Regulatory volume increase is associated with p38 kinase-dependent actin cytoskeleton remodeling in
- 格式:pdf
- 大小:836.41 KB
- 文档页数:13

网络出版时间:2021-9-714:47 网络出版地址:https://kns.cnki.net/kcms/detail/34.1065.R.20210907.1405.031.htmlCP 25调控GRK2/p38MAPK信号在血管紧张素Ⅱ诱导小鼠系膜细胞增殖中的作用王 祥,袁晓阳,徐 靓,江 丽,王盛付,黄李娜,魏 伟,严尚学2021-06-04接收基金项目:国家自然科学基金(编号:81330081)作者单位:安徽医科大学临床药理研究所,抗炎免疫药物教育部重点实验室,抗炎免疫药物安徽省协同创新中心,合肥 230032作者简介:王 祥,男,硕士研究生;魏 伟,男,教授,博士生导师,责任作者,E mail:wwei@ahmu.edu.cn;严尚学,男,副研究员,硕士生导师,责任作者,E mail:yan shx@163.com摘要 目的 研究芍药苷 6′ O 苯磺酸酯(CP 25)在血管紧张素Ⅱ(AngⅡ)诱导的肾小球系膜细胞(MCs)增殖中的作用及相关机制。
方法 体外培养SV40MES13系膜细胞,以AngⅡ诱导MCs增殖,高内涵成像显微镜检测不同浓度CP 25(10、100、1000nmol/L)对AngⅡ诱导的MCs增殖的影响;Westernblot检测MCs中G蛋白偶联受体激酶 2(GRK2)和磷酸化p38(p p38)蛋白表达水平;激光共聚焦显微镜检测GRK2和p p38蛋白的荧光信号并分析二者共定位率;成像流式细胞仪检测MCs中GRK2入胞质的细胞比例。
结果 与对照组比较,AngⅡ可诱导MCs的增殖,增加GRK2和p p38蛋白表达,上调GRK2和p p38MAPK共定位率,提高GRK2入胞比例,差异有统计学意义(P<0 01)。
CP 25(10、100、1000nmol/L)可不同程度地抑制AngⅡ诱导的增殖,抑制GRK2、p p38蛋白表达及GRK2和p p38的共定位水平,降低GRK2入胞比例。

p38 MAPK通路在造血系统调节中的作用李德冠;樊飞跃;孟爱民【摘要】Mitogen-activated protein kinase( MAPK ) superfamily is an important signal mediating many cellular reactions. The subfamily p38 MAPK plays an important role in the regulation of hematopoietic system. Inhibition of p38 MAPK pathway can alleviate the function decline of hematopoietic cells caused by senescence or deseases. To further research the substrates of p38 MAPK pathway in hematopoietic cells will provide new ideas for the clinical application of hematopoietic stem cells and mechanisms of hematopoietic system diseases.%丝裂原活化蛋白激酶( MAPK)超家族是介导细胞反应的重要信号系统.研究发现,该超家族亚族p38 MAPK通路在造血细胞调节过程中占据重要地位,抑制p38 MAPK通路可一定程度缓解衰老或者疾病引发的造血细胞功能下降.深入研究p38 MAPK通路在造血系统中的下游作用底物,将对临床造血干细胞的应用以及造血系统疾病发病机制研究提供新的思路.【期刊名称】《中国药理学通报》【年(卷),期】2011(027)001【总页数】3页(P4-6)【关键词】造血系统;丝裂原活化蛋白激酶;p38;活性氧;刺激;造血干细胞【作者】李德冠;樊飞跃;孟爱民【作者单位】中国医学科学院放射医学研究所、天津市核医学重点实验室,天津,300192;中国医学科学院放射医学研究所、天津市核医学重点实验室,天津,300192;中国医学科学院放射医学研究所、天津市核医学重点实验室,天津,300192【正文语种】中文【中图分类】R-05;R329.25;R331.23;R345.57;R977.3丝裂原活化激酶(mitogen-activated protein kinase,MAPK)是哺乳细胞内广泛存在的一类丝氨酸/苏氨酸蛋白激酶,是细胞重要的应激通路,在细胞生长、发育、分化、凋亡等许多生理过程中占据重要地位。

磷脂酰肌醇-3-激酶家族与急性肺损伤研究新进展华中科技大学同济医学院附属协和医院麻醉科(430022)王月兰姚尚龙摘要磷脂酰肌醇-3-激酶以其广泛的分布,参与并产生酯类第二信使,激活细胞内大量酶联级反应,调节多种细胞的生存、激活、分化和繁殖而成为研究细胞信号转导机制的热点。
该文重点介绍PI3K结构与功能、激活途径及参与急性肺损伤的机制及其临床意义。
关键词肺损伤;磷脂酰肌醇脂激酶;机制近年来对急性肺损伤(ALI)机制的研究发现,除了目前正在研究的MAPK信号转导通路对ALI影响外,越来越多的试验和临床发现PI3K在急性肺损伤的形成中有着不可忽视的作用。
并且还发现PI3K是肺细胞重要的调节酶之一,影响着肺组织的生理和病理变化【1】。
因此对PI3K及其下游底物的研究,可将成为研究和治疗肺部疾病的新靶点。
1 PI3Ks结构、功能与生物活性磷脂酰肌醇-3-激酶(phosphatidylinositol 3-kinases,PI3K)的研究始于1980s后期。
近年来对PI3Ks家族的构成、功能研究有了新的突破。
PI3Ks 是普遍存在于体内各类细胞,属于酯类的激酶(或酶),能磷酸化与膜相关的磷脂酸肌醇家族,它可以募集和激活下游的靶物质而启动一系列信号联级反应,其在细胞的有丝分裂发生、细胞存活、分化和激活、细胞骨架的构型与重塑以及囊胞的运输起着重要的作用[1]。
1. 1 结构与分类据其结构、特异底物和不同调节功能PI3Ks可分为三大类[2,3]。
其中PI3-Ks家族已有9个成员从哺乳类生物细胞中分离出来。
I类:由P110催化亚基和调配亚基(regulatory adapter subunit):P50、P55、P85、P101构成的异二聚体,其中P110又有四种亚型分别为:P110α、P110β、P110γ和P110δ。
目前已经明确的I类PI3Ks又有两个亚家族,分别为:IA和IB。
IA是由p110α、p110β和p110δ催化亚单位和调节亚基(P85α、P85β、P55γ)构成,对酪氨酸激酶相关受体(tyrosine kinase-lingked receptor )的信号转导系统比较敏感。


20解剖学研究2021年第43卷第丨期Anat Res,2021,Vol.43, No.l•论著•PI3K/Akt诱导激活F0X03a可防止TNF-〇a秀导的GnRH下降郭晗,张本斯,施荣杰、施雨辰*12*,石纯(大理大学基础医学院人体解剖学教研室,云南大理671003)【摘要】目的探究P13K/Akt通路对T N F-a介导的下丘脑GnRH释放减少的调节作用和机制。
方法采用F0X03a敲低的GT1 -7细胞系,用P I3K激活剂740Y-P和T N F-a处理细胞,采用ELISA检测培养基中的GnRH水 平。
用Western blot检测细胞中(p)-lKBot和p-A kt的磷酸化蛋白含量,用免疫荧光法对F0X03a和NF-k B的核定位 进行检测。
结果PI3K/Akt通路的激活可促进F0X03a核易位,进而逆转了 T N F-a诱导的GNRH1基因抑制和NF-k B激活。
通过这一机制可防止TNF-tx对GT1-7细胞释放GnRH的抑制效应。
结论PI3K/Akt通路的激活有助于 防止衰老相关的下丘脑GnRH释放减少。
【关键词】促性腺激素释放激素;衰老;NF-k B;F0X03aPI3K/Akt-induced activation of F0X03a prevents TNF-a-induced GnRH decline GUO Han, ZHANG Ben-si, SHI Rong-jie, SHI Yu-chen t SHI Chun.Department o f Human Anatomy of Basic Medical College, Dali University, Dali, Yunnan Province 671003, ChinaCorresponding author: SHI Chun, E-mail:*******************【Abstract】Objective To investigate the effect of PI3K/Akt on TNF-a-induced hypothalamic GnRH decline. Meth-ods GT1-7 cells were treated with PI3K activator (740 Y-P) and TNF-a. GnRH level in the medium was detected by ELISA method. p-lKBa and p-Akt were analyzed using western blot. The nuclear localization of F0X03a and NF-k B were analyzed by using western blot and immunofluorescence. Results The current study revealed that PI3K/Akt-induced F0X03a nuclear translocation protects GnRH neurons from TNF-a-induced decrease of GnRH release through preventing TNF-a-induced GNRH1 gene inhibition and NF-k B activation. Conclusion The results of the present study provided novel insights into the mechanisms underlying aging-associated hypothalamic GnRH decline.【Key words】Gonadotropin-releasing hormone; Aging; Nuclear factor-k B; Forkhead box protein 03a下丘脑是中枢神经内分泌系统的重要组成部 分,可调节脑与周围神经系统之间的相互作用'_2]。


hippocampus after hyp〇xia-ischemia(J]. Glia, 2015,63(12): 2220-2230. [18] Wang S, Xue H, Xu Y, et al. Sevoflurane postconditioning inhibitsautophagy through activation of the extracellular signal-regulated kinase cascade, alleviating hypoxic-ischem ic brain injury in neonatal rats[J]. Neurochem Res, 2019, 44(2):347-356.[19] Li X, Wang MH, Qin C, et al. Fingolimod suppresses neuronalauophagy through the mTOR/p70S6K pathway and alleviates ischemic brain damage in mire[J]. PLoS One, 2017, 12(1 l):e0188748.[20] Yan YQ, Zhang B, Wang L, et al. Induction of apoptosis andautophagic cell death by the vanillin derivative 6-brom ine-5-hydroxy-4-methoxybenzaldehyde is accompanied by the cleavage ofDNA-PKcs and rapid destruction of c-Myc oncoprotein in HepG2cells[J]. Cancer Lett, 2007, 252(2):280-289.[21 ]Requejo-Aguilar R, Lopez-Fabuel I, Fernandez E, et al. PINK1deficiency sustains cell proliferation by reprogramming glucosemetabolism through HIF1[J]. Nat Commun, 2014,5: 4514.[22] Devireddy S, Liu A, I^ampe T, et al. The organization of milochondrialquality control and life cycle in the nervous system in vivo in theabsence of PINKlfJ]. J Neurosci, 2015, 35(25): 9391-9401.(收稿日期:2020-10-10)•论著•芍药苷对急性脑梗死大鼠小胶质细胞激活调节的神经元焦亡的作用祖力菲亚•艾克木麦合莆热提•乌莆尔邢辉辉樊明新张俊江【摘要】目的研究芍药苷对急性脑梗死(AC1)大鼠神经元焦亡的影响。
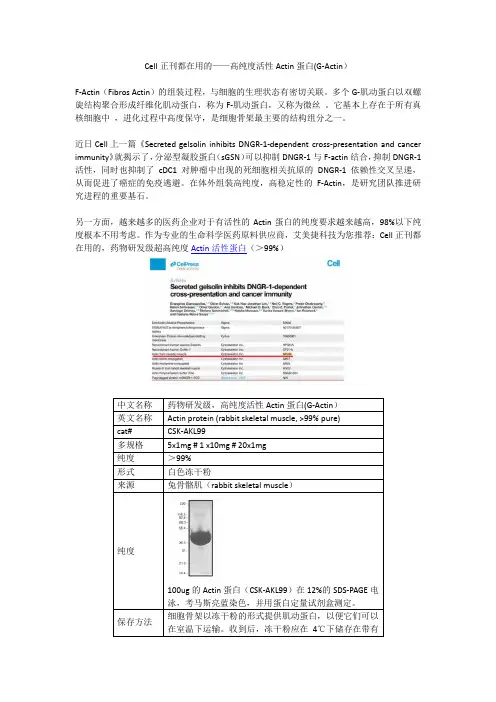
Cell正刊都在用的——高纯度活性Actin蛋白(G-Actin)F-Actin(Fibros Actin)的组装过程,与细胞的生理状态有密切关联。
多个G-肌动蛋白以双螺旋结构聚合形成纤维化肌动蛋白,称为F-肌动蛋白,又称为微丝。
它基本上存在于所有真核细胞中,进化过程中高度保守,是细胞骨架最主要的结构组分之一。
近日Cell上一篇《Secreted gelsolin inhibits DNGR-1-dependent cross-presentation and cancer immunity》就揭示了,分泌型凝胶蛋白(sGSN)可以抑制DNGR-1与F-actin结合,抑制DNGR-1活性,同时也抑制了cDC1 对肿瘤中出现的死细胞相关抗原的DNGR-1 依赖性交叉呈递,从而促进了癌症的免疫逃避。
在体外组装高纯度,高稳定性的F-Actin,是研究团队推进研究进程的重要基石。
另一方面,越来越多的医药企业对于有活性的Actin蛋白的纯度要求越来越高,98%以下纯度根本不用考虑。
作为专业的生命科学医药原料供应商,艾美捷科技为您推荐:Cell正刊都在用的,药物研发级超高纯度Actin活性蛋白(>99%)100ug的Actin蛋白(CSK-AKL99)在12%的SDS-PAGE电泳,考马斯亮蓝染色,并用蛋白定量试剂盒测定。
细胞骨架以冻干粉的形式提供肌动蛋白,以便它们可以Cell文章结果验证Actin Protein ( >99% Pure): Rabbit Skeletal Muscle:(A-C)体外聚合F-肌动蛋白(最高浓度0.2 mM)或无F-肌动蛋白(PBS;箭头)的连续(2 倍)稀释(箭头)被点到膜上。
在用(A) 指定剂量的FCS、(B) ABP 耗尽或模拟耗尽FCS 和(C) sGSN 预处理膜后,检测到DNGR-1 ECD (5 mg/mL) 与点的结合或cofilin(均为10mg/ml)。

细胞增殖信号通路调控机理细胞增殖是生物体维持生命所必需的过程之一,人体在成长、修复、繁殖和癌症等多个方面均需要细胞增殖。
细胞增殖不仅仅涉及到称为“阀门”的基因的开关,还需要增殖信号通路的精准调控。
细胞增殖信号通路调控机理是一个重要的研究领域,在医学上具有广泛的应用前景。
1.细胞增殖信号通路的基本架构细胞增殖信号通路是一个信号转导网格,由许多元件组成。
当外来信号或细胞自身环境的信号引发细胞内的分子反应时,就会产生一系列级联反应,从而引起细胞核内的基因表达变化,最终导致细胞增殖。
其中较为典型的细胞增殖信号通路包括:MAPK信号通路、PI3K-Akt信号通路和Wnt/beta-catenin信号通路。
MAPK信号通路是由ERK、JNK和p38组成的,这些酶是在细胞外信号激活后,经过磷酸化而激活的。
PI3K-Akt信号通路是一条反应细胞的营养水平的通路,主要是指抑制自噬和促进增殖之间的平衡调节。
Wnt/beta-catenin信号通路是关键的调节发育和细胞增殖的信号通路之一。
2.蛋白激酶与细胞增殖信号通路蛋白激酶是组成细胞增殖信号通路的基本分子,在细胞内发挥重要作用。
除了MAPK并不是一类特殊的酶之外,蛋白激酶的分类非常多。
在细胞增殖信号通路中主要涉及到的激酶包括:ERK1/2、JNK、p38MAPK、AKT、mTOR、EGFR、PI3K等。
3.信号转导通路的调控细胞增殖信号通路完整的调控网络是由多个分子的相互作用而构成的。
细胞环境的变化会导致信号转导通路的扰动,进而影响到分子通路的活动。
细胞增殖信号通路的调控可以分为两种类型:内在调控和外在调控。
内在调控主要涉及到细胞分子自身的反应过程,包括蛋白酶的磷酸化、内质网的应答、RNA后期修饰和基因表达的调控等。
而外在调控则是对细胞外的刺激做出反应,从而影响信号转导通路的活动。
4.细胞增殖信号通路与疾病细胞增殖信号通路在人体生理、病理以及临床诊断和治疗中都扮演着重要的角色。

p38MAPK及ERK信号通路激活在CD151促进内皮细胞迁移中的作用机制刘曌宇;曾和松;李鹏程;费宇杰;左后娟【摘要】目的研究p38MAPK信号通路及ERK信号通路激活在CD151促进人脐静脉内皮细胞(HUVECs)迁移中的机制.方法包装携带CD151、antiCD151和GFP 的重组腺相关病毒,转染HUVECs.Western blot检测HUVECs 中CD151、细胞外信号调节激酶(ERK)、p38丝裂原活化蛋白激酶(MAPK)蛋白的表达;利用改良的Boyden趋化小室进行细胞迁移检测;在细胞转染同时分别给予ERK抑制剂(PD98059,20 μmol/L),或p38MAPK抑制剂(SB203580,20μmol/L),然后检测细胞迁移.结果①CD151组较空白对照组及GFP组明显促进细胞迁移的能力,而antiCD151组则显著抑制细胞的迁移,差异具有统计学意义(P<0.05).②CD151高表达促进ERK信号通路激活,而对p38MAPK信号通路无显著作用.③ERK抑制剂(PD98059)显著抑制CD151转染后的促HUVECs迁移的作用(P<0.05),而p38MAPK抑制剂(SB203580)无明显作用.结论 CD151具有促进细胞迁移的作用,CD151通过激活ERK信号通路促进细胞迁移,而p38MAPK信号通路的激活则在CD151促进HUVECs迁移中无显著作用.【期刊名称】《华中科技大学学报(医学版)》【年(卷),期】2015(044)003【总页数】5页(P258-262)【关键词】CD151;人脐静脉内皮细胞;细胞外信号调节激酶;丝裂原活化蛋白激酶;细胞迁移【作者】刘曌宇;曾和松;李鹏程;费宇杰;左后娟【作者单位】华中科技大学同济医学院附属同济医院消化内科,武汉430030;华中科技大学同济医学院附属同济医院心血管内科,武汉430030;华中科技大学同济医学院附属同济医院心血管内科,武汉430030;华中科技大学同济医学院附属同济医院心血管内科,武汉430030;华中科技大学同济医学院附属同济医院心血管内科,武汉430030【正文语种】中文【中图分类】R329.28血管再生性治疗已成为当前冠心病基础和临床研究的热点。
p38 MAPK在腹泻型肠易激综合征大鼠中的变化及其免疫调控作用郭军雄; 汪斌; 马丽; 康万荣; 许小敏; 徐生刚【期刊名称】《《中国实验动物学报》》【年(卷),期】2019(027)006【总页数】7页(P709-715)【关键词】肠易激综合征; 腹泻型; p38MAPK信号通路; 白细胞介素1β; 白细胞介素6; 肿瘤坏死因子α; 大鼠【作者】郭军雄; 汪斌; 马丽; 康万荣; 许小敏; 徐生刚【作者单位】河西学院医学院河西学院丝绸之路中医药研究中心河西学院中西医结合研究所甘肃张掖 734000; 甘肃中医药大学兰州 730000【正文语种】中文【中图分类】Q95-33肠易激综合征(irritable bowel syndrome,IBS)是一种以腹痛或腹部不适并常伴排便习惯改变为特征的慢性肠功能紊乱性疾患,胃肠动力和内脏感知异常是IBS发病的主要病理生理学特征,迄今具体发病机制尚无完全定论[1-2]。
近年来,随着生活节奏增快和压力增大,发病率有上升的趋势[3]。
而免疫系统作为D-IBS精神-肠道因素互动的关键纽带,其平衡对D-IBS内脏敏感性有重要影响[4]。
前期研究表明[5]:慢性束缚(S)联合番泻叶(F)灌服法复建的大鼠Th1促炎因子IL-12的含量升高,Th2 抑炎因子IL-10的含量降低,可能存在Th1/Th2平衡漂移。
p38 MAPK是MAPK家族中最重要的成员之一,其作为细胞信号传递的交汇点,能被应激、氧化反应以及炎性细胞因子等多种因素所激活。
有研究表明[6-7]:p38 MAPK信号通路系统在疼痛感知、胃肠动力紊乱和免疫炎症异常激活中均有重要作用。
本研究旨在通过观察D-IBS模型大鼠结肠组织中p38 MAPK蛋白的表达和血清中IL-1β、IL-6、TNF-α的变化及其相关性,探讨p38 MAPK通路在S+F灌服法诱发的D-IBS“肝郁脾虚证”大鼠模型中的免疫作用。
1 材料与方法1.1 材料1.1.1 实验动物SPF级Wistar大鼠40只(雌雄各半),8~10周龄,体重(180±20)g,购自甘肃中医药大学科研实验中心【SCXK(甘)2015-0002)】。
中国细胞生物学学报Chinese Journal of Cell Biology2021,43(3): 499-507DOI: 10.11844/cjcb.2021.03.0002研究论文长链非编码RNA KCNQIOTl影响脂多糖诱导的血管内皮细胞凋亡及其炎性因子表达的机制谢斌1邓超1陈栩栩1王红梅2苏醒1:11G中南大学湘雅医学院附属海口医院重症医学科,海口570203;2中南大学湘雅医学院附属海口医院检验科,海口570203)摘要 该研究探讨了长链非编码RNA KCN Q IO Tl对脂多糖(LPS)诱导的血管内皮细胞(VEC)凋亡和炎性因子表达的影响以及其可能机制。
通过体外培养VEC,分别转染K C N Q IO T l过 表达栽体、miR-223抑制剂或共转染KCNQ1OT1过表达载体与miR-223模拟物后,用1.0mg/mL LPS 干预24 h,然后采用RT-qPCR法检测细胞中KCN Q IO Tl和miR-223的表达水平,流式细胞仪检测细 胞凋亡,Western blot检测细胞中Bcl-2和Bax蛋白表达,ELISA试剂盒检测细胞培养上清中TNF-a、IL-1和IL-6水平。
双荧光素酶报告基因实验验证KCNQ1OT1与miR-223的调控关系。
结果显示,LPS可抑制V E C中KCNQ1O T1的表达,而促进miR-223表达;上调KCNQ1OT1或下调miR-223后均 可降低LPS诱导的V E C凋亡率、Bax蛋白及TNF-a、IL-1和IL-6表达(P<0.05),而促进Bcl-2蛋白表达 (尸<0.05)。
K C N Q IO Tl靶向负调控miR-223表达,上调miR-223则逆转上调KCN Q IO Tl对LPS诱导 的V E C凋亡及炎性因子表达的抑制作用。
这表明,上调KCN Q IO Tl抑制LPS诱导的V E C凋亡及炎 性因子表达,其作用机制可能与靶向负调控miR-223有关,K C N Q lO Tl/miR-223轴可能为血管内皮 细胞损伤的治疗提供了新靶点。
网络出版时间:2013-04-10 09:23网络出版地址:/kcms/detail/51.1705.R.20130410.0923.002.html叶酸抑制同型半胱氨酸诱导的人肾脏系膜细胞增殖曹露,娄晓盈,牟娜娜,张泉,谭红梅中山大学中山医学院病理生理教研室,广东广州510080△[基金项目] 国家自然科学基金项目(NO.30600250),广东省自然科学基金项目(NO.10151008901000146)及高校基本科研业务费中山大学青年教师培育项目(NO.10ykpy33)资助。
【摘要】目的观察叶酸对同型半胱氨酸(homocysteine, Hcy)诱导的人肾脏系膜细胞增殖作用的影响及其可能机制。
方法人肾脏系膜细胞采用含5%胎牛血清的RPMI1640培养,采用不同浓度Hcy(200、400、600、800、1000μmol/L)处理细胞,MMT法观察Hcy对人肾脏系膜细胞增殖的影响。
选取理想的Hcy浓度,加入不同浓度叶酸(50、100、200、400μmol/L)处理细胞,观察叶酸对细胞增殖的干预作用。
相同方法处理细胞后提取总蛋白,western blot检测转化生长因子-β(transforming growth factor-β, TGF-β)和细胞外调节蛋白激酶(extracellular regulatedprotein kinases, ERK)表达。
结果Hcy能诱导人肾脏系膜细胞的增殖,促进TGF-β和ERK蛋白表达,呈现剂量依赖性。
叶酸干预后能抑制Hcy诱导的细胞增殖,下调TGF-β和ERK蛋白表达。
结论叶酸能抑制Hcy诱导的人肾脏系膜细胞增殖,可能与抑制TGF-β和ERK蛋白表达有关。
【关键词】同型半胱氨酸;叶酸;转化生长因子-β;细胞外调节蛋白激酶;人肾脏系膜细胞Folic acid attenuated homocysteine-induced proliferation of human renal mesangial cellsLu Cao, Xiaoying Lou, Nana Mou, Quan Zhang, Hongmei TanDepartment of Pathophysiology, Zhongshan School of Medicine, Sun Yat-sen UniversityGuangzhou, 510080, China.Correspondencing author: Hongmei Tan, MD, PhD, Associate professor, Department ofPathophysiology, Zhongshan School of Medicine, Sun Yat-sen University, 74 ZhongshanRoad II, Guangzhou 510080, China, Tel: (8620) 87330028, Fax: (8620) 87330026,E-mail: tanhm@【Abstract】Objective This study aimed to investigate the effect of folic acid (FA) onhomocysteine (Hcy)-induced proliferation of human renal mesangial cells (HMCs) andpossible signaling mechanisms. Methods HMCs were cultured in RPMI1640 mediumsupplemented with 5% fetal bovine serum. HMCs were treated with PBS, Hcy (200, 400,600, 800 and 1000μmol/L) or Hcy (400μmol/L) plus folic acid (50、100、200、400μmol/L)for 24 hours. Cells viability were assessed by using Methylthiazolyldiphenyl-tetrazoliumbromide (MMT) assay and the alterations of transforming growth factor-β (TGF-β) and extracellular regulated protein kinases (ERK) were detected by western blot analysis. Results Hcy significantly stimulated the proliferation of HMCs in a dose dependent manner. However, Hcy-induced proliferation of HMCs was attenuated by additional supplement of FA. Protein levels of TGF-β and ERK were increased in the presence of Hcy, which was reversed by additional supplement of FA. Conclusion FA attenuated Hcy-induced proliferation of HMCs, which was associated with decreased expression of TGF-β and ERK protein.[Key words] Homocysteine; Folic acid; Transforming growth factor-β; Extracellular regulated protein kinases; Human renal mesangial cells同型半胱氨酸(homocysteine, Hcy)是蛋氨酸转甲基过程中的一个产物,高同型半胱氨酸血症(hyperhomocysteinemia, HHcy)是心血管疾病的独立危险因素[1]。
NAADP 对溶酶体数量的影响。
采用LDL 受体敲除(LDLr −/−)小鼠的骨髓源性巨噬细胞作为观察对象,以更好地模拟高胆固醇的细胞环境。
结果显示,NAADP 可明显促进巨噬细胞中溶酶体数量的增加(溶酶体再生增强),这种效应可被NAADP 拮抗剂NED -19、Ca 2+螯合剂BAPTA 及Cn (Ca 2+直接激活的下游分子)抑制剂CsA 明显抑制(P <0.05);有趣的是,溶酶体数量增加后,其分布范围朝细胞边缘明显靠近(P <0.05),见图1。
2 CD38促进LDLr −/−巨噬细胞中NAADP 的合成为明确CD38和NA 在LDLr −/−巨噬细胞NAADP 合成中的作用,我们检测了NA 对细胞内NAADP 水平的影响;同时将LDLr −/−小鼠的CD38基因敲除,以确定CD38对NAADP 合成的作用。
结果显示:加入NAADP 合成底物NA 后,经oxLDL 处理LDLr −/−巨噬细胞中的NAADP 水平显著升高(P <0.05);该现象在CD38基因敲除LDLr −/−巨噬细胞中消除(P <0.05),见图2。
3 NA 促进CD38 mRNA 和蛋白表达为探讨NA 与CD38之间是否存在相互作用的关系,我们观察了NA 对CD38 mRNA 和蛋白表达的影响。
为了排除不同基因背景下CD38表达的异同,我们同时观察了NA 对野生型(wild type , WT )和LDLr −/−小鼠巨噬细胞CD38表达的调节。
结果显示,不论是在WT 还是在LDLr −/−巨噬细胞中,NA 均可促进CD38Figure 1. NAADP increased lysosome number in LDLr −/− macrophages. Macrophages were incubated with oxLDL (40 mg/L ) and DiI -oxLDL (1 mg/L ) for 48 h , followed by being chased for 24 h , then incubated with NAADP (100 nmol/L ) with or withoutNAADP antagonist NED -19 (100 µmol/L ), calcium chelator BAPTA (5 µmol/L ) or calcineurin inhibitor CsA (5 µmol/L ) and proceeded to live cell confocal microscopic scanning (duration 12 h , interval 2 h ). A : real -time (living cell ) confocal microscopic images of DiI -oxLDL staining ; B : summarized lysosome number calculated with Image -Pro Premier software ;C : percentage of lysosome closer to cell edge analyzed with Image -Pro Premier software. Mean±SD. n =6. *P <0.05 vs con‐trol group.图 1 NAADP 增加LDLr −/−巨噬细胞溶酶体数量31的mRNA 和蛋白表达,尤其对LDLr −/−巨噬细胞的作用更明显(P <0.05),见图3。
图7APN 各组大鼠子宫内膜胞饮突发育的影响Fig.7Effects of APN on the development of endometrial pinopodes (PP)in each group.Red arrows indicate the pinopodes.P C O S +A P N +V E H P C O S +A P N +G WP C O S +A P NP C O S C O N ×5000×20000P COS +A P N +V E H P C O S +AP N +G W P CO S+A P NP CO S C ON 120%100%80%60%40%20%0%P r e g n a n c y r a tePC O S +A P N+VE HP CO S +A PN +G W PC O S +APN PC O S C ONP CO S+A P N+VE HP CO S+A P N+GW P CO S +A P NP CO S C ON*******N u m b e r o f e m b r y o s20181614121086420图8APN 对各组大鼠妊娠的影响Fig.8Effects of APN on pregnancy of the rats ineach group.A :Statistics of pregnancy rate in each group.B :Pregnant uterine specimens of the rats in each group.C :Statistics of the number of embryos.*P <0.05,**P <0.01,***P <0.001.BCAJ South Med Univ,2024,44(2):298-307··306参考文献:[1]Kostroun KE,Goldrick K,Mondshine JN,et al.Impact of updated international diagnostic criteria for the diagnosis of polycystic ovary syndrome[J].F&S Rep,2022,4(2):173-8.[2]李予,苏圣梅,杨冬梓.多囊卵巢综合征患者子宫内膜容受性的特点[J].生殖与避孕,2014,34(7):571-4.[3]Huang L,Liang A,Li T,et al.Mogroside V improves follicular development and ovulation in young-adult pcos rats induced by letrozole and high-fat diet through promoting glycolysis[J].Front Endocrinol(Lausanne),2022,Mar28Online ahead of print.[4]Jiang NX,Li XL.The disorders of endometrial receptivity in PCOS and Its mechanisms[J].Reprod Sci,2022,29(9):2465-76.[5]Pathare ADS,Hinduja I,Mahadik RC.Basic aspects of endometrial receptivity in PCOS patients[J].Mol Biol Rep,2022,49(2):1519-28.[6]龚斐,郭慧,彭炬,等.改良超长方案改善多囊卵巢综合征(PCOS)不孕患者子宫内膜容受性的研究[J].生殖与避孕,2015,35(5): 300-9.[7]Shan H,Luo R,Guo X,et al.Abnormal endometrial receptivity and oxidative stress in polycystic ovary syndrome[J].Front Pharmacol, 2022,Jul25Online ahead of print.[8]Achari AE,Jain SK.Adiponectin,a Therapeutic target for obesity, diabetes,and endothelial dysfunction[J].Int J Mol Sci,2017,18(6): 1321.[9]Dos Santos E,Pecquery R,de Mazancourt P,et al.Adiponectin and reproduction[J].Vitam Horm,2012,90:187-209.[10]Michalakis KG,Segars JH.The role of adiponectin in reproduction: from polycystic ovary syndrome to assisted reproduction[J].Fertil Steril,2010,94(6):1949-57.[11]Zhang N,Hao C,Liu X,et al.A potential determinant role of adiponectin and receptors for the early embryo development in PCOS patients with obesity hinted by quantitative profiling[J].Gynecol Endocrinol,2017,33(2):113-8.[12]Bannigida DM,Nayak SB,R V.Serum visfatin and adiponectin-markers in women with polycystic ovarian syndrome[J].Archives of Physiology&Biochemistry,2020,126(4):283-6.[13]Benrick A,Chanclón B,Micallef P,et al.Adiponectin protects against development of metabolic disturbances in a PCOS mouse model[J].Proceedings of the National Academy of Sciences,2017, 114(34):E7187-96.[14]Wagner N,Wagner KD.The role of PPARs in disease[J].Cells, 2020,9(11):2367.[15]Yang J,Chen L,Zhang X,et al.PPARs and female reproduction: evidence from genetically manipulated mice[J].PPAR Res,2011,31(23):4692-705.[16]尹天晓,温菁,魏巍等.小檗碱对PCOS大鼠子宫内膜PPARs的表达及内分泌代谢的影响[J].中国医学创新,2019,16(7):1-6.[17]Froment P,Gizard F,Defever D,et al.Peroxisome proliferator-activated receptors in reproductive tissues:from gametogenesis toparturition[J].The Journal of endocrinology,2006,189(2):199-209.[18]李维宏,牟晓玲,漆洪波.脂联素通过p38MAPK信号通路影响URSA患者外周血AdipoR1和免疫细胞平衡[J].免疫学杂志,2018,34(5):436-42.[19]Silber M,Miller I,Bar-Joseph H,et al.Elucidating the role of pigment epithelium-derived factor(PEDF)in metabolic PCOSmodels[J].J Endocrinol,2020,244(2):297-308.[20]Fauser BC,Tarlatzis BC,Rebar RW,et al.Consensus on women's health aspects of polycystic ovary syndrome(PCOS):theamsterdam ESHRE/ASRM-Sponsored3rd PCOS consensusworkshop group[J].Fertil Steril,2012,97(1):28-38.[21]Oróstica L,Rosas C,Plaza-Parrochia F,et al.Altered steroid metabolism and insulin signaling in PCOS endometria:Impactin tissue function[J].Bentham Science,2016,22(36):5614-24.[22]Wang J,Wu D,Guo H,et al.Hyperandrogenemia and insulin resistance:The chief culprit of polycystic ovary syndrome[J].LifeSci,2019,11(1):236:116940.[23]Lessey BA,Young SL.What exactly is endometrial receptivity[J].Fertil Steril,2019,111(4):611-7.[24]Bui AH,Timmons DB,Young SL.Evaluation of endometrial receptivity and implantation failure[J].Curr Opin Obstet Gynecol,2022,34(3):107-13.[25]韩华,薛改,李洁,等.宫腔粘连模型大鼠子宫内膜胞饮突发育和整合素β3表达[J].现代妇产科进展,2017,26(5):345-8.[26]Kara M,Ozcan SS,Aran T,et al.Evaluation of endometrial receptivity by measuring HOXA-10,HOXA-11,and leukemiainhibitory factor expression in patients with polycystic ovarysyndrome[J].Gynecol Minim Invasive Ther,2019,8(3):118-22.[27]Michalakis KG,Segars JH.The role of adiponectin in reproduction: from polycystic ovary syndrome to assisted reproduction[J].Fertility and Sterility,2010,94(6):1949-57.[28]Cermik D,Selam B,Taylor HS.Regulation of HOXA-10expression by testosteronein vitroand in the endometrium of patients withpolycystic ovary syndrome[J].J Clinical Endocrinol Metab,2003,88(1):238-43.[29]Cabrera-Cruz H,Orostica L,Plaza-Parrochia F,et al.The insulin-sensitizing mechanism of myo-inositol is associated with AMPKactivation and GLUT-4expression in human endometrial cellsexposed to a PCOS environment[J].Am J Physiol EndocrinolMetab,2020,318(2):E237-48.[30]Zhang W,Zuo M,Lu J,et al.Adiponectin reduces embryonic loss rate and ameliorates trophoblast apoptosis in early pregnancy ofmice with polycystic ovary syndrome by affecting the AMPK/PI3K/Akt/FoxO3a signaling pathway[J].Reprod Sci,2020,27(12):2232-41.(编辑:吴锦雅) J South Med Univ,2024,44(2):298-307··307动脉粥样硬化(AS )是泛指动脉壁增厚、失去弹性、硬化性的疾病,以脂质沉积和内膜纤维化为特征,所导致的冠状动脉粥样硬化性心脏病(CHD )、心肌梗死、脑梗死等心脑血管疾病严重威胁人类生命健康[1,2]。
doi:10.1152/ajprenal.00003.2003 285:336-347, 2003. First published Apr 29, 2003;Am J Physiol Renal PhysiolPierre-Yves Martin and Eric Féraille Mauro Bustamante, Frank Roger, Marie-Luce Bochaton-Piallat, Giulio Gabbiani,kidney MTAL kinase-dependent actin cytoskeleton remodeling in rat Regulatory volume increase is associated with p38 You might find this additional information useful...52 articles, 38 of which you can access free at: This article cites http://ajprenal.physiology.org/cgi/content/full/285/2/F336#BIBL
1 other HighWire hosted article: This article has been cited by
[PDF] [Full Text] [Abstract], January 1, 2006; 290 (1): C134-C142. Am J Physiol Cell PhysiolK. B. E. Gagnon, R. England and E. Delpire kinases: Ste20-related proline-alanine-rich kinase and WNK4Volume sensitivity of cation-Cl- cotransporters is modulated by the interaction of two
including high-resolution figures, can be found at: Updated information and services http://ajprenal.physiology.org/cgi/content/full/285/2/F336
can be found at: AJP - Renal Physiologyabout Additional material and information http://www.the-aps.org/publications/ajprenal
This information is current as of October 20, 2006 .
http://www.the-aps.org/.American Physiological Society. ISSN: 0363-6127, ESSN: 1522-1466. Visit our website at (monthly) by the American Physiological Society, 9650 Rockville Pike, Bethesda MD 20814-3991. Copyright © 2005 by therespective cells and vasculature, as well as to the control of body fluid volume and composition. It is published 12 times a year publishes original manuscripts on a broad range of subjects relating to the kidney, urinary tract, and theirAJP - Renal Physiology on October 20, 2006
ajprenal.physiology.org
Downloaded from Regulatoryvolumeincreaseisassociatedwithp38kinase-dependentactincytoskeletonremodelinginratkidneyMTAL
MauroBustamante,1FrankRoger,1Marie-LuceBochaton-Piallat,2GiulioGabbiani,2Pierre-YvesMartin,1andEricFe´raille11DivisiondeNe´phrologie,FondationpourRecherchesMe´dicales,
and2De´partementdePathologie,CentreMe´dicalUniversitaire,CH-1211Gene`ve4,Switzerland
Submitted7January2003;acceptedinfinalform22April2003
Bustamante,Mauro,FrankRoger,Marie-LuceBo-chaton-Piallat,GiulioGabbiani,Pierre-YvesMartin,andEricFe´raille.Regulatoryvolumeincreaseisassociatedwithp38kinase-dependentactincytoskeletonremodelinginratkidneyMTAL.AmJPhysiolRenalPhysiol285:F336–F347,2003.FirstpublishedApril29,2003;10.1152/ajprenal.00003.2003.—Thekidneymedullaisphysiologicallyexposedtovariationsinextracellularosmolality.Inresponsetohypertoniccellshrinkage,cellsoftheratkidneymedullarythickascendinglimbofHenle’sloopundergop38kinase-dependentregulatoryvolumeincrease(RVI).Inthepresentstudy,weinvestigatedtheroleofactincytoskeletonreorga-nizationinthisprocess.AdditionofhyperosmoticNaClorsucrose,whichactivatesMAPkinasesandreducescellularvolume,inducedasustainedactinpolymerizationoccurringafter10minandconcurrentlywithRVI.Incontrast,hyper-osmoticurea,whichdoesnotmodifyMAPkinaseactivityandcellularvolume,didnotinducesustainedactinpolymeriza-tion.FluorescencemicroscopyrevealedthathyperosmoticNaClandsucrose,butnoturea,inducedtheredistributionofF-actinfromadensecorticalringtoadiffusenetworkofactinbundles.StabilizationofactinfilamentsbyjasplakinolideandinhibitionofthegenerationofnewactinfilamentsbyswinholideApreventedRVI,whereasdepolymerizationofactinfilamentsbylatrunculinBattenuatedcellshrinkageandenhancedRVI.Theseactin-interferingdrugsdidnotalterextracellularregulatedkinaseandp38kinaseactiva-tionunderhypertonicconditions.SimilartoswinholideA,inhibitingp38kinasewithSB-203580abolishedsustainedactinpolymerization,actinredistribution,anddecreasedRVIefficacy.Wethereforeproposethatinratkidneythemedul-larythickascendinglimbofHenle’sloopexposedtoextracel-lularhypertonicity,p38kinaseactivationinducesdepoly-merizationoftheF-actincorticalringandpolymerizationofadensediffuseF-actinnetworkthatbothcontributetoin-creaseRVIefficacy.
mitogen-activatedproteinkinase;osmolarity;kidneyme-dulla;cellvolume
DURINGDIURESISANDantidiuresis,thekidneymedullaofvertebratesisexposedtolargefluctuationsinintersti-tialosmolality(23),whichchallengescellvolumecon-stancy.Underantidiuresis,thecountercurrentconcen-trationmechanisminitiatedbyactiveNaClreabsorp-tionbythemedullarythickascendinglimbofHenle’s
loop(MTAL)leadstoNaClandureaaccumulationintherenalmedulla.MTALcellsarethereforeofspecialinterestbecausetheyhavedevelopedadaptivemecha-nismstosurviveandfunctioninahypertonicenviron-ment.Thehighinterstitialosmolalityinduceschangesintheactivityofsolutetransportersandenzymesinvolvedinsoluteaccumulationandintheexpressionofgenesencodingenzymesrequiredforsolutesynthe-sis,stressresistance,andyeastcellwallstructure(34).Aftercellshrinkageinresponsetoextracellularhyper-tonicity,MTALcellsprogressivelyrecovertheirinitialvolumethroughregulatoryvolumeincrease(RVI).Thisprocessoccurswithinminutesandismediatedbystimulationofiontransportersthatincreaseintracel-lularionicconcentrations,whichdrivewaterinfluxandrestoreinitialcellularvolume(34,43).Therapidincreaseinionicconcentrationsisfollowedbyaslowaccumulationofintracellularcompatibleosmolytes,suchassorbitol,myoinositol,taurine,betaine,andglycerophosphocholine,allowingrecoveryofnormalionicconcentrations(7).Thisprocessisalong-termmechanismoccurringwithinhoursordaysandcoun-teractsincreasedextracellularosmoticpressure.HypertonicconditionsactivateMAPK,whichisanimportantsignaltransducerlinkingsignalsfromthecellsurfacetothenucleus.MAPKsareserine/threo-ninekinasesactivatedbyacascadeofkinasesinvolv-ingtwoupstreamkinases,MAPKKKandMAPKK(48).Inmammaliancells,MAPKsaredividedintothreefamilies,eachrespondingtodistinctextracellu-larstimuli:ERK1and2,JNK(alsoknownasstress-activatedproteinkinases1),andp38kinases(orstress-activatedproteinkinase2).Inourpreviousstudy,weshowedthatcellshrinkage,ratherthanintracellularhypertonicity,triggerstheactivationofERKandp38kinaseinratMTALs(35).MAPKacti-vationlevelsweredependentontheosmolyteusedtoincreaseextracellularosmolality.HyperosmoticNaClinducedcellshrinkageandactivatedERKandp38kinasebutnotJNK.Incomparison,hyperosmoticsu-croseinducedevengreatercellshrinkageandstrongeractivationofERKandp38kinaseandalsoactivatedJNKbuttoalesserextent.Bycontrast,hyperosmotic