching.3.In principle,use the E3X-NM (four channels)for the close connection of a maximum of four Fiber Units.When closely connect-ing two to three Fiber Units to more than one E3X-NT (one channel),perform with/without-object teaching,in which case teach-ing must be performed on a single E3X-NT at a time.Therefore,turn on only the E3X-NT on which teaching is performed.If all the E3X-NTs are turned on,interrupt the emitters of the Fiber Units on which teaching is not performed.E3X-NE3X-N6J MAXIMUM SENSITIVITY SETTINGNote:The sensitivity of the E3X-NT and E3X-NM are set to maximum before shipping.When resetting the sensitivity of the E3X-NT orE3X-NM to maximum after no-object teaching or with/without-object teaching,follow the steps described below.Procedure OperationE3X-NTE3X-NM1Locate the sensor head within the rated sensing range with the E3X-N j .2Set the mode selector to TEACH.TEACHRUNTEACH TIMER SET RUN3The flashing function of the E3X-N j will be activated.Therefore,adjust the optical axes so that the tip of the emitting fiber will be lit.If the optical axes are divergent,the tip of the emitting fiber will flash and the built-in buzzer of the E3X-N j will beep.---4Press the teaching button for three seconds minimum with or without a sensing object.In the case of the E3X-NM,select a channel with the channel selector,at which time the stability indicator for the selected channel will flash.The teaching indicator (red)turns green.The built-in buzzer beeps once when the color of the teaching indicator is red.The built-in buzzer beeps continuously when the color of the teaching indicator is green.Note:The built-in buzzer will stop beeping when the teaching buttonis no longer being pressed.TEACHTEACH5Set the mode selector to RUN to complete the sensitivity setting.The teaching indicator is OFF.Note:When the sensitivity is set to maximum,the sensitivity will beautomatically adjusted regardless of the set distances of the fibers or light.TEACHRUNTEACH TIMER SET RUN6Select the logical output required with the operation mode selector.L D ON ON1234L ON D ONCHE3X-NE3X-N7J WITH/WITHOUT-OBJECT TEACHINGRUNteaching buzzer Light is terrupted.TEACHTEACHobject is is have Through-beam Modelis received.TEACHTEACHTEACHRUN TEACH TIMER SET RUN L D ON ON1234L ON D ONCH Note:1.Even if the E3X-N j is turned off,the E3X-N j will retain the sensitivity set at the time of teaching.2.Channels (E3X-NM)are selected in the following order.When all the channels are selected,it is possible to set the sensitivity of the E3X-NM on all channels.1CH2CH 3CH4CHALLE3X-NE3X-N8J NO-OBJECT TEACHINGProcedure OperationE3X-NTE3X-NM1Locate the sensor head within the rated sensing range with the E3X-N j .2Set the mode selector to TEACH.TEACHRUNTEACH TIMER SET RUN3The flashing function of the E3X-N j will be activated.Therefore,adjust the optical axes so that the tip of the emitting fiber will be lit.If the optical axes are divergent,the tip of the emitting fiber will flash and the built-in buzzer of the E3X-N j will beep.---4Press the teaching button for 0.5to 2.5seconds without a sensing object.In the case the E3X-NM,select a channel with the channel selector and press the teaching button,at which time the stability indicator for the selected channel will flash.The teaching indicator (red)is lit.The built-in buzzer beeps once.TEACHTEACH5Set the mode selector to RUN.No-object teaching will be set when the first sensing object passes through the sensing area.The teaching indicator (red)turns green (automatically turned off in one second).TEACHRUN TEACH TIMER SET RUN6Select the logical output required with the operation mode selector.L D ON ON1234L ON D ONCH Note:1.To detect dark objects in front of bright backgrounds,set the operation mode selector to D.ON.2.If the set distance is very short (i.e.,0to 12mm for the E32-TC200and 0to 4mm for the E32-DC200),no-object teaching isnot possible due to excessive light,in which case,perform with/without-object teaching.3.If the teaching button is pressed for more than three seconds,the sensitivity of the E3X-N j will be set to maximum,at whichtime the green indicator will be lit.4.The E3X-N j will be ready to detect objects in approximately one second after the mode selector is set to RUN.J SENSITIVITY ADJUSTMENTCombination of the E3X-NT/E3X-NM and Fine Through-beam Fiber Units (E32-T22S/T24S/T84S)No-object TeachingPress the teaching button once with no object in the sensing area.ObjectNote:If detection is not stable after no-object teaching,performwith/without-object teaching.With/Without-object TeachingPress the teaching button once with no object in the sensing area.。

Wireless Sensor Network, 2012, 4, 162-166doi:10.4236/wsn.2012.46023 Published Online June 2012 (/journal/wsn)MNMU-RA: Most Nearest Most Used Routing Algorithm for Greening the Wireless Sensor NetworksHafiz Bilal Khalil, Syed Jawad Hussain ZaidiSchool of Electrical Engineering & Computer Sciences, National University of Sciences and Technology, Islamabad, PakistanEmail: {10mseetkhalil, 10mseejzaidi}@.pkReceived February 22, 2012; revised March 22, 2012; accepted April 10, 2012ABSTRACTWireless sensors are widely deployed in military and other organizations that significantly depend upon the sensed in-formation in any emergency situation. One of the main designs issues of the wireless sensor network (WSN) is the con-servation of energy which is directly proportional to the life of the networks. We propose most nearest most used rout-ing algorithm (MNMU-RA) for ad-hoc WSNs which vitally plays an important role in energy conservation. We find the best location of MNMU node for energy harvesting by apply our algorithm. Our method involves the least number of nodes in transmission of data and set large number of nodes to sleep in idle mode. Based on simulation result we shows the significant improvement in energy saving and enhance the life of the network.Keywords: Energy Efficiency; Wireless Sensor Networks; Routing1. IntroductionThe growth in wireless sensor networks and its applica- tions dramatically increased in last decade. Wireless sen- sor nodes are widely used in military surveillance, intel- ligence and targeting in war operations. Energy available at each sensor for sensing and communications is limited because of the cost constraints and smaller size, which affects the sensor application and network lifetime. The purpose of green networking is to overcome the carbon foot print, reduce the energy consumption and energy losses. Energy efficiency is an important issue to enhance the life time of the network. To achieve the green net- working every component of the network is integrated with energy efficient protocols, e.g., energy-aware rout- ing on network layer, energy-saving mode on MAC layer, etc. One of the most important components of the sensor node is the power source. In sensor networks generally there are three modes of power consumption: sensing, data processing, and communication. Compared to sensing and data processing, much more energy is required for data communication in a typical sensor node [1]. These are also categorized as sleep (idle) and wakeup (trans-mission) mode.In ad-hoc WSNs (Wireless Sensor Networks) always the nodes are cooperative, they sense and transmit their own data and also act as router to route the sensed infor- mation of other nodes towards the data center or gateway node which is connected to the internet. Most of the nodes consumed their power resource while transmitting the data of neighboring nodes. The scope of this paper is to minimize the power consumption in transmitting or routing process and set large number of nodes into sleep mode. The remaining sections of this paper organized as follows. Section 2 explains related work and current en-ergy efficient techniques for sensor networks. Section 3 introduces some problems and research issues in current work. Section 4 describes overview of network model, our proposed algorithm and proposed solution respec-tively. In Section 5 experiment, Results and comparisons are given.2. Related WorkEnergy efficiency is already achieved by many appro- aches. These approaches include energy aware protocol development and hardware optimizations, such as sleep- ing schedules to keep electronics inactive most of the time, dynamic optimization of voltage, and clock rate. In[2] Smart Dust motes are designed that are not more thana few cubic millimeters. They can float in the air, keep sensing and transmitting for hours or days. In [3] authors described the µAMPS wireless sensor node, it is hard- ware based solution in which they simultaneously con- sider the features of the microprocessors and transceivers to reduce the power consumption of the each wireless sensor node in network. Routing algorithms also play an important role to reduce the energy consumption during the routing of data. A lot of work is done in MAC layer and Mac protocols;MAC protocol for wireless sensorH. B. KHALIL, S. J. H. ZAIDI163networks is not like the traditional wireless MACs such as IEEE 802.11. One of the most important goals is en-ergy conservation, fairness and latency is less important [4].SMAC/AL (Sensor MAC with Adaptive Listening) is a famous MAC protocol for WSNs proposed by Ye et al. [5,6]. Main purpose of SMAC/AL is to reduce energy consumption. But in SMAC/AL without considering the distance among the nodes, all nodes unnecessarily con- sume the energy by transmitting information with con- stant power level. An energy efficient MAC protocol with adaptive transmit power scheme named ATPM (Adap- tive Transmit Power MAC) is proposed in [7]. By meas- uring the received power ATPM can calculate the dis- tance between the sender and the receiver, and then adap- tively choose the suitable transmit power level according to the propagation model and distance. The ATMP can not only conserve the energy source, but also decrease the collision probability. A Novel Clustering Algorithm for Energy Efficiency in Wireless Sensor Networks (AN-CAEE) has been proposed [8]. It minimizes energy utili-zation during data transmission and energy consumptions are distributed uniformly among all nodes. Each cluster contains cluster head, each node send its data to cluster head with single hop transmission. And cluster transmits the combined data to the base station with multi hope transmission. This approach reduces energy consumption of nodes within the cluster.3. Problem StatementSensor nodes which are one hope away or closest to the gateway node always consume their power more quickly than others because they have to transmit the data of other nodes in addition to transmission of their own sensed information. In [9] a solution was proposed for such type of scenario by implementing the multiple base stations and periodically changing their positions. But the prob- lem is that if every time the most far away sensor trans- mits its data then major part of overall network energy will be consumed. Another solution for prolong the sen- sor network lifetime is to divide sensors nodes into dis- joint sets, such that all the targets completely covered by every set [3]. Authors consider that within an active sen- sor’s operational range a target is covered. These disjoint sets are activated in round robin fashion, such that at a time only one set is active. Sensors are into the active state in an active set and all other sensors are in a low- energy sleep state. According to this method almost half of the sensor remains active and remaining half goes to sleep mode which reduce energy down to 50%. To make it more efficient and conserve the larger amount of en- ergy we proposed an algorithm named as MNMU-RA (Most nearest most used routing algorithm). That algo- rithm finds the efficient placement of active sensor nodes and set other nodes into sleep mode. An issue is also re- solved by our algorithm, reducing the number of multiple base stations by finding out the best location for the base station without changing its location periodically.4. Synopsis of Our Network ModelIn this paper we deal with the issue of energy efficiency in wireless sensor networks for surveillance of a set of targets with known locality. Scenario of the network is chosen for armed forces purposes like surveillance of the boarder, battle fields and no go areas to acquire the in- formation about enemies and their locations without tak- ing the risk for human personal. We consider that a large number of sensors are distributed randomly in close prox- imity for monitoring and send the monitored information to a gateway node. All nodes are static and makes ad-hoc wireless sensor network. Every sensor nodes must moni-tor the area all the time in its operational range and each sensor has fixed transmission range. In network model we assume that each sensor has unique pre configured Id and Global/proactive routing algorithms are used. Main advantage of proactive algorithm is not route latency but drawback is the high maintenance overhead when many of the routes are never used.Proactive routing is appro-priate for networks with: Small size, low mobility and high communication rates. We proposed an algorithm called as most nearest most used routing algorithm for this purpose. By using MNMU-RA we can find the per-fect location of node for energy harvesting which also reduce the overall energy consumption and cost.4.1. Most Nearest Most Used Routing Algorithm Run shortest path routing algorithm or link state routing to find the shortest path for each node in the wireless sensor network. Calculate all the possible shortest paths for each node. Then find the MNMU node (Figure 1).∙ A node which is most nearest to the gateway node.∙Select a node which is used in maximum number of shortest paths.Figure 1. Location of selected MNMU node.H. B. KHALIL, S. J. H. ZAIDI 164In above network model we assumed that sensed in- formation is equally probable for all the nodes. Then we calculate the shortest path for the nodes A, B and C. Then we find out the nodes which are most nearest to the gate way node. In above network model there are only two nodes X and Y which are closer to the gateway node. Then for selection we give the preference to the node which is most used in shortest paths. In above model Y is node which is most used in all shortest paths. If nodes A, B and C transmit their data the entire time node Y will be included in their path. Then every node keeps its routes information towards the node Y for future communica- tions. Flow chart of our algorithm is given in Figure 2. 4.2. Proposed SolutionWe used our algorithm to find most nearest most used node in a network, that node should be active all the time while other sensors remain in sleep mode and keep sens- ing. As we use proactive routing so each sensor knows its path towards the MNMU node. If a node has to send its information before sending it will wake up the nodes along his route. When MNMU nodes receive the infor- mation it will forward the data to the gateway and sets all the nodes into sleep mode. The critical issue in this solu- tion is that if a node (MNMU node) remains active all the time then its energy source will be empty soon. We re- solve this issue by using the energy harvesting concept at MNMU node [1]. We can also use secondary batteryFigure 2. MNMU routing algorithm flow chart. which is rechargeable and coupled with photovoltaic cell[10]. If all the nodes can generate energy from light, vi-bration, heat etc [11,12] it will increase the system cost.We don’t need to replace all the nodes with secondary sources. By replacing only one node (MNMU node) re-solves the issue and slightly increases the cost of theoverall system. But effectively prolong the life time ofsensor network. A solution given by Gandham et al. [9]can be more energy efficient if we implement our pro-posed algorithm with every new location of mobile basestation. Split the network in equal parts and periodicallychange the position of base station in each part. Basestation can be easily implemented at the place of MNMUnode in each part of the network instead of replacing itoutside the network. MNMU node will reduce the multihop and number of transmission which directly reducethe energy consumption.5. ExperimentWe done the experiment by implementing our proposedalgorithm in a network and calculate the amount of en-ergy utilization using MATLAB. Then implement theconcept of disjoint set and analyze the values at same network. For simulation 20 nodes containing one gate-way node are distributed randomly in 30 meter squarearea. We consider the features of MICA2 mote platform.It is third generation mote specifically built for WSNs [4].MICA2 have selectable transmission power range whichoffers adjustable communication ranges, selected trans-mission range for each node is 10 meters. The packetlength is fixed at 200 bits. MICA2 usually operated with3 V battery and other features mentioned in Table 1.We divided our analysis in three parts; first we calcu-late the power consumption using disjoint sets methods[3], then we apply our algorithm and calculate & com-pare power consumption. Same network and topologytaken in which each node remains active all the time andno energy saving protocol and technique is implemented.Energy calculated during the 20 rounds, all nodes areactive in first five rounds in which they sense and trans-mit the data. After ten rounds there is no activity andnodes go to sleeping mode according to implemented Table 1. Features of MICA2 motes platform [12,13].Operation/Features UnitListening 8mA Receiving 10mA Transmission 17mA Sleep 19µA Radio Frequency 900 MHzCPU 8 bit Atmel at 8 MHzBandwidth 40KbpsH. B. KHALIL, S. J. H. ZAIDI165methodology. Calculated results are given in Figures3 and 4.Simulation ResultsFigure 3 shows the result comparison of energy con- sumption in different modes; sensing, Transmission and sleeping of network. In Figure 3(a) set of all the active nodes shown by blue line are transmitting the data with- out applying any energy saving protocol. During the transmission if all nodes are active they will keep trans- mitting the information to each other and maximum amount of energy is consumed. In disjoint system only active set take part in transmission and inactive nodesFigure 3. Power consumptions in different modes. (a) Trans- mission mode; (b) Power consume by sleeping nodes; (c)Power consume by active nodes in sleep mode. Figure 4. Result and comparison of energy consumption in different modes.remain inactive during the transmission of active set. Our proposed algorithm gives lowest amount of energy con- sumption because only the MNMU node and less number of nodes take part in transmission. Energy consumed by inactive nodes in sleeping modes is shown in Figure 3(b). Energy consumption of sleeping nodes is in µwatts. Ac- cording to our algorithm 19 nodes set to sleep mode and only one MNMU node is active. While Figure 3(c) shows the separately calculated energy consumption by active nodes when there is no activity and network is in idle mode. Similarly in sleeping mode only MNMU node remains active and rest of the network sets to sleep mode. Figure 4 shows the result of energy consumption of entire network in different rounds. In first 5 rounds we assume that there is no sensed information to send; all the nodes are active in listening mode and keep sensing. In 5 to 10 rounds nodes are transmitting their sensed in- formation to the gateway. After round 10 there is no ac- tivity and nodes set to sleep mode in sleep mode only energy consumed by active nodes are calculated and en- ergy consumed by sleeping nodes which is in µwatts is neglected. Our algorithm gives the minimum energy con- sumption during the transmission in which fewer num- bers of nodes take part in routing and also in sleep mode by keeping only MNMU node active.6. ConclusionWe presented the most nearest most used routing algo- rithm to reduce the energy utilization in wireless sensor networks. Using this algorithm we find the best location of energy harvested node in a network. Our algorithm involves least number of nodes during transmission and keeps one node active in sleep mode. That significantly reduces the energy consumption during the transmissionH. B. KHALIL, S. J. H. ZAIDI 166and sleep mode when there is no activity. An open re- search issue is the heterogeneity of energy resources of the nodes that must be resolved after practical imple- mentation in any network. In our solution there is uneven energy consumption due to the topology of the network and nature of data flow. But that uneven energy con- sumption is helpful to reduce the energy consumption of entire network7. Future DirectionDesired goal in wireless networks is energy efficiency to maximize the network life. Our algorithm can be used to find the location of cluster header quickly in novel clus- tering algorithm for energy efficiency in wireless sensor networks [8]. Further we can implement coding tech- niques to reduce the number of transmissions at MNMU node. Energy consumes per bit or per packet transmis- sion can be reduce. Number of packets can be transmit- ted as a single packet by applying x-or Operations which reduces the energy but may cause of slighter delay. To apply this technique sensor nodes must be smarter and have ability to do this quickly.REFERENCES[1]I. F. Akyildiz, T. Melodia and K. Chowdhury, “A Surveyon Wireless Multimedia Sensor Networks,” ComputerNetworks, Vol. 51, No. 4, 2007, pp. 921-960.doi:10.1016/net.2006.10.002[2]J. M. Kahn, R. H. Katz and K. S. J. Pister, “EmergingChallenges: Mobile Networking for Smart Dust,” Inter-national Journal of Communication Networks, Vol. 2, No.3, 2000, pp. 188-196.[3]M. Cardei and D. Z. Du, “Improving Wireless SensorNetwork Lifetime through Power Aware Organization,”Wireless Networks, Vol. 11, No. 3, 2005, pp. 333-340.doi:10.1007/s11276-005-6615-6[4]Q. Hu and Z. Z. Tang, “An Adaptive Transmit PowerScheme for Wireless Sensor Networks,” 3rd IEEE Inter-national Conference on Ubi-Media Computing, Jinhua, 5-7 July 2010, pp. 12-16.[5]W. Ye, J. Heidemann and D. Estrin, “An Energy-EfficientMAC Protocol for Wireless Sensor Networks,” Proceed- ings of the IEEE INFOCOM, New York, 23-27 June 2002, pp. 1567-1576.[6]W. Ye, J. Heidemann and D. Estrin, “Medium AccessControl with Coordinated Adaptive Sleeping for Wireless Sensor Networks,” IEEE/ACM Transactions on Network- ing, Vol. 12, No. 3, 2004, pp. 493-506.doi:10.1109/TNET.2004.828953[7]Q. Hu and Z. Tang, “ATPM: An Energy Efficient MACProtocol with Adaptive Transmit Power Scheme for Wire- less Sensor Networks,” Journal of Multimedia, Vol. 6, No.2, 2011, pp. 122-128. doi:10.4304/jmm.6.2.122-128[8] A. P. Abidoye and N. A. Azeez, “ANCAEE: A Novel Clus-tering Algorithm for Energy Efficiency in Wireless Sen- sor Networks,” Journal of Wireless Sensor Networks, Vol.3, No. 9, 2011, pp. 307-312. doi:10.4236/wsn.2011.39032 [9]S. R. Gandham, M. Dawande, R. Prakash and S. Venkate-san, “Energy Efficient Schemes for Wireless Sensor Net- works with Multiple Mobile Base Stations,” Global Tele- communications Conference, San Francisco, 1-5 Decem- ber 2003, pp. 377-381.[10]M. A. M. Vieira, C. N. Coelho, D. C. Silva and J. M. Mata,“Survey on Wireless Sensor Network Devices,” Proceed- ings of IEEE International Conference on Emerging Tec- hnologies and Factory Automation (ETFA’03), Lisbon, 16-19 September 2003, pp. 537-544.[11]J. Paradiso and T. Starner, “Energy Scavenging for Mo-bile and Wireless Electronics,” Pervasive Computing, Vol.4, No. 1, 2005, pp. 18-27. doi:10.1109/MPRV.2005.9 [12]V. Gungor and G. Hancke, “Industrial Wireless SensorNetworks: Challenges, Design Principles, and Technical Approaches,” IEEE Transactions on Industrial Electron- ics, Vol. 56, No. 10, 2009, pp. 4258-4265.doi:10.1109/TIE.2009.2015754[13]CrossBow, Mica2 Data Sheet./Products/Product_pdf_files/MICA%20data%20sheet.pdf。


Historic Achievement RecognizedSHIPPINGPORT ATOMIC POWER STATIONA NationalHistoric Mechanical Engineering LandmarkOn December 8, 1953 President Dwight D. Eisenhower went before the United National General Assembly to propose an “Atoms For Peace” plan. One of the cornerstones of the plans for the peaceful uses of atomic energy was the building of a nuclear power plant for the commercial generation of electricity to be used as part of a utility system. Thus the congressional Joint Committee on Atomic Energy and the Atomic Energy Commission developed plans for an atomic power station.A large-scale light water reactor for a proposed aircraft carrier was then being designed at Westinghouse Electric Corporation’s Bettis Atomic Power Laboratory under the technical direction and in cooperation with the Division of Naval Reactors of the Atomic Energy Commission. However, the Federal Government had cancelled the plans for the aircraft carrier. It was decided this design effort could be redirected into a civilian reactor for electric power production. The AEC notified the electric utility industry that they were accepting bids from utilities with the proposal that the utility participate in building and operating a full-scale atomic power station as part of its generating system.The AEC gave responsibility for supervision of the building and operation of the reactor plant portion of the electric generating station to its experts on light water reactors: The Division of Naval Reactors. With a bid that included supplying the land, the turbine generator and $5 million toward the research and development of the light water reactor, Duquesne Light Company was chosen as the utility for the project. The land was located in the small village of Shippingport, Pennsylvania on the Ohio River about 25 miles from Pittsburgh.The principal responsibility for carrying the Shippingport project to a successful conclusion was given to then Rear Admiral H.G. Rickover, Director of the AEC Division of Naval Reactors.SHIPPINGPORT ATOMICPOWER STATIONNational Historic MechanicalEngineering LandmarkPresentation CeremonyMay 20, 1980PROGRAMWelcoming Remarks JOHN T. POPEVice President, Region VAmerican Society of Mechanical EngineersIntroduction of Guests THOMAS R. CURRANChairman, Pittsburgh SectionAmerican Society of Mechanical EngineersASME Landmark Program PROFESSOR J. J. ErmencChairman, National History and Heritage CommitteeAmerican Society of Mechanical EngineersHistory of Shippingport STANLEY G. SCHAFFERAtomic Power Station President, Duquesne Light CompanyPresentation of Plaque DONALD N. ZWIEPPresident, American Society of Mechanical EngineersAcceptance STANLEY G. SCHAFFERPresident, Duquesne Light CompanyClosing Remarks THOMAS R. CURRANChairman, Pittsburgh SectionAmerican Society of Mechanical Engineers Luncheon Remarks J. J. TAYLORVice President and General Manager, Water ReactorsDivisions, Westinghouse Electric CorporationInformal viewing of exhibits in the Shippingport Visitors’ Centerand a luncheon at the Willows Motel Restaurant, Industry,Pennsylvania, follow the presentation ceremonies.ATOMS FOR PEACE - President Dwight D. Eisenhower, participated via electronic communi-cations in both the groundbreaking and dedication of the Shippingport Atomic Power Station. For the groundbreaking, he passed a neutron wand over neutron counter which flashed an electronic signal from Denver, Colorado to Shippingport, activating large highlift which turned the first scoop of ground.The Shippingport AtomicPower Station project receivedworld wide attention from thevery beginning. The groundbreaking ceremony on September6, 1954 (Labor Day) was attendedby about 1,400 people, includingdignitaries representing nationsfrom around the world. PresidentDwight D. Eisenhower, the fathero f t h e“A t o m s F o r P e a c e”program, participated in theceremony via an electric hookupfrom a summer White House inDenver, Colorado.In his address, PresidentEisenhower said: “. . . For today atShippingport, Pennsylvania, webegin building our first atomicpower plant of commercial size—a plant expected to produceelectricity for 100,000 people. Inthus advancing toward theeconomic production of elec-tricity by atomic power, mankindcomes closer to fulfillment of theancient dream of a new and betterearth.“. . .through knowledge weare sure to gain from this newplant we begin today, I am con-fident that the atom will not bedevoted exclusively to the destruc-tion of man, but will be his mightyservant and tireless benefactor.“It is then with profoundhope and confidence—and withprayer for future ages of mankind— that I now, by this act beginconstruction of America’s firstcommercial-size atomic powerplant.”The President’s address was broadcast to the construction site via television and shown to the guests on 20 strategically located television sets. As he finished, President Eisenhower passed neutron wand over a neutron counter which flashed an electronic signal 1,200 miles to Shippingport, activating a large highlift which moved forward and scooped the first dirt in the ground breaking ceremony. Sensing the historic significance of the moment, the crowd rose to its feet and applauded.Speaking for Duquesne Light Company at the ceremony, Philip A. Fleger, Chairman of the Board, said, when describing the Shippingport project, “Here private industry invests its capital, its experience, and its productive skill in cooperation with the government to bring the benefit of a great, new natural resource to the American people.INDUSTRY AND GOVERNMENT COOPERATION was evident through-out the project. Participants in meetings might have included, from left, Walter J. Lyman, Duquesne’s Vice President, Operations, Admiral H.G. Rickover, Director of the AEC Division of Naval Reactors, standing, and Carroll T. Sinclair, Duquesne’s Vice President, Engineering and Construction.“Here the Atomic Energy Commission, the Westinghouse Electric Corporation, and the Duquesne Light Company embark upon an adventure which should do much to advance the use of atomic energy—through electricity—and thereby promote the well-being of all the people.“The Duquesne Light Company accepts its share of the responsibility for this epochal undertaking with the greatest humility but with equally great determination.”Millions of people across thenation were able to see and listento the proceedings of the ground-breaking via radio and televisionas Chairman Lewis L. Strauss of theU.S. Atomic Energy Commissionaddressed the crowd. “Only alittle more than a year ago,” Mr.Strauss said, “it was believed thatp r o d u c t i o n o f c o m m e r c i a lamounts of electric energy fromnuclear power would have to bedemonstrated by the government. . . and by the government alone. . . before private industry wouldor could afford to take part in it.“But so rapid have been thestrides in scientific and engineer-ing achievement that here, todaythe government . . . this is to say,the people . . . begin such anenterprise which is more funda-mentally a pioneer adventurethan the first railroad to penetratethe West or the first airline to spanthe continent.”In an effort to put the days proceedings in perspective, the Honorable W. Sterling Cole, Chairman, Joint Congressional Committee on Atomic Energy said, “Now we are at the end of the beginning. Now we translate our hope and dreams of using the atom for the pursuits of peace into the concrete and steel of a plant producing large amounts of electricity. Along the shores of this river a new marvel of science and engineering shall soon testify that the atom has been bent to the way of peace.”Westinghouse Electric Corporation President, Gwilym A. Price, termed the building of the atomic power plant as “Mankind’s progress towards a bright horizon.”Mr. Price said, “I am, I believe, excusably proud that only forty miles from here the Westinghouse Atomic Power Division is building the reactor—or atomic furnace—which will energize America’s first full-scale atomic power plant. And if we finish well what we begin here today, the atom will bring into existence the truly ‘Golden Age of Electricity’.“The success of this plant eventually will mean great benefits for people everywhere. It forecasts readily available electric power in almost any corner of the earth and the great productivity which alone can bring material well-being to the ‘have not’ peoples of the world. It may indeed be the first dawning of the new age of plenty which the world has long awaited.”GROUNDBREAKlNG CEREMONlES featured a large highlift which when electronically activated by President Eisenhower over 1,200 miles away, turned the first scoop of ground for the Shippingport project. When this occurred, the crowd at the ceremonies, sensing the historic significance of the moment, rose to its feet and applauded. Pictures of this occasion were featured worldwide by newspapers, television and movies.After the ground breaking,the serious business of construc-ting the new plant began inearnest. Throughout constructionthe Shippingport Atomic PowerStation would continue to receiveinternational attention as a majoradvancement in technology.During construction, AdmiralRickover would frequently visitthe construction site to checkprogress. He would confer withthe project managers: Joseph C.Rengel of Westinghouse, JohnGray of Duquesne Light, JohnSimpson, head of the Westing-house Bettis Atomic Power Labora-tory, and Melvin Oldham whowould be the Duquesne Lights u p e r i n t e n d e n t o f t h e n e wgenerating station.The new technology anddiplomacy mixed together whenon May 7, 1956, Admiral Rickoverand AEC Chairman, Lewis L.Strauss, conducted a tour of theconstruction site for the head of Atomic Energy of Great Britain. Visiting the shippingport site as part of a tour of U.S. atomic facilities were Sir Edwin Plowden, Chairman of the United Kingdom Atomic Energy Authority, and John A. V. Willis, Scientific Attache to the British Embassy.The Shippingport Atomic Power Station would be somewhat different from the nuclear power stations that were to follow. From the outset, the Shippingport project was directed towards advancing the basic technology of light water cooled reactors, through design, development, building and testing and operation of a large power reactor as part of a public utility system. Because of this primary mission, the design of the station is markedly influenced by its dual role of test facility and power producer. The test facility concept results in a duplication and isolation of equipment and in a large amount of instrumentation compared to that required solely for operation as a power producer.The Shippingport station was constructed for the purpose of advancing nuclear power technology generally. Accordingly, the entire plant was built along very flexible lines. The reactor portion could accommodate cores of different types, and greater power. Multiple components were made by a number of manufacturers using different designs and materials of various kinds were utilized.Because the technology was new and little understood by the public, some people feared atomic energy. The three principle organizations involved in the Shippingport project, Naval Reactors, Westinghouse Electric Corporation and Duquesne Light Company, patiently explained in both appearances before groups and through the news media, that all safety precautions possible were taken in order to minimize the possibility of the escape of any radiation fromNUCLEAR TECHNOLOGY AND DIPLOMACY mixed together during a tour of the Shippingport constructionsite by representatives of Great Britain. Participating in the tour were from left, Charles H. Weaver, VicePresident, Westinghouse Electric Corporation, Sir Edwin Plowden, Chairman of the United Kingdom Atomic Energy Authority, Lewis L. Strauss, Chairman of the AEC, Philip A. Fleger, Chairman of the Board, Duquesne Light Company, Admiral H. C. Rickover, Chief of the Naval Reactors branch of the AEC, and H. Briggs, Jr., Duquesne’s Manager of Advertising and Public Relations.the plant. They pointed out thatthe difference between Shipping-port and a conventional coal, oilor gas-fired power station was theheat source. The heat was pro-vided by nuclear fission, while inconventional power plants thechemical reaction of the coal, gasor oil with oxygen to produce afire, provided the heat.The station, when completed,consisted of a pressurized waterreactor and associated systems;four steam generators heated bythe reactor; a single turbinegenerator and associated systems;a radioactive waste disposalsystem; laboratories; shops; andadministrative facilities.The reactor and other systemscarrying high pressure waters u b j e c t t o r a d i o a c t i v econtamination were housed inf o u r i n t e r-c o n n e c t e dcontainment vessels in order top r e v e n t t h e p o s s i b i l i t y o fradioactivity escaping into the environment. These inter-connected containment structures of reinforced concrete and steel were buried beneath the ground. The turbine generator was located outdoors on the turbine deck below which were located the bulk of the steam and electrical systems.The Shippingport pressurized water reactor plant consisted of two main parts: a primary system containing the nuclear reactor which produced heat and the water which circulates through the reactor to cool it; and a secondary system, containing other water completely isolated from the reactor, which transferred the heat for use in a steam turbine.At Shippingport, as with all nuclear plants, the nuclear reactor itself is the key element in the primary system. The heart of the nuclear reactor is the core which is housed in the reactor pressure vessel, a large steel container 33 feet high and about 9 feet in diameter.The first core was an assembly of plates and rods arranged in the general shape of a cylinder, 6 feet high and 7 feet in diameter. The plates in the first core were enriched uranium clad with an alloy of zirconium as protection from the hot water; the rods were hollow zirconium alloy tubes filled with natural uranium oxide pellets. The core was a seed and blanket type with the enriched uranium plates constituting the seed and natural-uranium rods the blanket. The reactor contained enough fissionable fuel to form a critical mass: one capable of sustaining a nuclear chain reaction. The chain reaction could be started, stopped and controlled by 32 neutron absorbing control rods made of the element hafnium. When inserted into the reactor, the control rods slowed the chain reaction and lowered the power level: when withdrawn, they allowed the chain reaction to increase, thus increasing the power level.The Shippingport reactor is a pressurized light water reactor. In this type of reactor water is pumped through the primary system to flow around the nuclear fuel elements in the reactor. As it flows around the reactor core, the water absorbs heat from the fissioning nuclear fuel. The whole primary system is kept under high pressure at about 2,000 pounds per square inch, to prevent the primary water from boiling, thus the name pressurized water reactor. When the water is heated,it flows to the heat exchanging steam generators where it gives up the heat to water in the secondary system.The secondary system is a relatively low pressure system so that when it absorbs heat in the steam generator, the water in the secondary system is turned to steam. This steam, when sent to the turbine generator, provides the necessary energy to drive the turbine.The Shippingport reactor has four steam generators where the heat exchange takes place. By using the method of exchanging heat where the primary water is completely isolated, the secondary water does not become contaminated with radioactivity, thus reducing the chance of any escape of radioactivity. The primary system also includes a number of auxilliary systems, such as the pressurizing system, valve operating systems and coolant purification systems.THE FIRST CORE, six feet high and seven feet in diameter,weighing 58 tons and containing 14 tons of natural uranium and 165 pounds of high-enriched uranium, was installed inOctober, 1957. It took more than eight hours to lower the core, the heart of the reactor system, into position with only six-hundredths of an inch clearance between it and the walls of the steel con-tainer known as the pressure vessel.One of the most important features of the Shippingport reactor was that it had a negative temperature coefficient of reac-tivity. This means the reactor inherently tended to maintain the power level at which it was set. If,for example, the temperature of the water entering the reactor dropped for any reason, the reactor automatically produced more heat, and thus a higher outlet water temperature. If the inlet water increased in tempera-ture, the heat output of the reactor automatically dropped. Thus, the reactor itself automatically main-tained the correct power level with no controls being involved.This inherent automatic control feature was true for normal power changes in electric systems so that c o n t r o l -r o d m o v e m e n t w a s necessary only for large changes in power output of the plant.THE AGE OF NUCLEAR POWER came to the Pittsburgh area on Wednesday, December 18, 1957 at 12:39 a.m. Duquesne Light engineers synchronized the turbine-generator at Shippingport with the Duquesne Light system as the first commercial electricity produced by nuclear energy was sent out to customers in the Pittsburgh area. By 7:00 a.m. that morning, the turbine-generator fed by steam produced from the heat of the nuclear reaction was generating more than 12,000 kilowatts of electricity.After more than our years of planning,construction,testingand plenty of plain hard work, the Shippingport reactor was ready for fuel loading. In October, 1957 the reactor core weighing 58 tonsand containing 14 tons of natural uranium and 165 pounds of highly-enriched uranium was installed. It took more than 8 hours tolower the core, the heart of the reactor system, into position withonly six-hundreds of an inch clearance between it and the walls ofthe steel container, known as the pressure vessel. A little more than45 days later the plant was ready for operation. At 4:30 a.m. on December 2, 1957, the control rods were raised just to the point of criticality, where the atomic reaction would maintain itself. This historic milestone for Shippingport also coincided with the 15th Anniversary of the World’s first nuclear fission reactor which was built under the football stadium at The University of Chicago by a group of scientists headed by Enrico Fermi.Testing continued as the reactor was operated below half of itsdesign potential during the next two weeks. Slowly the reactor wasbrought to the point where the pressurized water was steadily producing steam as it circulated through the heat exchange tubes in the steam generator. On Wednesday, December 18, 1957 at 12:30 a.m. the first power was produced at Shippingport Atomic Power Station as engineers synchronized the plant with the Duquesne Light Company system. Thus the first electricity to be generated at Shippingport was fed into the grid that carried electricity throughout the Pittsburgh area.By 7:00 a.m. that morning theShippingport plant was producingmore than 12,000 kilowatts ofelectricity. As with many signifi-cant historic occasions, the eventwas accomplished before mostpeople knew what was happening.The people of Pittsburgh did notknow they were receiving electricenergy generated by the atom asthey made their toast, brewedtheir coffee and read their news-papers. Later the news mediawould inform the public of theimportant event.W i t h i n a f e w d a y s,o nDecember 23, 1957, full power of68,000 kilowatts was attained byt h e S h i p p i n g p o r t r e a c t o r.Although full power was reached,testing continued to determinethe operating characteristics anddependability of the station. On the basis of this early “shakedown” operation, it was found that the reactor and station as a whole equalled or bettered the expectations of its designers and operators and they noted that it operated with an ease and responsiveness surpassing conventional coal-fired stations.THE AGE OF NUCLEAR POWER came to the Pittsburgh area on Wednesday, December 18, 1957 at 12:39 a.m. Duquesne Light engineers synchronized the turbine-generator at Shippingport with the Duquesne Light system as the first commercial electricity produced by nuclear energy was sent out to customers in the Pittsburgh area. By 7:00 a.m. that morning, the turbine-generator fed by steam produced from the heat of the nuclear reaction was generating more than 12,000 kilowatts of electricity.After more than four years of planning, construction, testingand plenty of plain hard work, the Shippingport reactor was ready for fuel loading. In October, 1957 the reactor core weighing 58 tonsand containing 14 tons of natural uranium and 165 pounds of highly-enriched uranium was installed. It took more than 8 hours tolower the core, the heart of the reactor system, into position withonly six-hundreds of an inch clearance between it and the walls ofthe steel container, known as the pressure vessel. A little more than45 days later the plant was ready for operation. At 4:30 a.m. on December 2, 1957, the control rods were raised just to the point of criticality, where the atomic reaction would maintain itself. This historic milestone for Shippingport also coincided with the 15th Anniversary of the World’s first nuclear fission reactor which was built under the football stadium at The University of Chicago by a group of scientists headed by Enrico Fermi.Testing continued as the reactor was operated below half of itsdesign potential during the next two weeks. Slowly the reactor wasbrought to the point where the pressurized water was steadily producing steam as it circulated through the heat exchange tubes in the steam generator. On Wednesday, December 18, 1957 at 12:30 a.m. the first power was produced at Shippingport Atomic Power Station as engineers synchronized the plant with the Duquesne Light Company system. Thus the first electricity to be generated at Shippingport was fed into the grid that carried electricity throughout the Pittsburgh area.By 7:00 a.m. that morning theShippingport plant was producingmore than 12,000 kilowatts ofelectricity. As with many signifi-cant historic occasions, the eventwas accomplished before mostpeople knew what was happening.The people of Pittsburgh did notknow they were receiving electricenergy generated by the atom asthey made their toast, brewedtheir coffee and read their news-papers. Later the news mediawould inform the public of theimportant event.W i t h i n a f e w d a y s,o nDecember 23, 1957, full power of68,000 kilowatts was attained byt h e S h i p p i n g p o r t r e a c t o r.Although full power was reached,testing continued to determinethe operating characteristics anddependability of the station. On the basis of this early “shakedown” operation, it was found that the reactor and station as a whole equalled or bettered the expectations of its designers and operators and they noted that it operated with an ease and responsiveness surpassing conventional coal-fired stations.Speaking for Westinghouse Electric Corporation, Mark W.Cresap, Jr., the Company president said, “The significance of Shippingport is two fold. It is a revolutionary type of station for full-scale electric power generation. It is also a full-scale training and testing facility of historic meaning to the world.“From what is learned here, as this partnership between the Atomic Energy Commission and the Duquesne Light Company progresses, will represent a priceless contribution to the harnessing of nuclear power for the benefit of humanity. For Westinghouse,participation in designing and developing the nuclear reactor has been a most challenging and inspiring experience.”Representing the Congressional Joint Committee on Atomic Energy, the Honorable James E. VanZandt, said “. . . we are developing the atom for peaceful purposes and we are doing it by means of ‘partnership’ between government and industry. All of us can be justifiably proud of this, the first all-commercial reactor. Let us consider it as a monument to American genius and to our system of free enterprise which is the envy of the world.”Lewis L. Strauss, Chairman of the Atomic Energy Commission recalled for the audience the ground breaking ceremony and said, “In the three years and nine months which have elapsed, a period comparing favorably with the time required to build a con- ventional power plant, the co-operative efforts of government and private industry have brought this plant into being. It is the first large-scale power plant yet built to convert the energy within the atom exclusively to the peaceful service of man’s need for electric power to ease his burden and provide additional comfort for his living. While the creation of this great nuclear power station is of historic significance, we are reminded that this is only one facet of a broad program which the world has come to call Atoms for Peace.”Speaking for Duquesne Light,Philip A. Fleger, Chairman of the Board said, “It is altogether fitting that this station should be located close to the birthplace of the petroleum industry and almost on top of one of the world’s greatest coal fields. For the history of SPECIAL CONSTRUCTION was usedin order to industry and man’s progress is minimize the possibility of radioactivity escaping into closely bound up with the history of fuel. Where fuels have been the environment from the reactor and other systems plentiful and man has developed techniques to harness their energy,carrying high pressure water subject to radioactive industry has flourished and people have prospered.contamination. The reactor and other systems were housed in four interconnected containment vessels,one under construction above, whlch were built of reinforced concrete and steel and were buried beneath the ground.the lessons yet to be learned in operating it, Will be applied throughout the free world,” he continued. “Atomic power stations “In a larger sense progress is the real significance of Shippingport. . . . This is true progress. It comes from the work of free country.”printed in scientific and technical journals about the facility. And many papers about its operation and testing programs have been delivered at scientific meetings throughout the world. It has also served as a training ground for many key personnel in nuclear generating plants throughout the world.Shippingport also established itself as a source of electricity to help power the Pittsburgh area. During the life of Shippingport’s first core, from 1957 to 1964, the atomic power station produced almost 2“The lessons already learned in building this pioneer station,now under construction as well as others still to be designed, will be more efficient and more economical because of it.men, with free hands and free minds, in a free society.”The historical significance of Shippingport was well established at its very beginning. Shippingport Atomic Power Station would be more than merely the first full-scale commercial atomic power station. Duquesne’s Chairman of the Board, Philip A. Fleger, said it perhaps best when he said in Los Angeles in 1955: “Shippingport will be a university so to speak for the electric power industry of this Shippingport, as it operated through the years, established itself as a source of valuable information on reactor technology for the entire nuclear power industry. Hundreds of articles have been billion kilowatt-hours of electricity. In 1964 the plant was removed from service for an extended period of time to permit installation of the second core.The new core increased the HISTORIC ACHIEVEMENT RECOGNIZED — The Shippingport Atomic Power Station was honored by the American Society of Mechanical Engineers as a National Historic Mechanical Engineering Landmark on May 20, 1980. This plaque was presented to Shippingport in recognition of its historicalsignificance to the development of the engineering profession and of the nation.plant’s electrical generating capacity from 60,000 to 100,000kilowatts and was capable of producing an additional 50,000kilowatts (equivalent) of heat. It had more than 5 times the design energy output and over twice the power of the original core. Core #2 was instrumented so that infor-mation concerning power distri-bution in the reactor was available for further improvements and refinements in future nuclear power stations. It is important to know how core performance varies over the life of the core so that more powerful longlasting cores can be built at lower costs.From February 3, 1965, when the second core went into operation, until February 4, 1974,Shippingport Atomic Power Station had generated almost 3½billion kilowatt hours of electricity.The plant went out of service on February 4, 1974, because of amechanical failure in the turbine-generator. Meanwhile, as part of。

江苏省苏州市2021-2022高二英语下学期学业质量阳光指标调研(期末)试题注意事项:1.本调研卷分为第I卷(选择题)和第II卷(非选择题),满分150分。
答题时间120分钟。
2.请将第I卷的答案填涂在答题卡上,第II卷请直接在答题卡上规定的地方作答。
答题前,务必将自己的学校、姓名、调研序列号等相关息写在答题卡上规定的地方。
第I卷选择题(共75分)第一部分听力(共两节,满分20分)做题时,先将答案标在调研卷上。
录音内容结束后,你将有两分钟的时间将调研卷上的答案转涂到答题卡上。
第一节(共5小题;每小题1分,满分5分)听下面5段对话,每段对话后有一个小题。
从题中所给的A、B、C三个选项中选出最佳选项,并标在调研卷的相应位置。
听完每段对话后,你都有10秒钟的时间来回答有关小题和阅读下一小题。
每段对话仅读一遍。
1.What does the woman prefer to drink now?k.B.Coffee.C.Water.2.What will the man do first?A.Pick up lunch.B.Visit the bank.C.Go to the post office.3.What happened to the man?A.He lost his cat.B.He was nearly drowned.C.He was caught in the rain.4.What are the speakers probably doing?A.They are on vacation.B.They are shopping.C.They are on business.5.What is the man's opinion about high-speed rail?fortable but expensiveB.Convenient and relaxing.C.Fast butnot enjoyable.第二节(共15小题;每小题1分,满分15分)听下面5段对话或独白。


Home Search Collections Journals About Contact us My IOPscienceThe synthesis and selective gas sensing characteristics of SnO2/α-Fe2O3 hierarchical nanostructuresThis article has been downloaded from IOPscience. Please scroll down to see the full text article.2008 Nanotechnology 19 205603(/0957-4484/19/20/205603)View the table of contents for this issue, or go to the journal homepage for moreDownload details:IP Address: 202.113.13.10The article was downloaded on 09/09/2012 at 03:11Please note that terms and conditions apply.IOP P UBLISHING N ANOTECHNOLOGY Nanotechnology19(2008)205603(5pp)doi:10.1088/0957-4484/19/20/205603The synthesis and selective gas sensing characteristics of SnO2/α-Fe2O3 hierarchical nanostructuresYujin Chen1,2,4,Chunling Zhu3,Xiaoling Shi2,Maosheng Cao2andHaibo Jin21College of Science,Harbin Engineering University,Harbin15001,People’s Republic ofChina2School of Materials Science and Engineering,Beijing Institute of Technology,Beijing100081,People’s Republic of China3College of Materials Science and Chemical Engineering,Harbin Engineering University,Harbin150001,People’s Republic of ChinaE-mail:chenyujin@Received28January2008,infinal form10March2008Published14April2008Online at /Nano/19/205603AbstractSnO2/α-Fe2O3hierarchical nanostructures,in which the SnO2nanorods grow on the sidesurface ofα-Fe2O3nanorods as multiple rows,were synthesized via a three-step process.Thediameters and lengths of the SnO2nanorods are6–15nm and about120nm.The growthdirection of SnO2nanorods is[001],significantly affected by that ofα-Fe2O3nanorods.Thehetero-nanostructures exhibit very good selectivity to ethanol.The sensing characteristics arerelated to the special heterojunction structures,confirmed by high-resolution transmissionelectron microscopy observation.Therefore,a heterojunction barrier controlled gas sensingmechanism is realized.Our results demonstrate that the hetero-nanostructures are promisingmaterials for fabricating sensors and other complex devices.1.IntroductionHierarchical hetero-nanostructures are expected to have novel or multifunctional properties and have applications in the fabrication of complex nanodevices.In recent years, great progress has been made in the synthesis of such nanocomposites[1–7].Hierarchical ZnO nanostructures were synthesized by a vapor transport and condensation technique. In the nanocomposites,the secondary ZnO nanorods grew either as a single row or multiple rows on the side surfaces of the core In2O3nanowires[1].Xie et al reported the epitaxial growth of T-ZnO/SnO2hetero-nanostructures and found that the new luminescence properties were induced by the epitaxial interfaces[2].Recently,Fen and his co-workers grew carbon nanotubes,ZnO nanowires and silicon nanowires on carbon cloth[3–5].These nanocomposites exhibited enhancedfield emission properties and are expected to befield emitters with practical applications in highly efficient lamps,field emission displays,and micro vacuum electron sources.4Author to whom any correspondence should be addressed.As two important kinds of fundamental material, SnO2[8–18]andα-Fe2O3[20–26]have attracted a lot of attention due to their applications as chemical catalysts and devices such as sensors[16–18],UV detectors and transistors[19],etc.In this paper,we report a three-step route to grow SnO2nanorod arrays on the side surfaces of porousα-Fe2O3nanorods.The gas sensing properties and sensing mechanism of the SnO2/α-Fe2O3hierarchical hetero-nanostructures are investigated.2.Experimental detailsSnO2/α-Fe2O3hierarchical hetero-nanostructures were synthe-sized via a three-step process.First,β-FeOOH nanorods were synthesized by a hydrothermal route.In a typical procedure, 40ml of0.5M FeCl3solution was transferred into a50ml Teflon-lined autoclave.The autoclave was heated to120◦C, and kept for12h.After cooling to room temperature,the resulting precipitates were washed several times with water and absolute ethanol,and then dried at80◦C for6h.AfterFigure1.(a)SEM image ofβ-FeOOH nanorods.(b)SEM image of α-Fe2O3nanorods.(c)XRD pattern of the products.Pattern1:β-FeOOH,pattern2:α-Fe2O3nanorods,pattern3:SnO2/α-Fe2O3 hierarchical hetero-nanostructures.(d)SEM image ofα-Fe2O3 nanorods.calcinations of the obtainedβ-FeOOH nanorods at500◦C for 2.5h,α-Fe2O3nanorods with porous structures were obtained. The SnO2/α-Fe2O3hierarchical hetero-nanostructures werefi-nally prepared usingα-Fe2O3nanorods and SnCl4as starting materials through a hydrothermal method as follows.0.015g α-Fe2O3nanorods were dispersed into a mixed solution con-sisting of heptane(10.2ml),hexanol(3.0ml),sodium dodecylsulfate(SDS,1.44g),Sn(OH)2−6(2.0ml;the volume ratio of0.5M SnCl4and5M NaOH solutions is1:3).After ultra-sonication for5min,the mixture was transferred into a25mlTeflon-lined autoclave and heated to220◦C for6h.After cool-ing to room temperature,the obtained precipitates were washedseveral times with absolute ethanol and distilled water.The fabrication process of the sensors based on the hetero-nanostructures has been described elsewhere[16,17,27–30].The sensitivity(S)in this paper is defined as S= R a/R g[16,17],where R a is the sensor resistance in air,and R g is the resistance in target–air mixed gas.During the measuredprocess the ambient relative humidity(RH)is about30%. 3.Results and discussion3.1.Morphology study of hetero-nanostructuresβ-FeOOH nanorods can be obtained at temperatures in the range from90to120◦C.Samples prepared at different synthesis temperatures have slightly different morphologies. Figure1(a)is a typical scanning electron microscopy(SEM) image of the as-synthesizedβ-FeOOH nanorods that were synthesized at120◦C.It indicates that the diameters and the lengths of the nanorods are50–180nm and∼2μm, respectively.Pattern1infigure1(c)shows corresponding x-ray diffraction(XRD)pattern from the pared with the data in JCPDS card(No.75-1594),all the diffraction peaks in the pattern can be indexed toβ-FeOOH.After calcinations at500◦C for2.5h,β-FeOOH nanorodswere Figure2.(a)Low-resolution TEM image of SnO2/α-Fe2O3 hierarchical hetero-nanostructures.Scale bar:100nm.(b)High-resolution TEM image of an individual SnO2nanorod grown onα-Fe2O3nanorods.Scale bar:5nm.(c)High-resolution TEM image of the interface of SnO2/α-Fe2O3hierarchicalhetero-nanostructures.transformed intoα-Fe2O3nanorods,proved by XRD pattern 2infigure1(c)(JCPDS No.33-0664).They remained in the original morphologies(figure1(b)).Pattern3is the measured XRD pattern of thefinal products.The asterisks indicate the peaks coming fromα-Fe2O3(JCPDS No.33-0664),while the others come from SnO2(JCPDS No.41-1445).The results indicate that thefinal products were composites ofα-Fe2O3and SnO2.Figure1(d)shows a typical SEM image of thefinal products.It can be clearly observed that thefinal products exhibit hierarchical heterostructures,in which the second nanorod arrays grow on the side surface of theα-Fe2O3 nanorods as multiple rows.According to the XRD results,the secondary phase in the structures is SnO2nanorods.The morphology and size of the hierarchical hetero-nanostructures were further characterized by transmission electron microscopy(TEM).Figure2(a)displays a typical TEM image of the heterostructures.It is found that the average diameters and lengths of the SnO2nanorods are6–15nm and about120nm,respectively.It is clear that the SnO2nanorods are parallel to each other at the same side surface of anα-Fe2O3nanorod,and the angle between the SnO2andα-Fe2O3 nanorods is about122◦or58◦.3.2.Crystallographic study of hetero-nanostructuresIn order to clarify how SnO2nanorods grow onα-Fe2O3 nanorods,high-resolution TEM(HRTEM)observations of hi-erarchical hetero-nanostructures were conducted.Figure2(b) shows the HRTEM image of an individual SnO2nanorod.The two adjacent spacings are0.246and0.237nm,correspond-ing to(101)and(200)planes of SnO2,respectively.This in-dicates the single-crystal nature of the nanorods with prefer-ential growth along the[001]direction.The result is differ-ent from the previous report by Zhang et al that[101]is thepreferential growth direction for SnO 2nanorods grown on α-Fe 2O 3nanotubes [11].This may be related to the growth direc-tion of α-Fe 2O 3nanorods because the growth of heterostruc-tures requires low lattice mismatch between different materi-als [2,11].Figure 2(c)is an HRTEM image of the interface of hetero-nanostructures.It is obvious that the SnO 2nanorods still grow along the [001]direction at the interface.For α-Fe 2O 3nanorods,the two almost identical spacings of 0.252nmare consistent with the d values of the (2¯10)and (110)planes,indicating that the α-Fe 2O 3nanorods grow preferentially along the [110]direction.Therefore,the interfacial orientation relationships between the SnO 2and α-Fe 2O 3nanorods are(101)SnO 2/(110)α-Fe 2O 3and (200)SnO 2/(2¯10)α-Fe 2O 3.The lat-tice mismatch is lower for such interfacial orientation relation-ships [11],thereby resulting in SnO 2nanorods with growth along the [001]rather than [101]direction.The result demon-strates that the growth direction of the second phase can be affected by the first phase in a heterogeneous system.This may be an effective route to synthesize heterostructures with controlled crystalline and physical characteristics because dif-ferent growth directions may lead to different physical proper-ties.For example,SnO 2nanoribbons with exposed (10¯1)and (010)surfaces have highly sensing characteristics to NO 2even at room temperature [31].The experimentally measured angle between the (101)and (200)planes of SnO 2is about 56.2◦,while it is about60◦between the (2¯10)and (110)planes of α-Fe 2O 3,as shown in figure 2(c).Both values are consistent with the theoretically calculated values,suggesting that the misfit angle between those planes is about 3.8◦.According to the HRTEMobservation (figure 2(c)),however,the (2¯10)α-Fe 2O 3plane is not completely parallel to the (200)SnO 2plane:the acute angle between them is about 2◦.Therefore,the misfit angle between the (110)α-Fe 2O 3plane and the (101)SnO 2plane is about 1.8◦.The results reveal that SnO 2nanorods grown on α-Fe 2O 3nanorods should be parallel to each other at the same side surface and the angle between SnO 2nanorods and α-Fe 2O 3nanorod should be 122◦or 58◦,which is consistent with the low-resolution TEM observation (figure 2(a)).In addition,lattice distortion induced by lattice mismatch is also observed,as indicated by the white frame in figure 2(c).The phenomenon is also observed by Zheng et al in the ZnO/SnO 2system [2].It should be noted that the synthesized conditions have an important effect on the formation of SnO 2/α-Fe 2O 3hierarchical hetero-nanostructures.When the volume ratio of SnCl 4and NaOH was changed to 1:1or 1:2,the hierarchical hetero-nanostructures mentioned above were difficult to obtain.In those cases,only SnO 2nanoparticles or short rods grew on the side surfaces of α-Fe 2O 3nanorods.The same results were also observed when the solvents were changed into a mixture of ethanol and water,or without the addition of SDS.Therefore,the volume ratio of SnCl 4and NaOH,the solvents and SDS are important factors for the growth of SnO 2/α-Fe 2O 3hierarchical hetero-nanostructures.3.3.Gas sensitivity of hetero-nanostructuresIt is well known that SnO 2and α-Fe 2O 3are important sensing materials.However,gas sensors for practicalapplicationsFigure 3.The sensitivities of SnO 2/α-Fe 2O 3hetero-nanostructures to ethanol at different working temperatures:curve 1,at 350◦C;curve 2,at 250◦C.The inset shows the sensitivities of SnO 2/α-Fe 2O 3hetero-nanostructures to H 2,CH 4and C 4H 10gases at a working temperature of 350◦C.require not only high sensitivities,but also very good selectivity to target molecules.The hierarchical hetero-nanostructures prepared in this work exhibited very good selectivity as well as high sensitivity to ethanol.Figure 3shows the sensitivities of the hetero-nanostructures to ethanol with various concentrations at different working temperature.At a working temperature of 350◦C,the sensitivity is 4.6–10ppm,as shown in curve 1,while the sensitivities are less than 2.3–1000ppm of H 2,CH 4and C 4H 10as shown in the inset of figure 3.As the working temperature was reduced to 250◦C,the hetero-nanostructures still kept high sensitivity.The sensitivity is up to 2.9–10ppm of ethanol,as shown in curve 2;however,the hetero-nanostructures have no response to the three other gases.The results demonstrate that the hetero-nanostructures are promising materials for fabricating gas sensors with good sensing characteristics.For a pure metal oxide semiconductor,the sensing mecha-nism is usually explained by the space–charge model [16,17].In this model,the sensitivity of the materials is strongly de-pendent on the change of the space–charge length under differ-ent gas atmospheres,resulting in higher sensitivity of the sens-ing materials with smaller size [16,17].However,the elec-tron transport mechanism of the nanocomposites synthesized in this work should be significantly different from that of the pure SnO 2or α-Fe 2O 3nanorods due to the formation of many semiconductor heterojunctions at their interfaces,observed by HRTEM,as shown in figure 2(c).Electron transport is thus expected to be strongly tuned by the heterojunction (HJ)bar-rier,which has been widely investigated for many HJ devices such as lasers,photodiodes and HJ bipolar transistors.It is well known that the band gaps of SnO 2and α-Fe 2O 3at room temperature are about 3.6eV and about 2.1eV [12,32,33].Thus the energy band structure of the SnO 2/α-Fe 2O 3HJ can be schematically depicted in figure 4,where φeff denotes the effective barrier height,considering the contribution of other factors such as temperature to the barrier height [34].There-fore,it requires that the transport of electrons should overcome the heterojunction barriers [34].At the high temperature re-gion,the electron motility μis expressed by [34]μ=μ0exp (−q φeff /k B T ),(1)Figure4.A schematic diagram of the energy band structure of the SnO2/α-Fe2O3hetero-nanostructures:(a)in air,(b)in ethanol. where q is the charge of an electron,k B is Boltzmann’s constant,and T is the absolute temperature.According to equation(1),the conductivity G of the heterostructures under different gas atmospheres can be given by[34]G=G0exp(−φeff/k B T).(2) In this equation,G0can be considered as a constant parameter.φeff will increase in air because electrons are trapped both in SnO2andα-Fe2O3induced by adsorbed oxygen species,as shown infigure4(a).In this case,the conductivity of the HJ is very low.When the heterostructures are exposed to ethanol,the reaction between the adsorbed oxygen species and the ethanol molecules leads to the release of the trapped electrons back simultaneously into the conduction bands of the SnO2and Fe2O3rods,resulting in a decrease in the width and height of the barrier potential at their interfaces,as shown infigure4(b).In the case,the conductivity of the HJ will consequently be greatly increased,resulting in high sensitivity of the SnO2/α-Fe2O3HJs to ethanol. Therefore,the barrier with adjustable height controls the transport of electrons in the heterostructures,and accordingly controls the sensing characteristics of the nanocomposites.As for the good selectivity of the hetero-nanostructures,it may be related to the following factors.First,for pure SnO2 sensors the sensitivity to H2exhibited significantly different characteristics,depending on the fabrication techniques of both the sensing materials and the sensors.For example, W¨o llenstein et al fabricated a micromachined micro-hotplate sensor array based on ZnO,WO3and V2O5,in which the sensitivity of SnO2to100ppm H2is less than3.0at a working temperature of400◦C[35].Salehi et al reported that SnO2 sensingfilms had a sensitivity to1000ppm H2less than4.0 at200◦C[36].But the sensitivity of sensors based on SnO2 thickfilms with porous structures were above400–800ppm H2at350◦C[37].The sensitivity was as high as1000–800ppm H2at350◦C after SnO2nanoparticles were calcined at600◦C[38].This phenomenon may result from the size and crystalline phase of SnO2sensing materials and the sensor configurations.Therefore,the sensors based on the hetero-nanostructures presumably exhibited low sensitivity to H2. Second,the sensing properties of composites are significantly different from those of the single-component counterparts. For instance,CeO/SnO2and SnO2/CuO composites had excellent selectivity to H2S[39,40].SnO2/ZnO[41]and SnO2/α-Fe2O3[42,43]composites exhibited greatly improved sensitivity to ethanol vapor compared to H2,CH4and CO molecules.The reason for this is that the sensing mechanism of the composites is different from that of the single-component counterparts[43,44].It has been reported that when the test gas has a complex molecular structure or a reactive functional group such as ethanol,the surface reactions could differ depending on the acid–base properties.The base properties are more helpful to the surface reactions between C2H5OH and surface O species.In this work,the hetero-nanostructures are composed of SnO2andα-Fe2O3nanorods.They are basic oxides,and thereby the basicity of the hetero-nanostructures will increase.Due to the fact mentioned above,the SnO2/α-Fe2O3hetero-nanostructures exhibited higher sensitivity to ethanol than to H2,CH4and C4H10gases.In addition,water vapor cross-sensitivity is important for gas sensors based on metal oxides.Thus the response of the sensors was also measured at the RH of∼30%and∼54%, respectively.Negligible resistance changes of the sensors with the present configuration at these humidities were observed. Therefore,the sensors based on the hetero-nanostructures can be used to detect ethanol vapor.4.ConclusionsIn summary,we synthesized SnO2/α-Fe2O3hierarchical hetero-nanostructures by a three-step process.The SnO2 nanorods with average diameters in the range6–15nm and lengths of120nm grow on the side surface of theα-Fe2O3 nanorods as multiple rows,resulting in the formation of hierarchical structures.It is observed that the preferential growth direction of SnO2nanorods is[001],significantly affected by the growth direction ofα-Fe2O3nanorods.This may offer an effective route to synthesize heterostructures with controlled crystalline and physical characteristics.The hetero-nanostructures exhibit high sensitivity and very good selectivity to ethanol.Heterojunction barrier controlled gas sensing characteristics were realized in SnO2/α-Fe2O3 hierarchical nanostructures.Our results demonstrate that SnO2/α-Fe2O3hetero-nanostructures are very promising for fabricating sensors as well as other complex devices. AcknowledgmentsThe authors acknowledge the support from the National Natural Science Foundation of China(50772025),the PhD Programs Foundation of the Ministry of Education of China(20070217002),China Postdoctoral Science Foundation (20060400042),and also the Innovation Foundation of Harbin City(RC2006QN017016).References[1]Lao J Y,Wen J G and Ren Z2002Nano Lett.21287[2]Kuang Q,Jiang Z Y,Xie Z X,Lin S C,Lin Z W,Xie S Y,Huang R B and Zheng L S2003J.Am.Chem.Soc.12511306[3]Banerjee D,Jo S H and Ren Z F2004Adv.Mater.162028[4]Jo S H,Wang D Z,Huang Z Y,Li W Z,Kempa K andRen Z F2004Appl.Phys.Lett.85801[5]Zeng B Q,Xiong G Y,Chen S,Wang W Z,Wang P Z andRen Z F2007Appl.Phys.Lett.90033112[6]Zhang D F,Sun L D,Jia C J,Yan Z G,You L P andYan C H2005J.Am.Chem.Soc.12713492[7]Wan Q,Wei M,Zhi D,MacManus-Driscoll J L andBlamire M G2006Adv.Mater.18234[8]Wu N L,Wang S Y and Rusakova I A1999Science2851375[9]Comini E,Faglia G,Sberveglieri G,Pan Z and Wang Z L2002Appl.Phys.Lett.811869[10]Law M,Kind H,Messer G,Kim F and Yang P D2002Angew.Chem.Int.Edn412405[11]Zhang D F,Sun L D,Yin J L and Yan C H2003Adv.Mater.151022[12]Dai Z R,Pan Z W and Wang Z L2003Adv.Funct.Mater.139[13]Chen Y J,Li Q H,Liang Y X,Wang T H,Zhao Q andYu D P2004Appl.Phys.Lett.855682[14]Cheng B,Russell J M,Shi W S,Zhang L andSamulski E T2004J.Am.Chem.Soc.1265972[15]Chowdhuri A,Gupta V,Sreenivas K,Kumar R,Mozumdar S and Patanjali P K2004Appl.Phys.Lett.841180[16]Chen Y J,Xue X Y,Wang Y G and Wang T H2005Appl.Phys.Lett.87233503[17]Chen Y J,Xue X Y,Wang Y G and Wang T H2006Appl.Phys.Lett.88083105[18]Kuang Q,Lao C S,Wang Z L,Xie Z X and Zheng L S2007J.Am.Chem.Soc.1296070[19]Dattoli E N,Wan Q,Guo W,Chen Y B,Pan X and Lu W2007Nano Lett.72463[20]Rockenberger J,Scher E C and Alivisatos A P1999J.Am.Chem.Soc.1211595[21]Park J,An K,Hwang Y,Park J G,Noh H J,Kim J Y,Park J H,Hwang N M and Hyeon T2004Nat.Mater.3891[22]Wang D S,He J B,Rosenzweig N and Rosenzweig Z2004Nano Lett.4409[23]Jia C J,Sun L D,Yan Z G,You L P,Luo F,Han X D,Pang Y C,Zhang Z and Yan C H2005Angew.Chem.Int.Edn444328[24]Xiong G,Joly A G,Holtom G P,Wang C M,McCready D E,Beck K M and Hess W P2006J.Phys.Chem.B11016937 [25]Wu J J,Lee Y L,Chiang H H and Wong D K P2006J.Phys.Chem.B11018108[26]Li L,Chu Y,Liu Y and Dong L H2007J.Phys.Chem.C1112123[27]Liang Y X,Chen Y J and Wang T H2004Appl.Phys.Lett.85666[28]Chen Y J,Zhu C L and Wang T H2006Nanotechnology173012[29]Chen Y J,Zhu C L and Xiao G2006Nanotechnology174537[30]Chen Y J,Zhu C L and Xiao G2008Sensors Actuators B129639[31]Maiti A,Rodriguez J A,Law M,Kung P,McKinney J R andYang P2003Nano Lett.31025[32]Preisinger M,Krispin M,Rudolf T,Horn S andStrongin D R2005Phys.Rev.B71165409[33]Gratzel M2001Nature414338[34]Weis T,Lipperheide R,Wille U and Brehme S2002J.Appl.Phys.921411[35]W¨o llenstein J,Plaza J A,Can´e C,Min Y,B¨o ttner H andTuller H L2003Sensors Actuators B93350[36]Salehi A and Gholizade M2003Sensors Actuators B89173[37]Sakai G,Baik N S,Miura N and Yamazoe N2001SensorsActuators B77116[38]Baik N S,Sakai G,Miura N and Yamazoe N2000SensorsActuators B6374[39]Fang G J,Liu Z L,Liu C Q and Yao K L2000SensorsActuators B6646[40]Chowdhuri A,Gupta V,Sreenivas K,Kumar R,Mozumdar S and Patanjali P K2004Appl.Phys.Lett.841180[41]de Lacy Costello B P J,Ewen R J,Guernion N andRatcliffe N M2002Sensors Actuators B87207[42]Gopal Reddy C V,Cao W,Tan O K and Zhu W2002SensorsActuators B81170[43]Tan O K,Cao W,Zhu W,Chai J W and Pan J S2003SensorsActuators B93396[44]Yamazoe N and Miura N1992Chemical Sensor Technologyvol4,ed S Yamauchi(Amsterdam:Elsevier)pp39–40。

1Does not qualify for DLC4 Cannot be used w/ FMB, HUB, or HOOKOne needed per project Must be used with FSIR for commissioningSensor Mounted on centerAvailable for EDV-8L–24L only 5Mounted externally to fixture 8Must be used with ES/PCD Only available with FRL driver channelAvailable for EDV-30L–60L onlyMust be used with ES/PCD and ES/IR 9 Reduces published performance by 16%Sensor Mounted off end cap of fixtureConsult Factoryoptions for commissioningSUITABLE APPLICATIONS • Gymnasiums • Manufacturing Plants • Warehouses • Distribution CentersIK10RATING Project Name:Part Number:Type:FEATURES• Sleek Curved Polycarbonate Injection Molded End Caps • Aluminum Body with steel channel for added rigidity • Toolless Hinged Driver Access • Wide & Narrow Distributions• Impact Resistant and IP5X Rated Lens Options:- Frosted Acrylic (FRL)- Frosted Polycarbonate (PFL) IK10 Impact Rated • V-Clips for dual point chain or cable hanging (std.)• Advanced and Basic Controls Options • -40˚F –122˚F Ambient Temperature Range • 3000K - 5000K CCT • Uplight Options• >80 Color Rendering Index (CRI)• Calculated L 70 >100,000 hrs• 0-10V Dimmable Driver (100%–10%)• 120-277V Universal Voltage • 5 Year Warranty• ETL Listed for Damp Location• DesignLights Consortium ® Premium Qualified LuminaireFixtures shown with Frosted Lens (FRL)1 2 3SPECIFICATIONSCONSTRUCTIONAluminum body with steel driver channel for maximum rigidity and thermal properties. Sleek curved polycarbonate injection moldedend caps provide a modern finish while adding rigidity and durability. Toolless hinged driver door access for ease of maintenance and installation. Power access plate mounted on back of center driver channel for easy access to wiring. Product proudly assembled in the United States of America.ELECTRICALClass 2 LED light engine for high efficacy and long life.Calculated L70 > 100,000 hrs. LED Lumen Maintenance Estimates based on TM-21 projections for the light source at 25˚C ambient. Power Factor > 0.90. Standard 120-277 Vac 0-10V dimmable (10%-100%) with optional 347-480 Vac step down transformer. Optional emergency battery backups and generator transfer device available to meet critical life safety lighting requirements. Fixture reduced maximum ambientfor emergency battery backup and step down options. Optional uplight wired to sensor it ordered together.MOUNTINGOptions include suspension chain, aircraft cables, hook, single pendant (HUB), surface mount (SMZBKIT). Reduced maximum ambient range for surface mount.WARRANTY5 Year Warranty (Terms and Conditions apply)OPTICSStandard without lens. Optional frosted acrylic (FRL) and frosted polycarbonate (PFL) lenses available for added LED protection and glare reduction. Both optics are IP5X rated; protected against dust intrusion. Frosted Polycarbonate lens (PFL) is IK10 Impact Rated; highest impact rating available. Wide (std.) & Narrow (ND) Distribution options available for all lumen packages.THERMALAmbient temperature range -40°F to 122°F (-40°C to 50°C). Fixtureto be mounted at a minimum plenum height of 18" to maintain maximum ambient temperature. Reduced maximum ambient for surface mounting.CONTROLSFactory installed Advanced and Basic integral occupancy sensor options available. All Basic sensors are end mounted. Advanced EasySense sensor is end mounted. All other Advanced sensors are center driver cover mounted. Inquire with manufacturer for additional controls options, including customer supplied items. CERTIFICATIONSETL listed to US and Canadian safety standards. Suitable for damp locations. American Recovery and Reinvestment Act Funding Compliant. DLC (DesignLights Consortium) Premium Qualified, with some Standard Qualified configurations. Please refer to the DLC Qualified Products List at /QPL to confirm specific product qualifications.FIOSStandard on/off motion sensor. Comes with 360 degree high bay lens, 360 degree low bay lens & aisle lens. 360 degree high bay lens factory installed, unless notified otherwise. Sensor mounted off end cap of fixture. Inquire for mounting on center of driver cover.Philips ES/HBEasySense sensor combines occupancy sensing, daylight harvesting and institutional tuning in one compact solution. This is an ideal control for any high bay application. Each device allows you to turn on/off occupancy and/or daylight harvesting functions as well as set the full light output, background light level, occupancy prolong time, occupancy grace fading time & occupancy time out/hold time. Requires use of EasySense Pre-Programmed Control Device (ES/PCD) & Commissioning IR Blaster (ES/IR); one of each needed per project. (Sold separately). Sensor mounted off end cap of fixture.USBDUser Selectable Bi-Level Dimming. Allows the end user to manually select the unoccupied dimming level. Once motion is detected, the fixture will return to 100% light output. Sensor mounted off end cap of fixture.Enlighted 5S/CU/CLBundled System with Enlighted 5E sensor (SU-5S-H-CL), Control Unit (CU-4E-FM) & Cable (CBL-5E-CU4-30N). Fixtures are controlled based on occupancy & ambient light. Includes individual lighting control plus connection to Enlighted Energy Manager. The Energy Manager provides data analysis for all data collected by the Enlighted Smart Sensors for energy, occupancy & environment. Provided factory installed components only. Enlighted system provided by others. Contact factory for more information. Sensor mounted on center driver channel.FIOSPCMotion sensor with the addition of photocell control. When installed, fixture will not activate unless 1.motion is detected and 2.light levels are below the preset level. When activated, fixture will turn on to 100% light output & will only shut off when one of the two levels mentioned above are met. If no motion is detected & light levels fall below preset level the fixture remains off. Sensor mounted off end cap of fixture.Legrand FSP-211FSP-211 offers fully adjustable high and low dimmed light levels with optional photocell. Hold off setpoint with automatic calibration option. Adjustable time delay and cut off delay. Sensor adjustable programming with handheld wireless configuration tool; FSIR-100. (Sold separately. One needed per project). Sensor mounted on center driver channel.BDxxFactory set Bi-Level Dimming. Sensor is set to dim level, as indicated in the “xx” portion of the part number, when the area is unoccupied. Example: BD20 sets the fixture to dim down to 20% lumen output when the area is unoccupied. Once motion is detected, the fixture will return to 100% light output. Sensor mounted off end cap of fixture.Sensor mounted off end cap of fixture.Enlighted 5S/CU/IOTBundled System with Enlighted 5E sensor (SU-5S-H-IOT), Control Unit (CU-4E-FM) and Cable (CBL-5E-CU4-30N). Fixtures are controlled based on occupancy and ambient light. Includes individual lighting control plus connection to Enlighted Energy Manager. The Energy Manager provides data analysis for all data collected by Enlighted Smart Sensors for energy, occupancy & environment. Additionally, the IOT functionality takes the data access and function beyond just lighting to a wide range of applications beyond the standard connected lighting controls. Provided factory installed components only. Enlighted system provided by others.Contact factory for more information. Sensor mounted on center driver channel.FIOSPC/DLHFactory installed motion sensor with the addition of photocell control that will allow for daylight harvesting. When installed, fixture will not activate unless 1.motion is detected & 2.light levels are below the preset level. When activated, fixture will turn on only enough to meet the preset light levels and will adjust as the amount of light in the space adjusts between 100%-10%. If light levels are adequate the fixture will remain off. Sensor mounted off end cap of fixture.Legrand FSP-311FSP-311 offers fully adjustable high and low dimmed light levels with optional photocell. Hold off setpoint with automatic calibration option. Adjustable time delay and cut off delay. Sensor adjustable programming via iOS ®or Android ®'Sensor Configuration' App.Sensor mounted on center driver channel. Sensor mounted on center driver channel.BDxxPCFactory set Bi-Level Dimming with motion and photocell. Sensor is set to dim level, as indicated in the “xx” portion of the part number, when the area is unoccupied and will not return to 100% light output unless 1.motion is detected and 2.the light level is below the preset level. Example: BD20PC sets the fixture to dim down to 20% lumen output when the area is unoccupied and will not return to 100% light output unless the above conditions are met for both motion and light level readings. Sensor mounted off end cap of fixture.DHPCDaylight Harvesting off photocell. Fixtures will continually dim up/down throughout the day based on light level readings taken from the photocell control. Sensor mounted off end cap of fixture.Enlighted 5S/CU/ILBundled System with Enlighted 5E sensor (SU-5S-H-IL), Control Unit (CU-4E-FM) and Cable (CBL-5E-CU4-30N). Fixtures are controlled based on occupancy and ambient light. Includes ability to group fixtures together and connect to a switch to zone fixtures for group operation. Provided factory installed components only. Enlighted system provided by others. Contact factory for more information. Sensor mounted on center driver channel.CONTROLS OPTIONSBASICADVANCEDHUB 3/4Conduit Hub 3/4" for Pendant MountingHOOKCast Iron Hook for SinglePoint MountingSMBZKITSurface Mount Bracket Kit providesa 2" space above the fixtureHB-XX-18Y-PADIncludes 2 adjustable cable hangers. Available in 60", 120", 180" & 240"FMBIncludes rigid box to attach to the fixture, provides for single point mounting to accept a pendant, hook or conduit hub(sold separately)CORDxCords available in 6ft, 10ft, 12ft & 20ft (Inquire for additional options) Plugoptions availableQDCxQuick Disconnect Cords available in 6ft, 10ft, 12ft & 20ft (Inquire foradditional options) Plugoptions available.WC11 GA Wire CageOPTIONSLINE DRAWINGS8L - 24L30L36L - 60LQuadralaterally Sy mmetric Solid: 0 DegreesSm all Dash: 45 Degrees Large Dash: 90 DegreesQuadralaterally Sy mmetric Solid: 0 DegreesSm all Dash: 45 Degrees Large Dash: 90 DegreesQuadralaterally Sy mmetric Solid: 0 DegreesSm all Dash: 45 Degrees Large Dash: 90 Degrees。

May 20201FEATURES•Repeatable - self-restoring, nothing to replace, testable •Versatile - various temperature settings available •Durable - long lasting stainless steel shell•Economical - wide spacings reduce installation costs •Factory set•Internal contact area hermetically sealed in stainless steel shell •ROHS CompliantAPPLICATIONS•Protection of schools, factories, offices, libraries, or other non-residential buildings •Power generation •Gas station islands •Paint spray booths •Range hoods •Engine compartmentsDESCRIPTIONDETECT-A-FIRE ® (D-A-F) detectors are the "heart" of many fire protection systems. The highly reliable D-A-F has been the standard for over 75 years. The D-A-F is used for signaling overheat or fire conditions. In the vast majority of applications, the D-A-F provides the ini-tial heat sensing that is used to activate suppression systems using clean agent, CO 2, inert gases, wet or dry chemicals or water.D-A-F detectors are widely accepted, because they are designed with rate compensation. This provides a unique advantage over both fixed temperature and rate-of-rise types of detectors because the D-A-F detector accurately senses the surrounding air temperature regardless of the fire growth rate. At the pre-determined set point, the system is activated.Fixed temperature detectors must be completely heated to alarm temperature and therefore a lag in response time may occur with a fast rate fire. Rate-of-rise devices, on the other hand, are triggered by the rate of increase in ambient temperature and are subject to false alarms caused by harmless, transient thermal gradients such as the rush of warm air from process ovens.The key to the detector's sensitivity is in the design (Figure 1). The outer shell is made of a rapidly expanding alloy which closely follows changes in surrounding air temperature. The innerstrutsare made of a slower expanding alloy. Designed toresist thermal energy absorption and sealed inside the shell, the struts follow temperature changes more slowly.A slow rate fire (Figure 2) will heat the shell and struts together. At the "set point", the detector will trigger, actuating the alarm or releasing the extinguishant.A transient rush of warm air up to 40°F/min. may expand the shell, while not triggering the detector. By ignoring transient warm air excur-sions, the D-A-F detector virtually eliminates false alarms prevalent with rate-of-rise devices.If a fast rate fire (Figure 3) starts, the shell will expand rapidly. The struts will close, actuating the alarm and/or releasing the agent. The faster the fire rate of growth, the sooner the D-A-F detector will react.Figure 1. Ready Figure 2. Slow Fire Figure 3. Fast FireDETECT-A-FIRE ®Detection and Release DevicesVERTICAL DETECT-A-FIRE DETECTOR SPECIFICATIONSVertical D-A-F detectors are designed for use in both "ordinary" or "hazardous" locations. For "ordinary" use, they may be mounted to any approved junction box with 7/8" diameter opening by using 1/2-14 NPT mounting nuts. The device may be wired in or out of con-duit, depending on local preferences and codes. To facilitate supervision of system wiring, four lead wires are provided on normally open vertical detectors (that close on temperature rise). When mounted in a appropriately classified mounting box (i.e. Figure 7), detectors are Underwriters Laboratory and Underwriters Laboratory of Canada listed, and FM Approved for hazardous locations.VERTICAL MODELSTable 1: Model Numbers 27121, 28021, 27120*, 28020*°F Setting °F Tolerance °C Setting °C Tolerance Spacing (in feet)RTIColor Coding UL ULC FM140+7/-860+4/-5505020V-Fast Black 160+7/-871+4/-5252520V-Fast Black 190+7/-888+4/-5505025V-Fast White 210+7/-899+4/-5255025V-Fast White 225+7/-8107+4/-5255025V-Fast White 275±10135±6255025V-Fast Blue 325±10163±6505025V-Fast Red 360±10182±8255030V-Fast Red 450±15232±10255030V-Fast Green 500±15260±10505030V-Fast Orange 600**±20316±12N/A 5030V-Fast Orange 725**±20385±12N/A5030V-FastOrangeNotes:•For clean agents or CO2 suppression systems, ceiling spacing is 20ft. apart unless otherwise specified.•For NFPA Guidelines on ceiling height compensation, see Table 7.•*27120 and 28020 are normally closed devices and do not meet the requirements of NFPA-72 for useas initiating devices (they are 2-wire devices).•*27120 and 28020 are not listed by FM with RTI.•** Not available for Normally Closed detectorsTable 2: Vertical D-A-F SpecificationModel NumberHead MaterialContact Operation Electrical Rating (Resistive Only)27120-0Brass Normally Closed (Open on Rise)5.0 Amps 125 VAC 0.5 Amps 125 VDC 27120-22Stainless Steel27121-0Brass Normally Open (Close on Rise) 5.0 Amps 125 VAC 0.5 Amps 125 VDC 2.0 Amps 24 VDC 1.0 Amps 48 VDC 27121-20Stainless Steel 28020-3Stainless SteelNormally Closed (Open on Rise) 5.0 Amps 125 VAC 0.5 Amps 125 VDC 28021-5Stainless SteelNormally Open (Close on Rise)5.0 Amps 125 VAC 0.5 Amps 125 VDC 2.0 Amps 24 VDC 1.0 Amps 48 VDC12-200001-00X*Approximate weight: 5 oz. All shell material is Stainless Steel. All Stainless Steel is Type 300.* Specialty product with limited availability.Figure 4. Hexagonal HeadFigure 5. Coupling HeadHORIZONTAL DETECT-A-FIRE DETECTOR SPECIFICATIONSHorizontal D-A-F detectors are designed for locations where appearance is a factor. The low-profile, functional design lends physical protection of the detector while making it suitable for commercial, industrial, mercantile public buildings, institutions, and marine applications in non-hazardous locations (those classified as "ordinary" under the National Electric Code). Flush mounted detectors are designed to fit standard 4-inch octagonal electric boxes and surface mounting detec-tors are designed to mount directly on ceilings or on 4-inch electrical junction boxes. Canadian Electrical Codes requires mounting only to an electrical junction box.Horizontal ModelsTable 3: Model Numbers: 27021-0, 27021-1, 27020-0*, 27020-1*°F Setting°FToleranceSpacing(in feet)RTI ColorCodingUL ULCFM140+7/-8505020Quick Black 160+7/-8252520Quick Black 190+7/-8505025Fast White 210+7/-8255025Fast White 225+7/-8255025Fast White 275±10255025Fast Blue 325±10505025Fast Red •*27020-0 and 27020-1 are normally closed devices and do not meet the requirements of NFPA-72 for use as initiating devices (they are 2-wire devices).•*27020-0 and 27020-1 are not listed by FM with RTI.Table 4: Horizontal D-A-F SpecificationsModel Number Mounting Style Contact Operation Electrical Rating (Resistive Only)27020-0Flush Mount Normally Closed(Open on Rise)5.0 Amps 125 VAC 0.5 Amps 125 VDC27020-1Surface Mount27021-0Flush MountNormally Open(Close on Rise)5.0 Amps 125 VAC 0.5 Amps 125 VDC 2.0 Amps 24 VDC 1.0 Amps 48 VDC27021-1Surface Mount Approximate weight: 10 oz.Figure 6. Horizontal Detectors Note:Horizontal D-A-F detectors areequipped with connector blocks inplace of leadwires.HORIZONTAL AND VERTICAL DETECT-A-FIREDETECT-A-FIRE MOUNTINGD-A-F detectors are not position sensitive. Horizontal and vertical detectors refer to the most common mounting configuration for that detector. However each type can be mounted either horizontally or vertically depending on the application and installation requirements.Table 5: D-A-F Response Time Index (RTI) and SpacingModel No.Configuration Contact OperationTemperature (Set Point)Response Time Index (ft-s) 1/2RTIClassificationRTI Rated SpacingOld Rated Spacing27021-0Horizontal Flush Mount Normally Open (Close on Rise)140°F (60°C), 160°F (71°C)110Quick(20 X 20) ft (6 x 6) m (25 x 25) ft (8 x 8) m27021-1Horizontal Surface Mount 27021-0Horizontal Flush Mount Normally Open (Close on Rise)190°F (88°C), 210°F (99°C), 225°F (107°C), 275°F (135°C), 325°F (163°C)140Fast(25 x 25) ft (8 x 8) m (25 x 25) ft (8 x 8) m27021-1Horizontal Surface Mount 27121-0Vertical Brass Head Normally Open (Close on Rise)140°F (60°C), 160°F (71°C), 190°F (88°C), 210°F (99°C), 225°F (107°C), 275°F (135°C), 325°F (163°C), 360°F (182°C), 450°F (232°C), 500°F (260°C), 600°F (316°C), 725°F (385°C)99 (140°F, 160°F)148 (190°F, 210°F,225°F, 275°F, 325°F,360°F, 450°F, 500°F,600°F, 725°F)V-Fast(30 X 30) ft (9 x 9) m (25 x 25) ft (8 x 8) m27121-20Vertical Stainless Head 28021-5Vertical Stainless Coupling HeadNote: Spaces shown are distances between detectors on smooth ceilings, the distances from partitions or walls would be half that shown. Authority Having Jurisdiction (AHJ) should be consulted before installation.Table 6: Mounting and Hazardous Location ClassModel Number (Vertical Only)Hazardous LocationFitting required for UL & ULC Listing and FM Approval27120-227121-2028020-328021-5Class I, Groups A, B, C and D;Class II, Groups E, F and G Mount detector to a suitable listed fitting in accordance with National Electric Code and/or local authority having jurisdiction.27120-027121-0Class I, Groups B, C and D;Class II Groups E, F and GTable 7: Derating Factor for Ceiling HeightHeat Detector Spacing Reduction Based on Ceiling Height Ceiling Height AboveUp to and Including Derating Factormftmft00 3.0510 1.003.0510 3.66120.913.6612 4.27140.844.2714 4.88160.774.8816 5.49180.715.4918 6.10200.646.1020 6.71220.586.71227.32240.527.32247.93260.467.93268.54280.408.54289.14300.34This table outlines the derating factor required depending on ceiling height based on NFPA 72 guidelines for D-A-F detector installation .CONSTRUCTION•Stainless steel shell sensing element. Cold rolled steel mounting facility. Off-White finish.•#18 AWG Teflon™ insulated wire is used on detectors exposed to temperatures up to 375°F.•#16 AWG TGGT insulated wire is used on detectors exposed to temperatures above 375°F.MODIFICATIONS•Add ULC Label to any temperature setting.•Add fluorocarbon coating for better corrosion resistance on select models and temperatures.(Models 27120-22, 27121-20, 28020- 3, 28021-5. Maximum temperature is 500°F.)TEMPERATURE SETTING SELECTIONTo avoid nuisance activations, Fenwal Controls strongly suggests selecting a D-A-F detector with a temperature setting a minimum of 100°F above the maximum ambient expected temperature.DEVICE SELECTIONThe table below shows three categories of fire detection devices and their relative response levels for reaction to three different rate-of-rise conditions. Consult the AHJ for specific applications.AGENCY LISTINGSD-A-F detectors are UL and ULC listed and FM Approved as fire detection thermostats (close on temperature rise) and as releasing devices (open on temperature rise).NOTES:•D-A-F detectors are temperature preset at the factory.•For corrosive environments, care should be taken to protect the D-A-F detector to obtain optimal performance and maximum life. Consult factory for fluorocarbon coating option.•For field wiring requirements to connect to D-A-F, please refer to the installation instructions.•Per UL521 requirements - low temperature exposure test is - 22°F (-30°C)•D-A-F detectors are designed for long life expectancy, however due to various field conditions it is required that the detectors be tested annually per NFPA guidelines or local fire codes.•Replace D-A-F detector after any fire or heat related event, any mechanical damage, or after 10 years of continuous service.•D-A-F detectors are CE Listed. The product family has been evaluated in accordance with IEC 60947-1 and IEC 60947-5- 1, and is documented under Intertek Report No. 102294754 BOX-001 as an overheat detector. Detectors have a rated insulation and impulse voltages of 1500 VAC.Table 8: Device Selection for Rate-of-Rise ConditionsDevice Type Under 10°/MinBetween 10-40°/MinOver 40°/MinD-A-F Rate CompensatedDetector First First Secondbut at selected protection levelFixed Temp. Detector Second Second ThirdRate-of-Rise DetectorThirdThirdFirstbut may be a false alarmTable 9: Agency ApprovalsAgency File NumberLocation UL S492Ordinary UL E19310HazardousULC CS341-E Ordinary and HazardousFM J.I. OV2HO.AEHazardous FM 17302OrdinaryUL S2410Ordinary (600 & 725°F)UL E89599Hazardous (600 & 725°F)CEIEC 60947-5-1—DETECT-A-FIRE is a registered trademark of Kidde-Fenwal, Inc., or its parents, subsidiaries, or affiliates.All other trademarks are the property of their respective owners.EXPORT INFORMATION (USA)Jurisdiction: EAR Classification: EAR99This document contains technical data subject to the EAR.DETECT-A-FIRE MOUNTING BOXFigure 7. Optional Explosion Proof Mounting Box,P/N 06-116317-001Note:Figure 7. Complies with NEC Class I, Groups A, B, C, D, Class II, Group E,F,G, Class III, and U.L. Standard 1203. Explosion Proof Mounting Box must be purchased separately from the D-A-F.HOW TO ORDER1.Select a D-A-F model from specifications in Table 2 for vertical design or Table 4 for horizontal design.2.Select a temperature rating from Table 1 for vertical design or Table 3 for horizontal design.Note:To avoid nuisance activations, Fenwal Controls strongly suggests selecting a D-A-F detector with a temperature setting a minimum of 100°F above the maximum ambient expected temperature.3.Optional, select item from the Modifications Section. Consult Fenwal Controls to ensure modifications are available on the selected model.Example: 27121-20 at 190°F with Fluorocarbon CoatingTable 10: Stocked Models and Temperature SettingsModelStyleContact TypeTemperature Setting (°F)14016019021022527532536045050060072527020-0Horizontal, Flush Mount NC S SN/A N/A N/A N/A N/A 27020-1Horizontal, Surface Mount NC S N/A N/A N/A N/A N/A 27021-0Horizontal, Flush Mount NO S S SN/A N/A N/A N/A N/A 27021-1Horizontal, Surface Mount NO S S N/AN/AN/AN/A N/A 27120-0Vertical, Brass Head NC S SS S SN/A N/A 27120-22Vertical, Stainless Steel NC S S S N/A N/A 27121-0Vertical, Brass Head NO S S S SS S S SS SS S27121-20Vertical, Stainless Steel NO SS S S SS S S28021-5Vertical, Coupling HeadNOSSSS SS - Stocked D-A-F detector available for quick delivery of a limited quantity. Other models and variations are manufactured on demand.。
Rev. 0.1 1/14Copyright © 2014 by Silicon LaboratoriesSi1146 Slider Demo KitLIDER EMO IT SER S UIDE1. Kit ContentsThe Si1146 UVIRSlider2EK Demo Kit contains the following items:⏹ Si1146 UVIRSlider2EK Demo Board⏹ Si1146 UVIRSlider2EK Demo Board Quick Start Guide ⏹ USB Cable2. IntroductionThe Si1146 UVIRSlider2EK Demo Board implements a UV Index sensing function based around the Si1146 UV,infrared proximity, and ambient light sensor. As shown in Figure 1, the main components of the board are the Si1146 sensor (U2), the C8051F800 microcontroller (U1), and two infrared emitters (DS1 and DS2). Hardware is also provided on-board to facilitate code development for the C8051F800 and communications with software support packages over a USB interface.The firmware running on the C8051F800 measures the UV Index of the environment of the Si1146 sensor and indicates the UV Index detected by illuminating blue LEDs on the board.The board is also capable of running a touchless slider algorithm using infrared light energy detected by the Si1146, while each of the two infrared emitters are independently activated. The infrared light from these emitters will be reflected back to the Si1146 by any object that is placed in proximity to the board. The left-right position is then calculated from these two measurements and used to illuminate the appropriate signal LED. If no object is close enough to the board, the measured signal levels will fall below pre-determined thresholds, and no signal LEDs will be illuminated. In addition to indicating the current position, the algorithm is also able to detect different gestures from the infrared sensor, as described in Table 2 on page 3. The Si1146’s ambient light sensor (ALS) is also monitored by the software, which can determine the amount of ambient light present.On this board, the infrared emitters used are OSRAM part number SFH 4056. These emitters have a power rating of 40mW and a half-angle of 22 degrees. Other emitters with different characteristics may also be used,depending on the specific application requirements.Note:The touchless infrared position and gesture detection implemented in the example is patent pending.Figure 1.Si1146 Slider Demo BoardD S 13DS101D S 12D S 8D S 9D S 14D S 5D S 6D S 7D S 4D S 3+5V_INSi1146 UVIRSlider2EK3. Running the Pre-Loaded UV Index Demo1. The board receives its power over the USB interface. Connect one end of a USB cable to the USBconnector (P1) on the Si1146 UVIRSlider2EK Demo Board and the other end to a USB Port on the PC.2. The red LED DS10 should light on the board, indicating that it is receiving power from the USB.3. The visible blue LEDs will light according to the perceived UV Index of the environment above the board.4. See Table1 on page2 for a description on what the number of blue LEDs illuminated represents.Table 1. UV Demo Output Description# Blue LEDs UV Index UV Index Description 511+Extreme48,9,10Very High36,7High23,4,5Moderate11,2Low4. Software OverviewThe UVIRSLIDER2EK is supported by the UVIRSLIDER Demo application and the Si114x Programmer's Toolkit. The software can be downloaded from the web at: /products/sensors/pages/optical-sensor-software.aspx.The UVIRSLIDER Demo application allows the user to select the UV Index demo, the IR Slider demo, or the UDP Waveform Viewer. The UV demo includes a GUI that displays the full 11+ UV Index value and the 5-level UV sun exposure level. The IRSLIDER demo tracks the movement of your hand over the 8 LEDs on the board using proximity to sense the position of your hand. The IRSLIDER demo can also detect hand gestures as described in Table2. The UDP Waveform Viewer allows users to see real-time infrared proximity and ambient light measurements from the Si1146 in a graphical form.The Si114x Programmer's Toolkit API enables rapid development of Si114x software in a PC environment using the Si1146 UVIRSlider2EK Demo Board. The Si114x Programmer's Toolkit contains example source code that allows developers to get started quickly and then tailor the code to their needs. In addition, the Silicon Labs Integrated Development Environment (IDE) provides a means of developing code for the C8051F800 and uses the USB connection on the board to program the MCU and perform in-system debugging.4.1. Using the UVIRSlider2EK Demo Board with the UVIRSLIDER Demo ApplicationThe Si1146 UVIRSlider2EK Demo Board is supported by the UVIRSLIDER Demo application. The UVIRSLIDER Demo application allows the user to run the UV Index or IR Slider demos as well as launch the UDP Waveform Viewer. The UDP Waveform Viewer allows users to see real-time infrared proximity and ambient light measurements from the Si1146 in a graphical form. The communications interface to the Si1146 UVIRSlider2EK Demo Board is provided over the USB connection.To use the UVIRSLIDER Demo with the Si1146 UVIRSlider2EK Demo Board, perform the following steps:1. Connect the Si1146 UVIRSlider2EK Demo Board to the PC using a USB cable.2. Launch the UVIRSLIDER Demo. If you do not have this application, it is available for download at the linkprovided at the beginning of the Software Overview description.3. Select the desired demo from the Main Menu.4. Follow the instructions on the screen for how to use the demo.5. The UDP Waveform Viewer can be launched by pressing on the Waveform Viewer button on the demoSi1146 UVIRSlider2EKapplication screen.Note:The the Si114x Programmer’s Toolkit, and the IDE cannot connect to the Si1146 UVIRSlider2EK Demo Board at the same time. Be certain to disconnect from the board in one software package before trying to connect in the other. Figure2 shows an example of the UVIRSLIDER Demo main window when connected to the Si1146 UVIRSlider2EK Demo Board.Figure2.UVIRSLIDER Demo Main WindowTable 2. Recognized GesturesGesture Name Parameter Description of Action and LED Indication Pause Position Hold hand steady in a single position above the board for 3/4 sec-ond. Current position LED will blink briefly.Swipe Left Speed Move hand rapidly from the right side to the left side of the board.LEDs will briefly indicate a leftward sweep pattern.Swipe Right Speed Move hand rapidly from the left side to the right side of the board.LEDs will briefly indicate a rightward sweep pattern.Si1146 UVIRSlider2EK4.2. Si114x Programmer’s Toolkit4.2.1. Software APIThe Si114x Programmer’s Toolkit API enables rapid development of Si114x software in a PC environment using the Si1146 UVIRSlider2EK Demo Board. By emulating an I2C interface over USB, the Si114x Programmer’s Toolkit API allows source code to be developed on a PC and then migrated quickly and easily to an MCU environment once target hardware is available. Either commercially-available or free PC-based C compilers can be used for software development with the Si114x Programmer’s Toolkit API.The Si114x Programmer’s Toolkit API also includes the Si114x UDP Waveform Viewer Application. This tool runs in conjunction with user applications to display and debug the measurements taken from the Si1146 UVIRSlider2EK Demo Board.Note:The Si114x Programmer’s Toolkit and IDE cannot connect to the Si1146 UVIRSlider2EK Demo Board at the same time.Be certain to disconnect from the board in one software package before trying to connect in the other.4.2.2. Command Line UtilitiesFor evaluation of the Si1146 UVIRSlider2EK Demo Board without the need to develop and compile source code, a flexible set of command line utilities is also provided with the Si114x Programmer’s Toolkit. These utilities can be used to configure and read samples from the Si1146 UVIRSlider2EK Demo Board. For automated configuration and scripting, the command line utilities can be embedded into .bat files.4.2.3. Sample Source CodeFor faster application development, the Si114x Programmer’s Toolkit contains example source code for the Si1146 UVIRSlider2EK Demo Board and for each of the command line utilities. Developers can get started quickly by using the Si114x example source code and then tailoring it to their needs.4.2.4. Downloading the Si114x Programmer’s ToolkitThe Si114x Programmer’s Toolkit and associated documentation is available from the web at the following URL: /products/sensors/pages/optical-sensor-software.aspx.Si1146 UVIRSlider2EK4.3. Silicon Laboratories IDEThe Silicon Laboratories IDE integrates a source-code editor, a source-level debugger, and an in-system Flash programmer. This tool can be used to develop and debug code for the C8051F800 MCU, which is included on the Si1146 UVIRSlider2EK Demo Board. The use of several third-party compilers and assemblers is supported by the IDE.4.3.1. IDE System RequirementsThe Silicon Laboratories IDE requirements are:⏹ Pentium-class host PC running Microsoft Windows 2000 or newer⏹ One available USB port4.3.2. Third Party ToolsetsThe Silicon Laboratories IDE has native support for many 8051 compilers. The full list of natively-supported tools is as follows:⏹ Keil⏹ IAR⏹ Raisonance⏹ Tasking⏹ SDCC4.3.3. Downloading the Example Firmware ImageSource code that has been developed and compiled for the C8051F800 MCU on the Si1146 UVIRSlider2EK Demo Board may be downloaded to the board using the Silicon Laboratories IDE. Follow the instructions below to update or refresh the .HEX image in the Si1146 UVIRSlider2EK Demo Board.1. Connect the Si1146 UVIRSlider2EK Demo Board to the PC using a USB cable.2. Launch the Silicon Labs IDE, and click on Options->Connection Options.3. Select “USB Debug Adapter”, and then select the board from the list (it should show up as “TS” followed bya serial number).4. Select “C2” as the debug interface, and press “OK”.5. Connect to the board by pressing the “Connect” icon, or using the keyboard shortcut Alt+C.6. Click on the “Download” icon, or use the keyboard shortcut Alt+D.7. In the download dialog window, click “Browse”.8. Change to Files of Type “Intel Hex (*.hex)” and then browse to select the file.9. Click “Open” then “Download”.10. To run the new image, either press “Run” or “Disconnect” in the IDE.Note:The Si114x Programmer’s Toolkit and the IDE cannot connect to the Si1146 UVIRSlider2EK Demo Board at the same time. Be certain to disconnect from the board in one software package before trying to connect in the other.Si1146 UVIRSlider2EK5. SchematicF i g u r e 3.S i 1146 U V I R S l i d e r 2E K D e m o B o a r d S c h e m a t i cSilicon Laboratories Inc.400 West Cesar Chavez Austin, TX 78701USASmart.Connected.Energy-Friendly .Products/productsQuality/qualitySupport and CommunityDisclaimerSilicon Laboratories intends to provide customers with the latest, accurate, and in-depth documentation of all peripherals and modules available for system and software implementers using or intending to use the Silicon Laboratories products. Characterization data, available modules and peripherals, memory sizes and memory addresses refer to each specific device, and "Typical" parameters provided can and do vary in different applications. Application examples described herein are for illustrative purposes only. Silicon Laboratories reserves the right to make changes without further notice and limitation to product information, specifications, and descriptions herein, and does not give warranties as to the accuracy or completeness of the included information. Silicon Laboratories shall have no liability for the consequences of use of the information supplied herein. This document does not imply or express copyright licenses granted hereunder to design or fabricate any integrated circuits. The products are not designed or authorized to be used within any Life Support System without the specific written consent of Silicon Laboratories. A "Life Support System" is any product or system intended to support or sustain life and/or health, which, if it fails, can be reasonably expected to result in significant personal injury or death. Silicon Laboratories products are not designed or authorized for military applications. Silicon Laboratories products shall under no circumstances be used in weapons of mass destruction including (but not limited to) nuclear, biological or chemical weapons, or missiles capable of delivering such weapons.Trademark InformationSilicon Laboratories Inc.® , Silicon Laboratories®, Silicon Labs®, SiLabs® and the Silicon Labs logo®, Bluegiga®, Bluegiga Logo®, Clockbuilder®, CMEMS®, DSPLL®, EFM®, EFM32®, EFR, Ember®, Energy Micro, Energy Micro logo and combinations thereof, "the world’s most energy friendly microcontrollers", Ember®, EZLink®, EZRadio®, EZRadioPRO®, Gecko®, ISOmodem®, Precision32®, ProSLIC®, Simplicity Studio®, SiPHY®, Telegesis, the Telegesis Logo®, USBXpress® and others are trademarks or registered trademarks of Silicon Laborato-ries Inc. ARM, CORTEX, Cortex-M3 and THUMB are trademarks or registered trademarks of ARM Holdings. Keil is a registered trademark of ARM Limited. All other products or brand names mentioned herein are trademarks of their respective holders.。
MUST ANG MACH-EALL -ELECTRIC 2021 FORD 2021M UST ANG M ACH-E.The iconic Ford Mustang® DNA is undeniable in this 100% SOUL-STIRRING SUV. The sloping fastback and sequential® is easy to recognize. Yet, it’s like no other. SPACE FOR 5 and SUV versatility are just a few of the unique traits of this ALL-ELECTRIC PONY.PREMIUM. Black Onyx Perforated ActiveX ™ Seating Material. Heated steering wheel. B&O ™ Sound System by Bang & Olufsen. Available equipment.1. FordPass App, compatible with select smartphone platforms, is available via download. Message and data rates may apply. 2. Requires feature activation.Beautifully streamlined. SEAMLESSLY CONNECTED. Ford Mustang Mach‑E ® comes with an Intelligent Access key fob, and the FordPass ™ App 1 unlocks Phone As A Key access.2 Either way, your Mustang Mach‑E recognizes you as soon as you get close. Just press the illuminated E‑Latch button to release your door. Step into your cabin personalized by up to 3 driver profiles, and experience THE FUTURE OF EXHIL ARATION.MIND-BLOWING TORQUE. A THRILL TODRIVE.Ford Mustang Mach‑E ® delivers a driving experience like no other Mustang ® before it. We’re talking unbridled exhilaration on every drive – courtesy of the all‑electric drivetrain with its breathtaking power and torque. It takes less than half a second 1 for the motors that put Mustang Mach‑E in motion to reach maximum torque. Electric power‑assisted steering and a fully independent suspension designed for lightweight strength work together to channel this thrilling output to the road.1. Horsepower and torque are independent attributes and may not be achieved simultaneously. Based on manufacturer calculation using computer engineering simulations. Y our results may vary.E X H I L A R AT I O N. E L E C T R IF I E D.MORE POWER to the pavement. The available electric All‑Wheel‑Drive (eAWD) System can use the full power of both the front and rear axles to provide impressiveacceleration and handling.This all‑new SUV inspires ALL-SEASON ADVENTURES. The available eAWD System’s front and rearelectric motors can distribute power as needed to help you take on challenging road conditions. To help keepthe JOY IN EVERY RIDE, you’ll find both traction control and AdvanceTrac® electronic stability control1standard on every Ford Mustang Mach‑E.®IIMI N DR E A D E Enjoy wireless compatibility with Apple CarPlay® and Android Auto.™4Keep it clean and simple with the wireless charging pad.5Get heart-pounding bass with the available B&O™ Sound System. Watch quick how-to videos in the digital owner’s manual while in Park.31. Cargo and load capacity limited by weight and weight distribution.Share the exhilaration of Ford Mustang Mach‑E ® with YOUR PART Y OF 5. This spacious, well‑appointed SUV interior features PLENT Y OF HEAD AND LEG ROOM and storage spots for your road‑trip essentials. Split‑fold‑flat rear seats make way for cargo inside,1 while the water‑resistant front trunk offers washable, drainable FRONT-LOAD FREEDOM.PREMIUM. Rapid Red Metallic Tinted Clearcoat.4 Available equipment.1. Driver-assist features are supplemental and do not replace the driver’s attention, judgment and need to control the vehicle. It does not replace safe driving. See owner’s manual for details and limitations.2. Navigation services require SYNC 4 and FordPass Connect, complimentary Connected Service and the FordPass App (see FordPass Terms for details). Eligible vehicles receive a complimentary 3-year trial of navigation services that begins on the new vehicle warranty start date. Customers must unlock the navigation service trial by activating the eligible vehicle with a FordPass member account. If not subscribed by the end of the complimentary period, the connected navigation service will terminate, and the system will revert to embedded offline navigation. Connected service and features depend on compatible AT&T network availability. Evolving technology/cellular networks/vehicle capability may limit functionality and prevent operation of connected features. FordPass App, compatible with select smartphone platforms, is available via download. Message and data rates may apply.3. Active Drive Assist is a hands-free highway driving feature. T he Active Drive Assist Prep Kit contains the hardware required for this feature. Software for the feature will be available for purchase at a later date. Active Drive Assist functionality expected 3rd quarter 2021CY. Separate payment for feature software required to activate full functionality at that time.4. Additional charge.From the driveway to the highway, we can all use more confidence at the wheel. That’s why Ford Mustang Mach‑E® offers an extensive, well‑rounded collection of standard and available DRIVER-ASSIST TECHNOLOGIES. These advanced features are about supplementing your driving skills.1 Helping you feel confidently in command.FORD CO-PILOT360 ASSIST 2.0Stop‑and‑go traffic, unexpected challenges, and finding yourway with ease. Get help from these standard features:INTELLIGENT ADAPTIVE CRUISE CONTROLINTERSECTION ASSISTEVASIVE STEERING ASSISTCONNECTED BUILT-IN NAVIGATION SYSTEM2AVAILABLE TECHNOLOGYParallel and perpendicular parking, crowded lots and more.Drive confidently with help from these available features:360-DEGREE CAMERAWITH SPLIT VIEW AND FRONT WASHERF ORD CO-PILOT360 ACTIVE 2.0 PREP PACKAGE– ACTIVE PARK ASSIST™ 2.0– ACTIVE DRIVE ASSIST™ PREP KIT3FORD CO-PILOT360 2.0Changing lanes, backing up and more. These smart standards are designedto help make driving enjoyable:P RE-COLLISION ASSISTWITH AUTOMATICEMERGENCY BRAKINGB LIS® (BLIND SPOTINFORMATION SYSTEM)WITH CROSS-TRAFFIC ALERTLANE-KEEPING SYSTEMREVERSE SENSING SYSTEMREVERSE BRAKE ASSISTPOST-COLLISION BRAKINGAUTO HIGH-BEAM HEADLAMPSREAR VIEW CAMERAC O MMA ND T HE R O A D WITH FORD CO-PILOT360™ TECHNOLOGY.Fictional charger location. Visit for a full list of Electrify America charging stations.1.Range and charge time based on manufacturer computer engineering calculations. The charging rate decreases as battery reaches full capacity. Your results may vary based on peak charging times and battery state of charge. Actual range varies with conditions such as external environment, vehicle use, driving behaviors, vehicle maintenance, lithium-ion battery age and state of health.2.FordPass Connect, the FordPass App, and complimentary Connected Service are required for remote features (see FordPass Terms for details). Connected service and features depend on compatible AT&T network availability. Evolving technology/cellular networks/vehicle capability may limit functionality and prevent operation of connected features. Connected service excludes Wi-Fi hotspot. FordPass App, compatible with select smartphone platforms, is available via download. Message and data rates may apply.plimentary 2-year trial begins with EV New Vehicle Limited Warranty start date. Customer will be billed for all fees including but not limited to fees for use of charging stations and any related parking fees.4.Based on original equipment manufacturers (OEM)/automotive manufacturers that sell all-electric vehicles and have publicly announced charging networks. Department of Energy data used.FA R E W E L L TO GAS STATIONS AND VEHICLE EMISSIONS.y at-home charging options. Convenient charging across Nor th America.1Space White Metallic 5Iconic Silver MetallicLight Space Gray Perforated ActiveX Seating Material / 1–82 Star White Metallic Tri-coat110 Cyber Orange Metallic Tri-coat1,2E X TERIORINTERIORA CBDEA. Available Panoramic Fixed-Glass Roof filters UV rays and comes with an Infrared Reflective (IRR) windshield that helps the interior stay cooler in the summer and warmer in the winter.B. Available hands-free, foot-activated liftgate features an adjustable opening height.C. 10.2" digital cluster screen displays your preferred information.D. Combo connector system charge port with LED charge status indicator can charge the vehicle’s battery on 120V, 240V or Direct Current (DC Fast Charge).E. Heated sideview mirrors feature Pony projection lamps.Colors are representative only. See your dealer for actual paint/trim options. 1. Additional charge. 2. Available late summer 2021.SELECTCALIFORNIA ROUTE 1PREMIUMFIRST EDITIONGT 2GT PERFORMANCE EDITION 2The Ford Connected Charge Station provides fast charging at home for your Mustang Mach-E.® It can be installed near where you park, inside or out. You can remotelylock and unlock it, get insights on your charge sessions, and track your energy usage, all from the FordPass™ App, or even with Amazon Alexa and Google Assistant.™Cargo area protector Charge cord bag1Floor liners Dash cam systems1S H O P T H E C O M P L E T E C O L L E C T IO N|/suvs/mach-eThe 2021 Ford Mustang Mach-E qualifies for the $7,500 Federal Tax Credit.Depending on where you live and your personal tax situation, you may qualify for federal, state and city tax incentivesfor driving an all-electric vehicle. You can learn more at /laws/409.1. Ford Licensed Accessory.2.Based on full charge. Actual range varies with conditions such as external environment, vehicle use, driving behaviors, vehicle maintenance, lithium-ionbattery age and state of health.3. Final EPA-estimated ratings available in the 2021CY. 4. The Federal Tax Credit is a potential future tax savings. The amount of your tax savings will dependon your individual tax circumstances. Please consult with your own tax or legal professional to determine eligibility, and specific amount of incentives or rebates available. Incentives andadditional rebates are not within the control of Ford. This information does not constitute tax or legal advice.Ford and Oval Device, 911 Assist, Active Drive Assist, Active Park Assist, ActiveX, AdvanceTrac, AppLink, BLIS, Ford Co-Pilot360, Ford Credit, Ford Protect, FordPass, Mustang, MustangMach-E, MyKey, Personal Safety System, SYNC and related marks are trademarks of Ford Motor Company.Amazon, Alexa, and all related logos and marks are trademarks of , Inc. or its affiliates. Apple and Apple CarPlay are trademarks of Apple Inc., registered in the U.S. and othercountries. BANG & OLUFSEN and B&O are registered trademarks of Bang & Olufsen Group. Licensed by Harman Becker Automotive Systems Manufacturing Kft. All rights reserved. Brembois a trademark of Brembo S.p.A. Google, Android, Android Auto, Google Assistant and Google Play are trademarks of Google LLC. Requires the Android Auto app on Google Play and anAndroid compatible smartphone running Android™ 5.0 or higher. MagneRide is a trademarked technology of BWI Group. Miko is a registered trademark of Miko s.r.l. Pirelli is a trademarkof Pirelli & C. S.p.A. SiriusXM and all related marks and logos are trademarks of Sirius XM Radio Inc.Comparisons based on competitive models (class is Medium Utility), publicly available information and Ford certification data at time of release. Vehicles may be shown with optional equipment.Features may be offered only in combination with other options or subject to additional ordering requirements/limitations. Dimensions shown may vary due to optional features and/orproduction variability. Information is provided on an “as is” basis and could include technical, typographical or other errors. Ford makes no warranties, representations, or guarantees of anykind, express or implied, including but not limited to, accuracy, currency, or completeness, the operation of the information, materials, content, availability, and products. Ford reserves theright to change product specifications, pricing and equipment at any time without incurring obligations. Your Ford Dealer is the best source of the most up-to-date information on Ford vehicles.POTENTIALTAX INCENTIVES421MACPDF R1From day one, we had the future in mind. And over 117 years later, we continue that tradition ofinnovation with Ford Co-Pilot360,™ helping push us closer to autonomous driving, the latest generationof SYNC ® to communicate untethered and our first fully electric vehicle – Ford Mustang Mach-E.®We’ve always had our mind on the future. Now we’re building it.©2020Ford M otor Company 1231.2. 3.。
(完整版)S C I写作句型汇总-CAL-FENGHAI-(2020YEAR-YICAI)_JINGBIANSCI论文写作中一些常用的句型总结(一)很多文献已经讨论过了一、在Introduction里面经常会使用到的一个句子:很多文献已经讨论过了。
它的可能的说法有很多很多,这里列举几种我很久以前搜集的:A.??Solar energy conversion by photoelectrochemical cells?has been intensively investigated.?(Nature 1991, 353, 737 - 740?)B.?This was demonstrated in a number of studies that?showed that composite plasmonic-metal/semiconductor photocatalysts achieved significantly higher rates in various photocatalytic reactions compared with their pure semiconductor counterparts.C.?Several excellent reviews describing?these applications are available, and we do not discuss these topicsD.?Much work so far has focused on?wide band gap semiconductors for water splitting for the sake of chemical stability.(DOI:10.1038/NMAT3151)E.?Recent developments of?Lewis acids and water-soluble organometallic catalysts?have attracted much attention.(Chem. Rev. 2002, 102, 3641?3666)F.?An interesting approach?in the use of zeolite as a water-tolerant solid acid?was described by?Ogawa et al(Chem.Rev. 2002, 102, 3641?3666)G.?Considerable research efforts have been devoted to?the direct transition metal-catalyzed conversion of aryl halides toaryl nitriles. (J. Org. Chem. 2000, 65, 7984-7989)H.?There are many excellent reviews in the literature dealing with the basic concepts of?the photocatalytic processand the reader is referred in particular to those by Hoffmann and coworkers,Mills and coworkers, and Kamat.(Metal oxide catalysis,19,P755)I. Nishimiya and Tsutsumi?have reported on(proposed)the influence of the Si/Al ratio of various zeolites on the acid strength, which were estimated by calorimetry using ammonia. (Chem.Rev. 2002, 102, 3641?3666)二、在results and discussion中经常会用到的:如图所示A. GIXRD patterns in?Figure 1A show?the bulk structural information on as-deposited films.?B.?As shown in Figure 7B,?the steady-state current density decreases after cycling between 0.35 and 0.7 V, which is probably due to the dissolution of FeOx.?C.?As can be seen from?parts a and b of Figure 7, the reaction cycles start with the thermodynamically most favorable VOx structures(J. Phys. Chem. C 2014, 118, 24950?24958)这与XX能够相互印证:A.?This is supported by?the appearance in the Ni-doped compounds of an ultraviolet–visible absorption band at 420–520nm (see Fig. 3 inset), corresponding to an energy range of about 2.9 to2.3 eV.B. ?This?is consistent with the observation from?SEM–EDS. (Z.Zou et al. / Chemical Physics Letters 332 (2000) 271–277)C.?This indicates a good agreement between?the observed and calculated intensities in monoclinic with space groupP2/c when the O atoms are included in the model.D. The results?are in good consistent with?the observed photocatalytic activity...E. Identical conclusions were obtained in studies?where the SPR intensity and wavelength were modulated by manipulating the composition, shape,or size of plasmonic nanostructures.?F.??It was also found that areas of persistent divergent surface flow?coincide?with?regions where convection appears to be consistently suppressed even when SSTs are above 27.5°C.(二)1. 值得注意的是...A.?It must also be mentioned that?the recycling of aqueous organic solvent is less desirable than that of pure organic liquid.B.?Another interesting finding is that?zeolites with 10-membered ring pores showed high selectivities (>99%) to cyclohexanol, whereas those with 12-membered ring pores, such as mordenite, produced large amounts of dicyclohexyl ether. (Chem. Rev. 2002, 102, 3641?3666)C.?It should be pointed out that?the nanometer-scale distribution of electrocatalyst centers on the electrode surface is also a predominant factor for high ORR electrocatalytic activity.D.?Notably,?the Ru II and Rh I complexes possessing the same BINAP chirality form antipodal amino acids as the predominant products.?(Angew. Chem. Int. Ed., 2002, 41: 2008–2022)E. Given the multitude of various transformations published,?it is noteworthy that?only very few distinct?activation?methods have been identified.?(Chem. Soc. Rev., 2009,?38, 2178-2189)F.?It is important to highlight that?these two directing effects will lead to different enantiomers of the products even if both the “H-bond-catalyst” and the?catalyst?acting by steric shielding have the same absolute stereochemistry. (Chem. Soc. Rev.,?2009,?38, 2178-2189)G.?It is worthwhile mentioning that?these PPNDs can be very stable for several months without the observations of any floating or precipitated dots, which is attributed to the electrostatic repulsions between the positively charge PPNDs resulting in electrosteric stabilization.(Adv. Mater., 2012, 24: 2037–2041)2.?...仍然是个挑战A.?There is thereby an urgent need but it is still a significant challenge to?rationally design and delicately tail or the electroactive MTMOs for advanced LIBs, ECs, MOBs, and FCs.?(Angew. Chem. Int. Ed.2 014, 53, 1488 – 1504)B.?However, systems that are?sufficiently stable and efficient for practical use?have not yet been realized.C.??It?remains?challenging?to?develop highly active HER catalysts based on materials that are more abundant at lower costs. (J. Am. Chem.Soc.,?2011,?133, ?7296–7299)D.?One of the?great?challenges?in the twenty-first century?is?unquestionably energy storage. (Nature Materials?2005, 4, 366 - 377?)众所周知A.?It is well established (accepted) / It is known to all / It is commonlyknown?that?many characteristics of functional materials, such as composition, crystalline phase, structural and morphological features, and the sur-/interface properties between the electrode and electrolyte, would greatly influence the performance of these unique MTMOs in electrochemical energy storage/conversion applications.(Angew. Chem. Int. Ed.2014,53, 1488 – 1504)B.?It is generally accepted (believed) that?for a-Fe2O3-based sensors the change in resistance is mainly caused by the adsorption and desorption of gases on the surface of the sensor structure. (Adv. Mater. 2005, 17, 582)C.?As we all know,?soybean abounds with carbon,?nitrogen?and oxygen elements owing to the existence of sugar,?proteins?and?lipids. (Chem. Commun., 2012,?48, 9367-9369)D.?There is no denying that?their presence may mediate spin moments to align parallel without acting alone to show d0-FM. (Nanoscale, 2013,?5, 3918-3930)(三)1. 正如下文将提到的...A.?As will be described below(也可以是As we shall see below),?as the Si/Al ratio increases, the surface of the zeolite becomes more hydrophobic and possesses stronger affinity for ethyl acetate and the number of acid sites decreases.(Chem. Rev. 2002, 102, 3641?3666)B. This behavior is to be expected and?will?be?further?discussed?below. (J. Am. Chem. Soc.,?1955,?77, 3701–3707)C.?There are also some small deviations with respect to the flow direction,?whichwe?will?discuss?below.(Science, 2001, 291, 630-633)D.?Below,?we?will?see?what this implies.E.?Complete details of this case?will?be provided at a?later?time.E.?很多论文中,也经常直接用see below来表示,比如:The observation of nanocluster spheres at the ends of the nanowires is suggestive of a VLS growth process (see?below). (Science, 1998, ?279, 208-211)2. 这与XX能够相互印证...A.?This is supported by?the appearance in the Ni-doped compounds of an ultraviolet–visible absorption band at 420–520 nm (see Fig. 3 inset), corresponding to an energy range of about 2.9 to 2.3 eVB.This is consistent with the observation from?SEM–EDS. (Chem. Phys. Lett. 2000, 332, 271–277)C.?Identical conclusions were obtained?in studies where the SPR intensity and wavelength were modulated by manipulating the composition, shape, or size of plasmonic nanostructures.?(Nat. Mater. 2011, DOI: 10.1038/NMAT3151)D. In addition, the shape of the titration curve versus the PPi/1 ratio,?coinciding withthat?obtained by fluorescent titration studies, suggested that both 2:1 and 1:1 host-to-guest complexes are formed. (J. Am. Chem. Soc. 1999, 121, 9463-9464)E.?This unusual luminescence behavior is?in accord with?a recent theoretical prediction; MoS2, an indirect bandgap material in its bulk form, becomes a direct bandgap semiconductor when thinned to a monolayer.?(Nano Lett.,?2010,?10, 1271–1275)3.?我们的研究可能在哪些方面得到应用A.?Our ?ndings suggest that?the use of solar energy for photocatalytic watersplitting?might provide a viable source for?‘clean’ hydrogen fuel, once the catalyticef?ciency of the semiconductor system has been improved by increasing its surface area and suitable modi?cations of the surface sites.B. Along with this green and cost-effective protocol of synthesis,?we expect that?these novel carbon nanodots?have potential applications in?bioimaging andelectrocatalysis.(Chem. Commun., 2012,?48, 9367-9369)C.?This system could potentially be applied as?the gain medium of solid-state organic-based lasers or as a component of high value photovoltaic (PV) materials, where destructive high energy UV radiation would be converted to useful low energy NIR radiation. (Chem. Soc. Rev., 2013,?42, 29-43)D.?Since the use of?graphene?may enhance the photocatalytic properties of TiO2?under UV and visible-light irradiation,?graphene–TiO2?composites?may potentially be usedto?enhance the bactericidal activity.?(Chem. Soc. Rev., 2012,?41, 782-796)E.??It is the first report that CQDs are both amino-functionalized and highly fluorescent,?which suggests their promising applications in?chemical sensing.(Carbon, 2012,?50,?2810–2815)(四)1. 什么东西还尚未发现/系统研究A. However,systems that are sufficiently stable and efficient for practical use?have not yet been realized.B. Nevertheless,for conventional nanostructured MTMOs as mentioned above,?some problematic disadvantages cannot be overlooked.(Angew. Chem. Int. Ed.2014,53, 1488 – 1504)C.?There are relatively few studies devoted to?determination of cmc values for block copolymer micelles. (Macromolecules 1991, 24, 1033-1040)D. This might be the reason why, despite of the great influence of the preparation on the catalytic activity of gold catalysts,?no systematic study concerning?the synthesis conditions?has been published yet.?(Applied Catalysis A: General2002, 226, ?1–13)E.?These possibilities remain to be?explored.F.??Further effort is required to?understand and better control the parameters dominating the particle surface passivation and resulting properties for carbon dots of brighter photoluminescence. (J. Am. Chem. Soc.,?2006,?128?, 7756–7757)2.?由于/因为...A.?Liquid ammonia?is particularly attractive as?an alternative to water?due to?its stability in the presence of strong reducing agents such as alkali metals that are used to access lower oxidation states.B.?The unique nature of?the cyanide ligand?results from?its ability to act both as a σdonor and a π acceptor combined with its negativecharge and ambidentate nature.C.?Qdots are also excellent probes for two-photon confocalmicroscopy?because?they are characterized by a very large absorption cross section?(Science ?2005,?307, 538-544).D.?As a result of?the reductive strategy we used and of the strong bonding between the surface and the aryl groups, low residual currents (similar to those observed at a bareelectrode) were obtained over a large window of potentials, the same as for the unmodified parent GC electrode. (J. Am. Chem. Soc. 1992, 114, 5883-5884)E.?The small Tafel slope of the defect-rich MoS2 ultrathin nanosheets is advantageous for practical?applications,?since?it will lead to a faster increment of HER rate with increasing overpotential.(Adv. Mater., 2013, 25: 5807–5813)F. Fluorescent carbon-based materials have drawn increasing attention in recent years?owing to?exceptional advantages such as high optical absorptivity, chemical stability, biocompatibility, and low toxicity.(Angew. Chem. Int. Ed., 2013, 52: 3953–3957)G.??On the basis of?measurements of the heat of immersion of water on zeolites, Tsutsumi etal. claimed that the surface consists of siloxane bondings and is hydrophobicin the region of low Al content. (Chem. Rev. 2002, 102, 3641?3666)H.?Nanoparticle spatial distributions might have a large significance for catalyst stability,?given that?metal particle growth is a relevant deactivation mechanism for commercial catalysts.?3. ...很重要A.?The inhibition of additional nucleation during growth, in other words, the complete separation?of nucleation and growth,?is?critical(essential, important)?for?the successful synthesis of monodisperse nanocrystals. (Nature Materials?3, 891 - 895 (2004))B.??In the current study,?Cys,?homocysteine?(Hcy) and?glutathione?(GSH) were chosen as model?thiol?compounds since they?play important (significant, vital, critical) roles?in many biological processes and monitoring of these?thiol?compounds?is of great importance for?diagnosis of diseases.(Chem. Commun., 2012,?48, 1147-1149)C.?This is because according to nucleation theory,?what really matters?in addition to the change in temperature ΔT?(or supersaturation) is the cooling rate.(Chem. Soc. Rev., 2014,?43, 2013-2026)(五)1. 相反/不同于A. On the contrary, mononuclear complexes, called single-ion magnets (SIM), have shown hysteresis loops of butterfly/phonon bottleneck type, with negligible coercivity, and therefore with much shorter relaxation times of magnetization. (Angew. Chem. Int. Ed., 2014, 53: 4413–4417)B. In contrast, the Dy compound has significantly larger value of the transversal magnetic moment already in the ground state (ca. 10−1μB), therefore allowing a fast QTM. (Angew. Chem. Int. Ed., 2014, 53: 4413–4417)C. In contrast to the structural similarity of these complexes, their magnetic behavior exhibits strong divergence. (Angew. Chem. Int. Ed., 2014, 53: 4413–4417)D. Contrary to other conducting polymer semiconductors, carbon nitride ischemically and thermally stable and does not rely on complicated device manufacturing. (Nature materials, 2009, 8(1): 76-80.)E. Unlike the spherical particles they are derived from that Rayleigh light-scatter in the blue, these nanoprisms exhibit scattering in the red, which could be useful in developing multicolor diagnostic labels on the basis not only of nanoparticle composition and size but also of shape. (Science 2001, 294, 1901-1903)2. 发现,阐明,报道,证实可供选择的词包括:verify, confirm, elucidate, identify, define, characterize, clarify, establish, ascertain, explain, observe, illuminate, illustrate,demonstrate, show, indicate, exhibit, presented, reveal, display, manifest,suggest, propose, estimate, prove, imply, disclose,report, describe,facilitate the identification of举例:A. These stacks appear as nanorods in the two-dimensional TEM images, but tilting experiments confirm that they are nanoprisms. (Science 2001, 294, 1901-1903)B. Note that TEM shows that about 20% of the nanoprisms are truncated. (Science2001, 294, 1901-1903)C. Therefore, these calculations not only allow us to identify the important features in the spectrum of the nanoprisms but also the subtle relation between particle shape and the frequency of the bands that make up their spectra. (Science 2001, 294, 1901-1903)D. We observed a decrease in intensity of the characteristic surface plasmon band in the ultraviolet-visible (UV-Vis) spectr oscopy for the spherical particles at λmax = 400 nm with a concomitant growth of three new bands of λmax = 335 (weak), 470 (medium), and 670 nm (strong), respectively. (Science 2001, 294, 1901-1903)E. In this article, we present data demonstrating that opiate and nonopiate analgesia systems can be selectively activated by different environmental manipulationsand describe the neural circuitry involved. (Science 1982, 216, 1185-1192)F. This suggests that the cobalt in CoP has a partial positive charge (δ+), while the phosphorus has a partial negative charge (δ−), implying a transfer of electron density from Co to P. (Angew. Chem., 2014, 126: 6828–6832)3. 如何指出当前研究的不足A. Although these inorganic substructures can exhibit a high density of functional groups, such as bridging OH groups, and the substructures contribute significantly to the adsorption properties of the material,surprisingly little attention has been devoted to the post-synthetic functionalization of the inorganic units within MOFs. (Chem. Eur. J., 2013, 19: 5533–5536.)B. Little is known, however, about the microstructure of this material. (Nature Materials 2013,12, 554–561)C. So far, very little information is available, and only in the absorber film, not in the whole operational devices. (Nano Lett., 2014, 14 (2), pp 888–893)D. In fact it should be noted that very little optimisation work has been carried out on these devices. (Chem. Commun., 2013, 49, 7893-7895)E. By far the most architectures have been prepared using a solution processed perovskite material, yet a few examples have been reported that have used an evaporated perovskite layer. (Adv. Mater., 2014, 27: 1837–1841.)F. Water balance issues have been effectively addressed in PEMFC technology through a large body of work encompassing imaging, detailed water content and water balance measurements, materials optimization and modeling, but very few of these activities have been undertaken for anion exchange membrane fuel cells, primarily due to limited materials availability and device lifetime. (J. Polym. Sci. Part B: Polym. Phys., 2013, 51: 1727–1735)G. However, none of these studies tested for Th17 memory, a recently identified T cell that specializes in controlling extracellular bacterial infections at mucosal surfaces. (PNAS, 2013, 111, 787–792)H. However, uncertainty still remains as to the mechanism by which Li salt addition results in an extension of the cathodic reduction limit. (Energy Environ. Sci., 2014, 7, 232-250)I. There have been a number of high profile cases where failure to identify the most stable crystal form of a drug has led to severe formulation problems in manufacture. (Chem. Soc. Rev., 2014, 43, 2080-2088)J. However, these measurements systematically underestimate the amount of ordered material. ( Nature Materials 2013, 12, 1038–1044)(六)1. 取决于a. This is an important distinction, as the overall activity of a catalyst will depend on the material properties, synthesis method, and other possible species that can be formed during activation. (Nat. Mater. 2017,16,225–229)b. This quantitative partitioning was determined by growing crystals of the 1:1 host–guest complex between ExBox4+ and corannulene. (Nat. Chem. 2014, 6177–178)c. They suggested that the Au particle size may be the decisive factor for achieving highly active Au catalysts.(Acc. Chem. Res., 2014, 47, 740–749)d. Low-valent late transition-metal catalysis has become indispensable to chemical synthesis, but homogeneous high-valent transition-metal catalysis is underdeveloped, mainly owing to the reactivity of high-valent transition-metal complexes and the challenges associated with synthesizing them. (Nature2015, 517,449–454)e. The polar effect is a remarkable property that enables considerably endergonic C–H abstractions that would not be possible otherwise. (Nature 2015, 525, 87–90)f. Advances in heterogeneous catalysis must rely on the rational design of new catalysts. (Nat. Nanotechnol. 2017, 12, 100–101)g. Likely, the origin of the chemoselectivity may be also closely related to the H bonding with the N or O atom of the nitroso moiety, a similar H-bonding effect is known in enamine-based nitroso chemistry. (Angew. Chem. Int. Ed. 2014, 53: 4149–4153)2. 有很大潜力a. The quest for new methodologies to assemble complex organic molecules continues to be a great impetus to research efforts to discover or to optimize new catalytic transformations. (Nat. Chem. 2015, 7, 477–482)b. Nanosized faujasite (FAU) crystals have great potential as catalysts or adsorbents to more efficiently process present and forthcoming synthetic and renewable feedstocks in oil refining, petrochemistry and fine chemistry. (Nat. Mater. 2015, 14, 447–451)c. For this purpose, vibrational spectroscopy has proved promising and very useful. (Acc Chem Res. 2015, 48, 407–413.)d. While a detailed mechanism remains to be elucidated and there is room for improvement in the yields and selectivities, it should be remarked that chirality transferupon trifluoromethylation of enantioenriched allylsilanes was shown. (Top Catal. 2014, 57: 967. )e. The future looks bright for the use of PGMs as catalysts, both on laboratory and industrial scales, because the preparation of most kinds of single-atom metal catalyst is likely to be straightforward, and because characterization of such catalysts has become easier with the advent of techniques that readily discriminate single atoms from small clusters and nanoparticles. (Nature 2015, 525, 325–326)f. The unique mesostructure of the 3D-dendritic MSNSs with mesopore channels of short length and large diameter is supposed to be the key role in immobilization of active and robust heterogeneous catalysts, and it would have more hopeful prospects in catalytic applications. (ACS Appl. Mater. Interfaces, 2015, 7, 17450–17459)g. Visible-light photoredox catalysis offers exciting opportunities to achieve challenging carbon–carbon bond formations under mild and ecologically benign conditions. (Acc. Chem. Res., 2016, 49, 1990–1996)3. 因此同义词:Therefore, thus, consequently, hence, accordingly, so, as a result这一条比较简单,这里主要讲一下这些词的副词词性和灵活运用。
The research of RFID technology and the bus stops automatically What is the RFID?RFID is the abbreviation for Radio Frequency Identification, called electronics label .RFID recognition is a non-contact automatic identification technology, RFID signal through its automatic target recognition and access to relevant data, identify work without manual intervention, it can work in various environments. RFID technology can identify high-speed moving objects and can identify multiple tags, the operation is fast and convenient.RFID is a technology which uses radio communication between tags and readers to automatically identify the locations of items. In a networked environment of RFID readers, enormous data are generated from the proliferation of RFID readers. The raw data generated from the readers cannot be directly used by the application because it consists of enormous volume of data duplication, false positive, and false negative. Thus, the RFID data repositories must cope with a number of quality issues. These data quality issues include data redundancy, false positive, and false negative. Poor data quality has adverse effects at the operational, tactical, and strategic levels of an organization. This is especially true in the healthcare field where cost pressures and the desire to improve patient care drive efforts to integrate and clean organizational data.What is the basic component of RFID?The most basic RFID system consists of three parts:Tag:Tag consists of components and chips, each Tag has only electronic coding, adhere to the object for identifiering target;Reader:read (sometimes also can write) label information equipment,it canbe designed for portable or fixed;Antenna: Transmit radio frequency signals between in the label and reader. What is the basic principle of RFID technology ?The basic principle of RFID technology is not complex: Label into the field, Receive radio frequency signals from reader collision, with all the energy stored in a chip of the product information (passive tag, there is no source label or labels ), Oractively to send a frequency signals (active tag and to label or labels );to read information and understanding of the decoder after the central information systems to carry out the relevant data processing.Basic working process of the RFID systemThe RFID system of basic workflow is: reader through the antenna send certain frequencies of RFID signals, when the RFID card to enter the antenna working area induced current, RF cards gain energy to be activated,RF cards will own coding information through the card built-in transmitting antenna send out, System receiving aerial from RF card from the carrier signal, the antenna of a regulator to the reader, the reader to receive a signal and demodulates and decodes to the system to deal with the relevant ; the main system based on logic, determine the legality of smart cards, in different settings make the appropriate treatment and control signals control and direction of movement. In the coupling between inductors and electromagnetic way , communication process (FDX, HDX, SEQ)、From RF card to the reader of data transmission of the load method (modulation, reverse scattering, high time harmonics) and frequency, from the contact method transmission a fundamental difference, but all the reader in principle, and the decision of the design structures are very similar. all the reader is a simple matter of high frequency and control unit two basic module. high frequency interface includes both transmitters and receivers, its functions include: to produce high-frequency transmit power to start and provide RF card energy. To launch signal used to send data to RF cards, Receive and demodulation of high frequency signals from RF cards. Different RFID system with some difference frequency interface design, the system frequency inductive coupling interface diagram shown.quartzoscillatormodulator output stage amplifier antband passamplifier output stage TXD RXDReaders of the control unit features include: communicate with the application software, Application software and to execute the order of radio frequencies ;control and communication from the principle of the lord;signal of the decoding. to some specific systems are the collision, the algorithm to RF card reader, and to the transfer of data encryption and decryption, as well as on radio frequencies and the reader's authentication for an additional function.The RFID system is a key and distance of the parameters. At present, the price of long-distance RFID system is very expensive, thus to improve their reading for distance of the method is very important. Influence factors of distance and RF card reader, including antenna working frequency of RF output power, reader's reception sensitivity, RF card power, antenna and the resonant circuit Q value, antenna, RF card reader and the direction of the coupling, and RF card itself of energy and send information energy etc. Most of the system of writing is read and write different, read the distance is about 40% to 80%.The application of RFID system(1)Application in healthcareLocalization of patients, staff, supplies and equipment is very important for healthcare centers to improve service, save costs, and reduce risks. Pilot projects in hospitals demonstrate that RFID localization is a valuable approach. LANDMARC, an RFID localization system (Li et al. 2004), has been applied to facilitate the management of hospitals and other organizations in case of emergency. The system operates at the frequency of 308MHz, and is composed of active tags, landmark tags, readers, and communication devices. The position of an active tag can be determined by comparing the signal strength detected and the signal strengths of landmark tags. Using this system to monitor the infectious patients can reduce the number of healthcare professionals and ensure timely response for emergency. Meanwhile, integrated business systems and services (IBSS) announced a success pilot study for using RFID technology to locate assets and personnel at a large healthcare facility in Chicago (O’Connor 2005). Emory Healthcare successfu lly deployed 2.45GHz active RFID devices to track infusion pumps and other high-value equipment to improve asset management and utilization (O’Connor 2007). Note that the last two healthcareapplications do not employ localization algorithms, instead, they estimate the object location based on the detection from a nearby sensor. The accuracy of location can be improved by integrating localization algorithms.(2)Application in construction material managementTracking the location of construction resources enables effortless progress monitoring and supports real-time sensing of construction status. Jaselskis and ElMisalami (2003) developed a prototype RFID tracking system for improving the material procurement process on a construction site. Furthermore, Song et al. (2007) developed a method to locate materials on construction sites. The method requires a eld supervisor be equipped with an RFID reader and a GPS receiver. The GPS receiver provides the absolute position information of the supervisor, while the position of the material relative to the supervisor can be determined from the multiple communications between the reader and the tag attached to the materials.The bus is automatically stops the background and significanceThe people of car out for provides convenient service, while the bus stops directly affect the quality of the service. Traditional stops by the crew artificially, and in this way because of its poor and working intensity effect is too great, in many big ci ties have been eliminated. In recent years, with the development of science and technology progress and microcomputer technology in many fields has been widely used. In the acoustic field, with various pronunciation chip microcomputer technology ,can complete combined speech synthesis technology, makes the car stops controller is realized for citizens becomes possible, and thus provide a more personalized service. In view of the traditional bus stops system deficiency, combined with the use of public transport vehicle characteristics and practical operating environment, the design of a single-chip microcomputer control bus stops system automatically.The bus stops the design of automatic device is mainly to compensate for changing the traditional voice stops device must have driver control can work backward way, pitted ,automatic station broadcasts six-foot-tall service term for the public and provide mor e humanized more perfect service.The characteristics of the system and advantageThis system greatly improve the accuracy of bus stops, and reliability.Improving the service quality of the bus system. Promote the city economic developm ent and harmonious development of traffic changes. Made up for changing thetraditional voice stops device must have driver control can work means, in the bus sto ps behind when the station broadcasts, automatic six-foot-tall service term for the public and provide more humanized more perfect service.The present status of intelligent city bus stopsIn the process of informationization in China, a city's public transportation industry becomes the measuring level and a reference point of urban civilization for informationization as an important window. The developing level of public transportation not only represents the image of a city, but also relates with everyone in the city. The Broadcasting System for Bus well embodies services and careness for passengers.At present, the Broadcasting System installed on the bus is still operated manually with push-button, which may have potential safety hazard. In order to control the speed of each bus, Public Transport Company has to send a worker to record the time when the buses in the same line arrive at the station. The television installed on each bus transmits the same programme all day, which hardly satisfies every passenger. So the cost of advertisements broadcasted on the TV can' t drop. Based on the studies on present public transportation system, the dissertation aims to design the multi-function automatic broadcasting system for bus, which technologically combines ARM with RFID. RFID is a kind of non-contact automatic identification technology; it identifies and obtains the corresponding data automatically through the RF signal. This system can not only realize the basic function of automatic broadcasting but also do attendance checking for buses’arrival time as well. At the same time, it has realized Play AD at Real-Space-a new kind of advertising on the bus, thus enhencing the informationization level of public transpotation industry,reducing management costs, satisfing most passangers favors, improving the serving and operating level of automatic broadcasting and then makes the city more beautiful.ConclusionRFID plays an essential role in all the subdomains of the applications inhealthcare applications. The effectiveness in cleaning the RFID data in healthcare sectors remains a concern, even though a number of literary works are available.To a maximum, the dirty data that are read may even leads to patients’ death. The errors need to be cleansed in an effective manner before they are subjected to warehousing. Current solutions to correct missed readings usually use time window filtering. A serious issue is that a single static window size cannot compensate for missed readings while capturing the dynamics of tag motion. An adaptive time window filtering cannot deal with the condition that tags are always moving.In this paper, we have proposed algorithms to clean the anomalies false positive, missed readings, and duplications. It is decided to record all the values associated with each tag event for future reference otherwise too much valid data will be lost.Finally, the management can analyze the data and filter by applying business rules based on the requirement. The proposed algorithms predict and clean the anomalies based on the integration of middleware and deferred. Our experimental result proved that our algorithms predicts and removes the anomalies in an effective manner compared to the existing works. Thus, it will pave the way for an effective means of data warehousing system that will keep the RFID data safe for future mining.RFID技术及公交自动报站的研究RFID是什么?RFID射频识别的缩写,被称为电子标签。
JPhysiol574.1(2006)pp85–9385TopicalReviewDevelopingaheadforenergysensing:AMP-activatedproteinkinaseasamultifunctionalmetabolicsensorinthebrain
SantoshRamamurthy1andGabrieleV.Ronnett1,2Departmentsof1Neuroscienceand2Neurology,TheJohnsHopkinsUniversitySchoolofMedicine,Baltimore,MD21205,USAThe5-adenosinemonophosphate-activatedproteinkinase(AMPK)isametabolicandstresssensorthathasbeenfunctionallyconservedthroughouteukaryoticevolution.ActivationoftheAMPKsystembyvariousphysiologicalorpathologicalstimulithatdepletecellularenergylevelspromotesactivationofenergyrestorativeprocessesandinhibitsenergyconsumptiveprocesses.AMPKhasaprominentrolenotonlyasaperipheralsensorofenergybalance,butalsointheCNSasamultifunctionalmetabolicsensor.RecentworksuggeststhatAMPKplaysanimportantroleinmaintainingwholebodyenergybalancebycoordinatingfeedingbehaviourthroughthehypothalamusinconjunctionwithperipheralenergyexpenditure.Inaddition,brainAMPKisactivatedbyenergy-poorconditionsinducedbyhypoxia,starvation,andischaemicstroke.Undertheseconditions,AMPKisactivatedasaprotectiveresponseinanattempttorestorecellularhomeostasis.Howeverinvivo,itappearsthattheoverallconsequenceofactivationofAMPKismorecomplexthanpreviouslyimagined,inthatover-activationmaybedeleteriousratherthanneuroprotective.ThisreviewdiscussesrecentfindingsthatsupporttheroleofAMPKinbrainasamultidimensionalenergysensorandtheconsequencesofitsactivationorinhibitionunderphysiologicalandpathologicalstates.
(Received21March2006;acceptedafterrevision11May2006;firstpublishedonline11May2006)CorrespondingauthorG.V.Ronnett:1006BPreclinicalTeachingBuilding,JohnsHopkinsUniversitySchoolofMedicine,725NorthWolfeStreet,Baltimore,MD21205,USA.Email:gronnett@jhmi.edu
IntroductionItiswellestablishedthatafundamentalnecessityforsurvivalisthemaintenanceofenergyhomeostasis.Thisrequiresthepresenceofappropriateenergysensorsthatcandetectandinitiateadaptivechangesinresponsetovariationsinenergybalance.Theadenosinemonophosphate-activatedproteinkinase(AMPK)signallingcomplexisonesuchsensor.AlthoughpreviousstudiesfocusedontheroleofAMPKascellularmetabolicsensor,itisbecomingevidentthatAMPKalsoplaysmorecomplexrolesasanorganismalmetabolicsensor(Carling,2004).FunctionsdescribedforAMPKhaveincludedthecoordinationofanabolicandcatabolicmetabolicprocessesinvarioustissues,includingcardiacandskeletalmuscle,adiposetissue,pancreasandliver(Kahnetal.2005).Inthesetissues,AMPKrespondstodiversehormonal,physiologicalandpathologicalstimuli,andinturninhibitsATP-consuming(anabolic)processesandstimulatesATP-generating(catabolic)processes.Thus,
theconsequencesofAMPKactivationencompassacutemodulationofenergymetabolismandchronicchangesingeneexpression.Thisisaccomplishedbyphosphorylationandmodificationofnumeroustargetproteinsincludingbiosyntheticenzymes,transporters,transcriptionfactors,ionchannelsandcell-cycle/signallingproteins(Leff,2003;Hardie,2004a,2005;Hallows,2005).ItisonlywithinthepastseveralyearsthatthecomplexphysiologicalroleofAMPKinthebrainhasbecomeevident.IntheCNS,itappearsthatAMPKhasadualfunction,notonlyasacell-autonomousenergysensor,butalsoasanintegrativemetabolicsensor.Inthisregard,AMPKmayplayaroleintheneuronalsurvivalresponsetoenergydepletionandaswellinhypothalamiccontrolfoodintakeandperipheralenergyutilization.AnotheremergingconceptisthattheresultofAMPKactivationiscontextspecific,i.e.AMPKactivationcanbeeitherbeneficialordeleterious,dependingonthetissue,degreeofstimulation,orconditionsofactivation.ThedifferentialregulationofAMPKactivationisthereforeanotheractiveareaofinvestigation.Thus,theAMPK
C2006TheAuthors.JournalcompilationC2006ThePhysiologicalSocietyDOI:10.1113/jphysiol.2006.11012286S.RamamurthyandG.V.RonnettJPhysiol574.1
pathwayandmodulationofenergybalancethroughtheAMPKsystempresentsanattractivetherapeutictargetforinterventioninmanyconditionsofdisorderedenergybalance,includingobesity,type-2diabetes,andstroke.ThescopeofthisreviewwillbelimitedtonewdevelopmentsrelatedtoAMPKasametabolicsensorinthebrain.SeveralrecentreviewsprovidefurtherinformationontheroleoftheAMPKsignallingpathwayinthecellularphysiologyofothersystems(Kempetal.2003;Hardie,2004b;Kahnetal.2005;Luoetal.2005;Youngetal.2005).
RegulationofAMPKFunctionalAMPKisaheterotrimerickinasecomplexcomposedofacatalyticαsubunitandregulatoryβandγsubunits.Inmammals,severalalternativegeneshavebeenidentifiedforeachofthesubunits,e.g.α1,α2,β1,β2,γ1,γ2,γ3.Therefore,dependingonthetissueorcelltypeexamined,varyingcombinationsofαβγheterotrimersarepossible(Stapletonetal.1996).ThesubunitcompositioninaspecifictissueorcelltypemaybeimportanttotherolethatAMPKplaysinregardtoenergysensinginthatparticulartissue.SeveralstudieshaveexaminedAMPKsubunitexpressioninthebrainandarediscussedinfurtherdetailbelow(Turnleyetal.1999;Culmseeetal.2001).AMPKcatalyticactivityisenhancednotonlybyAMPbinding,butalsobyαsubunitphosphorylationbyanupstreamkinase(Daviesetal.1995;Hawleyetal.1996;Steinetal.2000).ThesearchfortheidentityoftheupstreamkinasesthatphosphorylateandactivateAMPKhasbeenalongone(Wittersetal.2006).However,theidentificationofthesekinasesmayprovidefurtherinsightintothetissue-specificaswellasbrain-specificregulationofAMPK.InitiallyitwasshownthatthePeutz–Jegherssyndrometumoursuppressorgene,LKB1,isanAMPKkinase(Hawleyetal.2003;Woodsetal.2003).Subsequently,studiesinvariouscell-lines,tissuesandinvivomodelsdemonstratedthephysiologicalimportanceofactivationoftheLKB1-AMPKsignallingpathway(Shawetal.2004,2005;Jonesetal.2005;Sakamotoetal.2005).However,despitethefactthatLKB1hasbeenreportedtobewidelyexpressedinvarioustissues,itsroleinbrainAMPKregulationhasnotyetbeenelucidated(Rowanetal.2000).Mostrecently,threegroupsindependentlyreportedthatcalcium/calmodulin-dependentproteinkinaseβ(CaMKKβ)isanadditionalupstreamkinaseofAMPK(Hawleyetal.2005;Hurleyetal.2005;Woodsetal.2005).Althoughtheprecisephysiologicalrelevanceofthisactivationisunclear,severalpossibilitiesexist.SinceCaMKKβisaCa2+-activatedkinase,itispossible