
第40卷 第5期 海 洋 与 湖 沼
Vol.40, No.5 2009年
9月
OCEANOLOGIA ET LIMNOLOGIA SINICA
Sep., 2009
* 国家“863”项目资助,2007AA09Z433号; 福建省科技重大专项前期项目资助,2005NZ1011号。杨秋玲,E-mail :
yangql0712@https://www.doczj.com/doc/a311105593.html,
1) ITIS (Integrated Taxonomic Information System), 2001. https://www.doczj.com/doc/a311105593.html, ① 通讯作者: 林祥志,副研究员,E-mail :lxz8848@https://www.doczj.com/doc/a311105593.html,
收稿日期: 2008-08-16, 收修改稿日期: 2008-12-19
东南沿海小孔蛸(Cistopus sp.)线粒体序列的
比较研究*
杨秋玲1, 2 林祥志1①
郑小东3 苏永全2
(1. 国家海洋局第三海洋研究所 厦门 361005; 2. 厦门大学海洋与环境学院 厦门 361005;
3. 中国海洋大学水产学院 青岛 266003)
提要 采用线粒体DNA 16S rRNA 和CO Ⅰ基因序列对福建、浙江沿海分布的5个小孔蛸(Cistopus sp.)群体进行了遗传多样性研究。结果表明, 序列总长度分别为497—501bp (16S rRNA)和652bp (CO Ⅰ); 小孔蛸5个群体的序列碱基组成均显示出较高的A+T 含量(16S rRNA 基因72.1%, CO Ⅰ基因65.9%)。16S rRNA 基因片段在种内和群体间个体差异均较小, 5个群体共检测到8个变异位点(其中7个插入/缺失位点); CO Ⅰ基因序列不存在插入/缺失位点, 群体内个体间变异位点数为21个。系统进化树表明, 宁德群体和象山群体与其它三个群体的亲缘关系较远, 但存在基因交流。线粒体16S rRNA 和CO Ⅰ基因在群体间无显著多态性分布。
关键词 小孔蛸, 线粒体DNA, 16S rRNA 基因, CO Ⅰ基因 中图分类号 Q959.216
头足类(cephalopods)遍布世界各大洋, 有记载的生活种有700多种(董正之, 1987; 郑玉水等, 1987; ITIS, 20011)), 由于过度捕捞, 一些重要经济物种, 特别是近岸种类的产量在过去20年来急剧下降。国际市场对头足类的需求却有增无减, 供求关系的严重失衡使得头足类的养殖作为资源恢复的重要手段受到广泛关注, 蛸类则成为头足类中最受关注的养殖对象之一(林祥志等, 2006)。小孔蛸(Cistopus sp.)隶属蛸科(Octopodidae)、小孔蛸属(Cistopus ), 是一种我国东南沿海底栖型重要经济头足类, 俗称“章鱼”、“望潮”, 其肉质细嫩、营养丰富、味道鲜美, 是东南沿海地区传统名点小吃, 也深受港澳台同胞和海外侨胞欢迎。由于其风味独特, 市场上供不应求, 加上该种经济价值高, 生长快, 养殖周期短(通常为8—10个月), 当年即可上市, 因此, 是理想的增养殖品种。
目前, 无序采捕使得小孔蛸资源量衰减严重, 增
养殖研究正逐步开展起来。不过我国沿海现存群体仍为野生型, 本文通过线粒体(mtDNA)16S rRNA 和CO Ⅰ基因序列测定和比较分析, 研究了福建、浙江沿海分布的5个小孔蛸群体的遗传结构和分化状况, 以期能为该种的种质鉴定、资源保护、养殖与育种管理提供理论依据。
1 材料与方法
1.1 材料来源
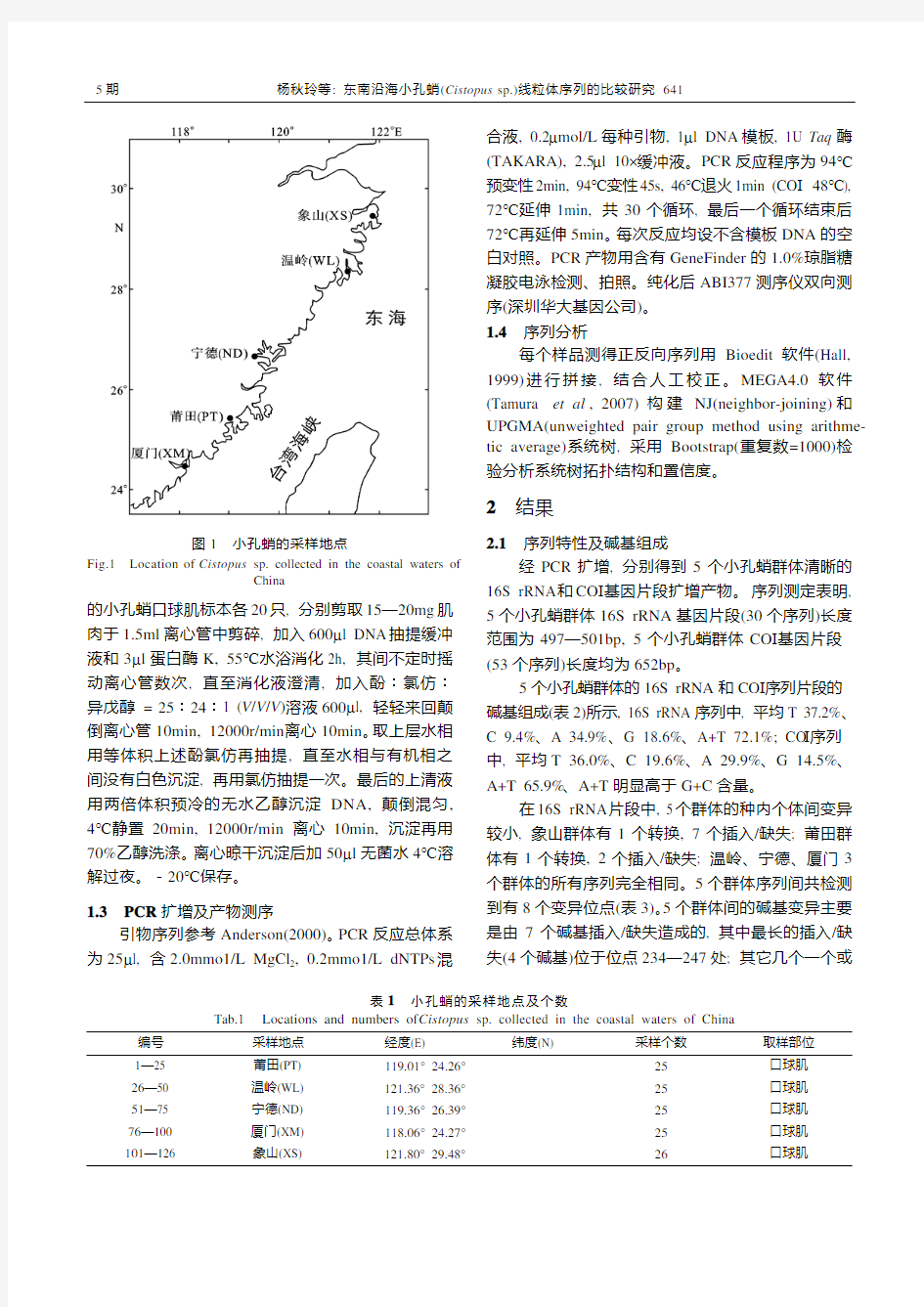
实验用小孔蛸样品于2007年6—11月分别采自浙江象山(XS)和温岭(WL), 福建宁德(ND)、莆田(PT)和厦门(XM)(图1, 表1)。所有个体(共126个)活体取样, 取口球肌组织浸泡于95%乙醇中, -20℃保存。 1.2 总DNA 提取
参考《分子克隆实验指南》(Sambrook et al , 1989)的酚/氯仿抽提法, 略有修改。随机选取上述5个群体
5期杨秋玲等:东南沿海小孔蛸(Cistopus sp.)线粒体序列的比较研究
641
图1 小孔蛸的采样地点
Fig.1 Location of Cistopus sp. collected in the coastal waters of
China
的小孔蛸口球肌标本各20只, 分别剪取15—20mg肌肉于1.5ml离心管中剪碎, 加入600μl DNA抽提缓冲液和3μl蛋白酶K, 55℃水浴消化2h, 其间不定时摇动离心管数次, 直至消化液澄清, ︰︰
加入酚氯仿
异戊醇 = 25︰24︰1 (V/V/V)溶液600μl, 轻轻来回颠倒离心管10min, 12000r/min离心10min。取上层水相用等体积上述酚氯仿再抽提, 直至水相与有机相之间没有白色沉淀, 再用氯仿抽提一次。最后的上清液用两倍体积预冷的无水乙醇沉淀DNA, 颠倒混匀, 4℃静置20min, 12000r/min 离心10min, 沉淀再用70%乙醇洗涤。离心晾干沉淀后加50μl无菌水4℃溶解过夜。-20℃保存。
1.3PCR扩增及产物测序
引物序列参考Anderson(2000)。PCR反应总体系为25μl, 含2.0mmo1/L MgCl2, 0.2mmo1/L dNTPs混合液, 0.2μmol/L每种引物, 1μl DNA模板, 1U Taq酶(TAKARA), 2.5μl 10×缓冲液。PCR反应程序为94℃预变性2min, 94℃变性45s, 46℃退火1min (COⅠ48℃), 72℃延伸1min, 共30个循环, 最后一个循环结束后72℃再延伸5min。每次反应均设不含模板DNA的空白对照。PCR产物用含有GeneFinder的1.0%琼脂糖凝胶电泳检测、拍照。纯化后ABI377测序仪双向测序(深圳华大基因公司)。
1.4序列分析
每个样品测得正反向序列用Bioedit软件(Hall, 1999)进行拼接, 结合人工校正。MEGA4.0软件(Tamura et al, 2007)构建NJ(neighbor-joining)和UPGMA(unweighted pair group method using arithme-tic average)系统树, 采用Bootstrap(重复数=1000)检验分析系统树拓扑结构和置信度。
2结果
2.1序列特性及碱基组成
经PCR扩增, 分别得到5个小孔蛸群体清晰的16S rRNA和COⅠ基因片段扩增产物。序列测定表明, 5个小孔蛸群体16S rRNA基因片段(30个序列)长度范围为497—501bp, 5个小孔蛸群体COⅠ基因片段(53个序列)长度均为652bp。
5个小孔蛸群体的16S rRNA和COⅠ序列片段的碱基组成(表2)所示, 16S rRNA序列中, 平均T 37.2%、C 9.4%、A 34.9%、G 18.6%、A+T 72.1%; COⅠ序列中, 平均T 36.0%、C 19.6%、A 29.9%、G 14.5%、A+T 65.9%、A+T明显高于G+C含量。
在16S rRNA片段中, 5个群体的种内个体间变异较小, 象山群体有1个转换, 7个插入/缺失; 莆田群体有1个转换, 2个插入/缺失; 温岭、宁德、厦门3个群体的所有序列完全相同。5个群体序列间共检测到有8个变异位点(表3)。5个群体间的碱基变异主要是由7个碱基插入/缺失造成的, 其中最长的插入/缺失(4个碱基)位于位点234—247处; 其它几个一个或
表1小孔蛸的采样地点及个数
Tab.1 Locations and numbers of Cistopus sp. collected in the coastal waters of China 编号采样地点经度(E) 纬度(N) 采样个数取样部位1—25 莆田(PT) 119.01° 24.26° 25 口球肌26—50 温岭(WL) 121.36° 28.36° 25 口球肌51—75 宁德(ND) 119.36° 26.39° 25 口球肌76—100 厦门(XM) 118.06° 24.27° 25 口球肌101—126 象山(XS) 121.80° 29.48° 26 口球肌
642 海洋与湖沼40卷
表25个群体小孔蛸16S rRNA和COⅠ基因片段的碱基组成(%)
Tab.2 The base composition (%) of 16S rRNA and COⅠgene fragments of five stocks of Cistopus sp.
16S rRNA片段COⅠ片段
群体
T C A G A+T T C A G A+T XS 37.32 9.46 34.68 18.54 72.0 36.19 19.39 29.92 14.43 66.11 WL 37.1 9.4 34.9 18.6 72.0 35.88 19.62 29.9 14.6 65.78 ND 37.1 9.4 34.9 18.6 72.0 35.98 19.52 29.88 14.62 65.86 PT 37.13 9.4 34.9 18.57 72.03 35.91 19.6 29.94 14.55 65.85 XM 37.1 9.4 34.9 18.6 72.0 35.9 19.6 29.92 14.58 65.82 平均37.2 9.4 34.9 18.6 72.1 36.0 19.6 29.9 14.5 65.9
两个碱基的插入/缺失散布于序列位点280—320之间的3个地方, 在466bp处有1个碱基转换(G→A)。
与16S rRNA片段相比, COⅠ序列不存在任何插入/缺失位点, 群体内个体间变异位点数为21个。象山群体有8个转换, 1个颠换; 温岭、厦门两个群体各有1个转换; 宁德群体有2个转换; 莆田群体有10个转换。群体间共检测到14个变异位点(表4), 有13个转换, 1个颠换。2.2系统树和单倍型
2.2.1系统树小孔蛸5个群体的NJ(neighbor- joining)和UPGMA系统树通过MEGA4.0软件构建, 采用Bootstrap(重复数=1000)检验分子系统树各分支的置信度。两种系统树的拓扑图总体趋势相似, 16S rRNA的系统树较为简单(图2), 30个样品聚成两支:一支为象山群体的3个样品和莆田群体1个样品聚类而成; 另一支为其它所有26个样品聚类而成(其中厦
表3 16S rRNA基因4种单倍型的变异位点和每种单倍型在各个群体中的个体数
Tab.3 Variable nucleotide positions in part of the 16S rRNA gene region of four haplotypes and the number of individual of each
haplotype found in each stock
变异位点(bp) 个体数(个)
单倍型
234 235 236 237 281 290 320 466 XS WL ND PT XM 合计
H1 A A T A A A -0 0 6 6 5 7 24 H2 . . . . . -T A 0 0 0 1 0 1 H3 . . . . -. . . 2 0 0 0 0 2 H4 ----. -T A 3 0 0 0 0 3 注:“-”表示碱基缺失; “.”表示碱基相同
表4 COⅠ基因8种单倍型的变异位点和每种单倍型在各个群体中的个体数
Tab.4 Variable nucleotide positions in the COⅠgene region of eight haplotypes and the number of individual for each haplotype
found in every stock
变异位点(bp) 个体数(个)
单倍型
53 71 77 165 204 211 293 361385400448508542622XS WL ND PT XM合计
H1 T A T T G A G G T C C C G C 5 9 0 8 9 31
H2 C . . . . . . . . . . . . . 0 1 0 0 0 1 H3 . G . . . . . . . . . . . . 0 0 2 0 0 2 H4 . . C C A T . A C T. T. T 1 0 0 1 0 2 H5 . . . C . T . A. T. T. T7 0 0 0 0 7 H6 . . . . . . A . . . . . . . 0 0 0 0 1 1 H7 . . . . . . . . . . T. . . 0 0 8 0 0 8 H8 . . . . . . . . . . . . A. 0 0 0 1 0 1 注:“.”表示碱基相同
5期杨秋玲等:东南沿海小孔蛸(Cistopus sp.)线粒体序列的比较研究 643
门、温岭、宁德三个群体个体全都聚为一支)。群体间的聚类可以看出:厦门、温岭、宁德三个群体所有个体完全聚在一起, 莆田群体和象山群体与其它三个群体交叉, 从系统树可以看出5个群体都有交叉, 不能很明显的分离出某个群体。COⅠ的系统树较16S rRNA的复杂(图3)。从图3中可以看出, 宁德群体所有个体先聚为独立的两支(8个个体聚为一支, 另两个聚为另一支), 温岭、莆田、厦门三个群体先聚类为一支, 再与宁德所有个体聚类, 并得到较高的Bootstrap支持(99%)。莆田群体的09号样与象山群体的120先聚类, 再与象山群体的大部分个体聚为一支, 然后再与另一大支聚在一起。
2.2.2单倍型
16S rRNA:共有4个单倍型, 单倍型H1为4个群体共有(象山群体除外), 且为最主要的单倍型, 出现的频率为80%(24/30)。厦门、温岭、宁德三个群体仅共有单倍型H1; 莆田、象山两个群体都有自己独有的单倍型, 象山群体具2个特有单倍型(单倍型H3、H4); 莆田群体除拥有共有单倍型H1外, 还有自己特有的单倍型H2。
COⅠ:共有8种单倍型, 单倍型H1为4个群体共有(宁德群体除外), 且为最主要的单倍型, 出现的频率是56.6% (30/53)。每个群体都有自己独有的单倍型, 单倍型H2、H3、H5、H6、H7、H8都为群体独有单倍型(其中单倍型H7和H3都为宁德群体独有; H8-莆田群体; H2-温岭群体; H6-厦门群体; H5-象山群体), 单倍型H4为莆田、象山两个群体共有。在5个群体中, 象山群体和莆田群体拥有最多单倍型, 都为3种, 且有两种是共有(单倍型H1、H4); 其它三个群体都有2种单倍型。其中宁德群体只拥有自己独有的两种单倍型, 没有单倍型H1; 温岭群体和厦门群体除有共有单倍型H1外, 还有自己独有的一个单倍型。
3讨论
本文得到的小孔蛸16S rRNA和COⅠ基因序列中A+T含量相差不大, 分别为72.1%和65.9%, 都明显高于G+C含量, 这一结果与其他研究者在头足类、双壳类等的16S rRNA和COⅠ基因中观察到的结果一致(Anderson, 2000; Zheng et al, 2003; 孔晓瑜等, 2001), 即较高的AT含量, 是目前已检测的无脊椎动物线粒体DNA序列中较为普遍的现象(杨建敏等, 2003)。
一般认为, 在线粒体基因组中, 12S rRNA和16S rRNA基因进化速率低, 比较保守, 而COⅠ基因为变异性较大的区域, 在软体动物(贝类)中检测到较大的
图2 基于16S rRNA基因片段序列构建的小孔蛸5个群体的NJ和UPGMA系统发生树Fig.2 NJ and UPGMA phylogenetic trees based on 16S rRNA gene sequences of the 5 stocks of Cistopus sp. A: NJ; B: UPGMA。群体后的数字表示样品编号(例如ND-42表示编号为42号的样品, 采集地点为宁德)
644 海 洋 与 湖 沼 40卷
图3 基于CO Ⅰ基因片段序列构建的小孔蛸5个群体的NJ 和UPGMA 系统发生树
Fig.3 NJ and UPGMA phylogenetic trees based on CO Ⅰ gene sequences of five stocks of Cistopus sp. A: NJ; B: UPGMA 。群体后的数字表示样品编号(例如ND-42表示编号为42号的样品, 采集地点为宁德)
变异(苏天凤等, 2005; 陈丽梅等, 2005; 程汉良等, 2007)。从本实验的结果可以看出, 在长度为497—501bp 的16S rRNA 基因片段中, 小孔蛸5个群体存在8个碱基变异, 仅有4种单倍型; 而在长度为652bp 的CO Ⅰ基因片段中, 存在14个碱基变异, 具有8种单倍型, 且每个群体的单倍型多样性多于16S rRNA 。本文中得到5个群体小孔蛸的16S rRNA 和CO Ⅰ基因序列中, CO Ⅰ基因表现出的群体内和群体间趋异都要高于16S rRNA 基因, 说明在小孔蛸的5个群体中, CO Ⅰ基因的进化速率与16SrRNA 基因相比要快, 16S rRNA 基因则相对保守一些。小孔蛸线粒体16S rRNA 和CO Ⅰ基因在不同群体间其多态性并
无明显差异, 不能很明显的将不同地理群体分开。Zheng 等(2003)采用线粒体16S rRNA 序列分析了我国沿海日本无针乌贼(又称曼氏无针乌贼)的群体结构; Oosthuizen 等(2004)采用mtDNA CO ⅠII 序列分析了南非沿海的真蛸群体; Aoki 等(2008)采用线粒体非编码区序列(Non-coding Region)研究了日本海域莱氏拟乌贼群体遗传结构, 均发现了较低的遗传多样性。头足类作为游泳型贝类, 其线粒体DNA 序列可能并不适合做种以下阶元的遗传多样性分析, 更适合当做条形码DNA 用于属、科以上阶元的系统发生学研究(Strugnell et al, 2008)。
自Shaw(1997)首次采用高度多态微卫星位点对
5期杨秋玲等:东南沿海小孔蛸(Cistopus sp.)线粒体序列的比较研究 645
东北大西洋的福氏枪乌贼群体进行了报道以来, 微卫星DNA(msDNA)技术为研究头足类群体结构和遗传多样性提供了一个新途径的应用, 尤其在采用其它方法(如同工酶电泳、线粒体DNA序列等)检测的低变异的种类[如真蛸(Murphy et al, 2002)、金乌贼(Zheng et al, 2009)]。有关小孔蛸种群的msDNA研究正在进行中。
由16S rRNA和COⅠ基因片段得到的系统树所表现的趋势相似。基于两段基因片段的系统树(NJ和UPGMA)聚类结果可看出宁德和象山两个群体通过COⅠ可以区别于其它三个群体。16S rRNA和COⅠ分析结果显示厦门、温岭、莆田3个群体几乎都完全交叉, 表明这三个群体由于地理位置邻近可能存在一定的基因交流, 而宁德为一个封闭的港湾, 与外海的基因交流受到限制, 而象山距离其它四个采样点较远。对于莆田群体的09号个体与象山群体的亲缘关系很近的原因还有待进一步的研究。
参考文献
孔晓瑜, 刘亚军, 喻子牛等, 2001. 栉孔扇贝和海湾扇贝线粒体DNA 16S rRNA基因片段序列研究. 见: 中国贝类学会主编. 贝类学论文集第IX辑. 北京: 海洋出版社, 9: 59—
63
苏天凤, 江世贵, 周发林等, 2005. 近江牡蛎16S rRNA基因片段序列变异分析. 高技术通讯, 15(2): 100—103
杨建敏, 郑小东, 王如才等, 2003. 3种鲍16S rRNA基因片段序列的初步研究. 青岛海洋大学学报, 33(1): 36—40
陈丽梅, 孔晓瑜, 喻子牛等, 2005. 3种蛏类线粒体16S rRNA 和COⅠ基因片段的序列比较及其系统学初步研究. 海洋科学, 29(8): 27—32
林祥志, 郑小东, 苏永全等, 2006. 蛸类养殖生物学研究现状及展望. 厦门大学学报(自然科学版), 45(2): 213—218
郑玉水, 李复雪, 1987. 东海头足类的研究. 福建水产, 3: 15—
21
董正之, 1987. 中国动物志——软体动物门: 头足纲. 北京: 科学出版社, 1—20
程汉良, 夏德全, 吴婷婷等, 2007. 6种帘蛤科贝类及4个地理
种群文蛤线粒体COⅠ基因片段序列分析. 海洋学报, 29(5): 109—116
Anderson F E, 2000. Phylogeny and historical biogeography of the loliginid squids (Mollusca: Cephalopoda) based on mi-
tochondrial DNA sequence data. Molecular Biology and Evolution, 15(2): 191—214
Aoki M, Imai H, Naruse T et al, 2008. Low Genetic Diversity of Oval Squid, Sepioteuthis cf. lessoniana (Cephalopoda: Lo-
liginidae), in Japanese Waters Inferred from a Mitochondrial DNA Non-coding Region. Pacific Science, 62: 403—411
Hall T A, 1999. BioEdit: a user-friendly biological sequence alignment editor and analysis program for Windows 95/98/NT. Nucleic Acids Symposium Series, 41: 95—98 Murphy J M, Balguerías E, Key L N et al, 2002. Microsatellite DNA markers discriminate between two Octopus vulgaris
(Cephalopoda: Octopoda) fisheries along the northwest Af-
rican coast. Bulletin of Marine Science, 71: 545—553 Oosthuizen A, Jiwaji M, Shaw P, 2004. Genetic analysis of the Octopus vulgaris population on the coast of South Africa.
South African Journal of Science, 100: 603—607
Sambrook J, Fitch E, Maniatis T, 1989. Molecular Cloning: A Laboratory Manual. 2nd eds. New York: Cold Spring Harbor Laboratory Press, 463—468
Shaw P W, 1997. Polymorphic microsatellite DNA markers in the veined squid Loligo forbesi. Molecular Ecology, 6: 297—
298
Strugnell J M, Collins M A, Allcock A L, 2008. Molecular evolu-tionary relationships of the octopodid genus Thaumeledone
(Cephalopoda: Octopodidae) from the Southern Ocean. Ant-
arctic Science, 20: 245—251
Tamura K, Dudley J, Nei M et al, 2007. MEGA4: Molecular evolutionary genetics analysis (MEGA) software Version 4.0.
Molecular Biology and Evolution, 24(8): 1596—1599
Zheng X D, Ikeda M, Kong L F et al, 2009. Genetic diversity and population structure of the golden cuttlefish, Sepia esculenta
indicated by microsatellite DNA variations. Marine Ecology, doi:10.1111/j.1439-0485.2009.00294.x
Zheng X D, Wang R C, Xiao S et al, 2003. Genetic diversity in populations of Sepiella maindroni using 16S rRNA gene sequence analysis. High Technology Letters, 9(1): 1—5
646 海洋与湖沼40卷GENETIC DIVERSITY IN POPULATIONS OF CISTOPUS SP. BASED ON THE
SEQUENCES OF MITOCHONDRIAL 16S rRNA AND COⅠ GENE FRAGMENTS
YANG Qiu-Ling1, 2, LIN Xiang-Zhi1, ZHENG Xiao-Dong3, SU Yong-Quan2
(1. Third Institute of Oceanography, State Oceanic Administration, Xiamen, 361005; 2. College of Oceanography and Environmental
Science, Xiamen University, Xiamen, 361005; 3. Fisheries College, Ocean University of China, Qingdao, 266003)
Abstract Sequences of 16S rRNA and COⅠgene fragments in mtDNA of Cistopus sp. were studied. The samples were collected from five populations in the East China Sea. The sequences were compared by PCR. Nucleotide sequences of 497—501bp for 16S rRNA gene and 652bp for COⅠgene were obtained. A high percentage of A+T base composition in the two sequences was observed. The alignment indicated that the variation level of intra-stocks was very low in both genes. Eight variable sites were detected for 16S rRNA gene fragment (of which seven were indels) across the five stocks.
21 polymorphic sites were detected in COⅠgene fragments among the five stocks, with no indels. The NJ and UPGMA phylogenetic trees showed that no remarkable genetic difference was observed among those populations.
Key words Cistopus sp., Mitochondrial DNA, 16S rRNA gene, COⅠ gene
自噬(Autophagy)及其研究方法概述汉恒Th物技术服务手册
目录 1概念 2自噬的过程 3自噬的特性 4自噬过程的调控 5自噬与肿瘤的关系 6自噬的研究方法概述7汉恒自噬研究特色服务
自噬研究相关产品及服务 1.病毒工具(独家推出) mRFP-GFP-LC3腺病毒系统,可高效感染目的细胞,表达mRFP-GFP-LC3,感染后细胞可在荧光显微镜下实时观察自噬发Th过程(具体内容后面有介绍); 2.自噬相关服务 汉恒Th物可提供自噬研究整体科研服务,若您的时间紧张或是对实验有所顾忌,我们可以为您代劳部分实验内容; 3.自噬研究相关试剂 汉恒Th物可以根据您的实验需求为您提供最实用的试剂产品,让你用的放心,省心!
产品厂商规格A14292,Premo自噬TB/GFP TR-FRET invitrogen6000Tests LC3B抗体试剂盒 pllabs0.1mg Anti-MAP1A/LC3A/B自噬微管相关蛋白 轻链3抗体 pllabs0.1mg Anti-MAP1LC3A(microtubule- associated protein1light chain 3)自噬微管相关蛋白轻链3抗体 Invitrogen1mg/1ml 兔抗人、大、小APG4B细胞自噬相关 抗体\Anti-APG4B/AUTL1 BD1mg/1ml 自噬微管相关蛋白轻链3抗体\ Anti-MAP1LC3A Anti-SQSTM1/p62antibody abcam Sigma1g 溶酶体抑制剂Hydroxychloroquine (羟氯喹) mTOR抑制剂rapamycin Sigma20mM 自噬抑制剂3-Methyladenine(3-MA)Sigma100mg
自噬研究鼻祖的最终选择 细胞自噬是细胞应对恶劣环境的一种主动反应,就是将自身一部分动员出来,采用自吃的方式,作为能量物质来应对各种不利因素,细胞凋亡则是细胞整体的主动死亡方式,最近有研究发现细胞坏死也存在一种主动的方式被称为程序性坏死,细胞自噬也是一种程序性坏死的类型。细胞自噬(autophagy)是继细胞凋亡(apoptosis)后,近年来生命科学领域的又一热门研究方向。 比利时科学家克里斯汀·德迪夫主要的研究领与在生物化学与细胞生物学,上世纪50年代,他利用刚刚出现的细胞分级分离技术(通过超速离心来分离细胞成分),发现了溶酶体(lysosome)和过氧化物酶体(peroxisome),让人们对细胞内部结构有了更清楚的认识,极大推动了细胞生物学研究。1974年因“细胞的结构和功能组织方面的发现(for their discoveries concerning the structural and functional organization of the cell)”而与阿尔伯特·克劳德、乔治·埃米尔·帕拉德分享了诺贝尔生理学或医学奖。后来,随着研究的深入,德迪夫的兴趣逐渐转向细胞起源,例如内共生学说。 克里斯汀·德迪夫Christian de Duve在上世纪50年代通过电镜观察到自噬体(autophagosome)结构,并且在 1963 年溶酶体国际会议(CIBA Foundation Symposium on Lysosomes)上首先提出了“自噬”这种说法。因此克里斯汀·德迪夫被公认为自噬研究的鼻祖。目前根据发生过程分为三类:Macroautophagy,Microautophagy和
自噬(Autophagy)及其研究方法概述 一、背景 概念: 目前根据发生过程分为三类:Macroautophagy,Microautophagy和Chaperone-mediated autophagy CMA), 大自噬(Macroautophagy)即我们说的自噬(autophagy);微自噬(Microautophagy):是指溶酶体主动、直接吞噬胞浆成分的一种方式;分子伴侣介导的自噬(Chaperone-mediated autophagy,CMA):一些分子伴侣,如hsp70,能帮助未折叠蛋白转位入溶酶体。通常说的自噬泛指Macroautophagy. 自噬是细胞内的一种“自食(Self-eating)”的现象,凋亡是“自杀(Self-killing)”的现象,二者共用相同的刺激因素和调节蛋白,但是诱发阈值和门槛不同,如何转换和协调目前还不清楚. 自噬是指膜(目前来源还有争议,大部分表现为双层膜,有时多层或单层)包裹部分胞质和细胞内需降解的细胞器、蛋白质等形成自噬体(autophagosome),最后与溶酶体融合形成自噬溶酶体(autophagolysosome),降解其所包裹的内容物,以实现细胞稳态和细胞器的更新。自噬的步骤可以大概总结为下面四步: 步骤1:细胞接受自噬诱导信号后,在胞浆的某处形成一个小的类似“脂质体”样的膜结构,然后不断扩张,但它并不呈球形,而是扁平的,就像一个由2层脂双层组成的碗,可在电镜下观察到,被称为Phagophore,是自噬发生的铁证之一。 步骤2:Phagophore不断延伸,将胞浆中的任何成分,包括细胞器,全部
揽入“碗”中,然后“收口”,成为密闭的球状的autophagosome,即“自噬体”。电镜下观察到自噬体是自噬发生的铁证之二。有2个特征:一是双层膜,二是内含胞浆成分,如线粒体、内质网碎片等。 步骤3:自噬体形成后,可与细胞内吞的吞噬泡、吞饮泡和内体融合(这种情况不是必然要发生的)。 步骤4:自噬体与溶酶体融合形成autolysosome,期间自噬体的内膜被溶酶体酶降解,2者的内容物合为一体,自噬体中的“货物”也被降解,产物(氨基酸、脂肪酸等)被输送到胞浆中,供细胞重新利用,而残渣或被排出细胞外或滞留在胞浆中。 自噬的特性: 1)自噬是细胞消化掉自身的一部分,即self-eating,初一看似乎对细胞不利。事实上,细胞正常情况下很少发生自噬,除非有诱发因素的存在。这些诱发因素很多,也是研究的热门。既有来自于细胞外的(如外界中的营养成分、缺血缺氧、生长因子的浓度等),也有细胞内的(代谢压力、衰老或破损的细胞器、折叠错误或聚集的蛋白质等)。由于这些因素的经常性存在,因此,细胞保持了一种很低的、基础的自噬活性以维持自稳。 2)自噬过程很快,被诱导后8min即可观察到自噬体(autophagosome)形成,2h后自噬溶酶体(autolysosome)基本降解消失。这有利于细胞快速适应恶劣环境。 3)自噬的可诱导特性:表现在2个方面,第一是自噬相关蛋白的快速合成,这是准备阶段。第二是自噬体的快速大量形成,这是执行阶段。 4)批量降解:这是与蛋白酶体降解途径的显着区别
自噬双标腺病毒(mRFP-GFP-LC3)使用指南 1 自噬双标腺病毒(mRFP-GFP-LC3)使用指南 背景: 自噬是细胞内的一种“自食(Self-eating )”的现象,凋亡是“自 杀(Self-killing )”的现象,二者共用相同的刺激因素和调节蛋白, 但是诱发阈值和门槛不同,如何转换和协调目前还不清楚. 自噬是指 膜(目前来源还有争议,大部分表现为双层膜,有时多层或单层)包 裹部分胞质和细胞内需降解的细胞器、蛋白质等形成自噬体,最后与 溶酶体融合形成自噬溶酶体,降解其所包裹的内容物,以实现细胞稳 态和细胞器的更新。目前文献对自噬过程进行观察和检测常用的策略 和手段有:通过western blot 检测LC3的剪切;通过电镜观测自噬体 的形成;在荧光显微镜下采用GFP (-RFP )-LC3等融合蛋白来示踪自 噬体形成以及降解。近几年对自噬流的研究日趋增多,针对于此我们 汉恒生物科技(上海)有限公司自主研发了用于实时监测自噬(流) 的mRFP-GFP-LC3腺病毒,mRFP 用于标记及追踪LC3,GFP 的减弱可指 示溶酶体与自噬小体的融合形成自噬溶酶体,即由于GFP 荧光蛋白对 酸性敏感,当自噬体与溶酶体融合后GFP 荧光发生淬灭,此时只能检 测到红色荧光。这种串联的荧光蛋白表达载体系统直观清晰的指示了 细胞自噬流的水平,是我们自噬研究尤其是自噬流研究不可或缺的利 器。
mRFP-GFP-LC3腺病毒的操作 收到病毒后的处理 (一)、腺病毒的储存 1、腺病毒采用冰袋运输。 (1)、收到病毒液后如未融化请置于-80℃冰箱,下次使用时再进 行分装; (2)、如客户收到时腺病毒已融化,请直接分装后置于-80℃冰箱 保存;若短期内用于实验,可分装部分于4℃保存(尽量一周内用完)。 2、尽量避免反复冻融,否则会降低病毒滴度(每次冻融会降低病 毒滴度10%)。建议不要在-20℃下长期保存。如果病毒储存时间 超过6个月,应该重新测定病毒滴度。 3、建议收到病毒产品后根据实验需求自行分装或购买经过分装的小 包装病毒产品(购买时请提出)。 (二)、腺病毒的稀释需要稀释病毒时,将病毒取出后置于冰上融解,使用培养目的细胞 用PBS 或培养基稀释到所需浓度后混匀分装后4℃保存,并尽快用于 实验(尽量一周内用完),动物实验建议使用注射用平衡液来稀释, 并尽快用完。 感染目的细胞 2
细胞自噬的研究进展 孙雅婧,郭青龙* 中国药科大学生理教研室,南京210009 细胞自噬(autophagy )是指细胞内受损、变性或衰老的蛋白质和细胞器被运输到溶酶体,溶酶体对其消化降解,以胞质内自噬体的出现为标志的细胞自我消化过程,以双层膜结构包裹部分胞质和细胞器的自噬体为判断指标。早在1962年,自噬现象的奠基人Ashford 和Porten 在人的肝细胞中用电子显微镜观察到了自噬现象。随着分子生物技术的发展,人们对自噬的形态特点和分子机制了解逐步深入。近年来对自噬的研究十分广泛,自噬是在体内普遍存在的过程,其在清除代谢废物进而回收能量为细胞正常运转提供能量的过程中发挥重要作用,因而对自噬的研究尤为重要。 1 细胞自噬的研究现状 1.1 自噬的过程 自噬的过程分为四个阶段(见图1)。 第一阶段:自噬诱导信号被细胞接受后,类“脂 质体”碗状结构即在胞浆某处形成小的膜结构,在电镜下观察到其不断扩张、呈非球形、扁平状双层膜的碗状结构,称为自噬前体(phagophore ),这种结构的电镜观察结果是指示自噬发生的金标准之一。 第二阶段:不断延伸的自噬前体,将胞浆中的若干成分(包括细胞器)收口包入,成为密闭的球状自噬体(autophagosome )。自噬体的电镜观察结果是指示自噬发生的金标准之一。自噬体的特征有两个:双层膜,内含诸如线粒体、内质网碎片等胞浆成分。 第三阶段:自噬体形成后,可能与细胞内吞的吞噬泡(phagocytic vacuole )、吞饮泡(pinosome )和内 体(endosome )融合(此阶段为非必需步骤)。 第四阶段:自噬体与溶酶体(lysosome )发生融合,形成自噬溶酶体(autolysosome )。期间溶酶体酶降解自噬体的内膜,使两者的内容物合为一体,自噬体中的包含物被降解,将产物诸如氨基酸、脂肪酸之类输送到胞浆中,重新利用供能,残渣则被排出细胞外或滞留于胞浆[1]。 1.2自噬的分类 根据细胞内底物运送到溶酶体腔方式的不同, 哺乳动物细胞自噬分为3种主要方式:巨自噬(macroautophagy )、微自噬(microautophagy )和分子伴侣介导自噬(chaperone-mediated autophagy ,简称 CMA )。巨自噬是最主要的自噬形式,在巨自噬中由 内质网来源的膜包绕待降解物,形成自噬体后与溶酶体融合并降解其内容物;然而在微自噬中,溶酶体膜直接内陷包裹长寿命蛋白等,并在溶酶体内降解,没有形成自噬小体的过程;分子伴侣介导自噬则为胞浆内蛋白结合到分子伴侣后转运到溶酶体腔中,被溶酶体酶消化。CMA 的底物是可溶蛋白分子,因此CMA 降解途径在清除蛋白质时有选择性,而前两者无明显的选择性[3]。 2自噬与凋亡 在多细胞生物体内,维持自身的稳态和内环境 的平衡,是保持复杂生物体系正常运转的重要条件。正常的细胞体系当中,有细胞的生长增殖必然 摘要本文综述了细胞自噬概念的研究现状、自噬与凋亡、自噬与肿瘤的关系,展望了自噬在抗癌药物介导的细胞死亡中发挥的重要作用以及自噬现象的临床意义。 关键词自噬;凋亡;肿瘤 中图分类号 Q25;R979.1文献标志码A 文章编号1673-7806(2012)03-236-04 作者简介 孙雅婧,女,硕士生E-mail:yj7782@https://www.doczj.com/doc/a311105593.html, 通讯作者郭青龙,男,教授,博士生导师,研究方向:肿瘤药理学 E-mail:anticancer_drug@https://www.doczj.com/doc/a311105593.html, 收稿日期 2012-03-14 修回日期2012-03-26* 图1自噬的基本过程[2] Jun;20(3) 236
线粒体脑肌病的诊治进展 MELAS综合征的临床研究进展 时间: 2009-02-13 14:54:00 作者:海总医院神经内科 1984年Pavlakis首先报告了线粒脑肌病伴高乳酸血症和卒中样发作(mitochondrial myopathy, encephalopathy, lactic acidosis and stroke-like episodes, MELAS)患者。随着人们对线粒体细胞病认识水平的提高,渐发现MELAS 综合征具有遗传基因和临床表型的广泛异质性。由于该病致残和致死率较高,故早期诊断和治疗十分重要。现结合临床实践从发病机制、临床表现、影像学特点、病理改变及遗传基因等方面对MELAS综合征进行综述。 一、病因和发病机制 目前有以下几种学说:①线粒体代谢异常学说:又称线粒体细胞病学说。线粒体是细胞内能量产生的重要场所,其功能异常可导致ATP产生减少,从而引起细胞凋亡。这样能量代谢旺盛的器官如脑、骨骼肌、心肌以及胰腺等器官最易累及。枕叶皮质与视觉有密切联系,其细胞和突触密度较高,因此,线粒体氧化磷酸化异常在枕叶最常受累。目前观点认为能量代谢异常可导致细胞稳态失调及慢性乳酸酸中毒,当能量需求增高时,可诱发脑卒中样发作[1]。研究发现MELAS
患者胰岛细胞线粒体功能异常可以导致胰岛素抵抗和胰岛 素分泌减少,引发糖尿病。因此,有的学者认为MELAS综合征为内分泌代谢障碍性疾病[2]。②线粒体血管病学说:该学说最早由Ohama 提出。病理形态学观察发现颅内小血管平滑肌细胞、内皮细胞以及肌间小血管壁琥珀酸脱氢酶(SDH)反应阳性;分子生物学研究发现MELAS患者颅内外血管壁的基因突变比率明显增高[3]。证实了异常线粒体在这些小血管壁平滑肌细胞和内皮细胞的沉积,最后导致脑组织缺血缺氧而致病。但该机制不能解释MELAS卒中样病灶不符合血管分布的特点。③非缺血性神经血管细胞学说:2002年Iizuka 和Sakai[4]发现MELAS患者卒中样发作的病理生理改变主要表现为毛细血管通透性增加、过度灌注、神经元易损伤以及神经元过度兴奋等,其中神经元过度兴奋发挥核心作用。2005年[5]提出了非缺血神经血管细胞学说,该学说认为病变区域的脑组织毛细血管内皮细胞、神经元及胶质细胞的线粒体功能不良导致病变区域的神经元兴奋性增高,其癫痫样活动使邻近区域神经元去极化,并向周围大脑皮层传播,最终导致能量代谢失衡,诱发脑卒中发作。2007年Iizuka 和Sakai[6]进一步证明MELAS患者卒中样发作是神经元兴奋性过高导致的非缺血神经血管事件,解释了MELAS卒中样病灶不符合血管分布的特点。但是,由于组织和器官能量需求不
第一节 线粒体DNA的结构和功能特征 一、mtDNA的结构特征 mtDNA是惟一存在于人类细胞质中的DNA分子,独立于细胞核染色体外的基因组,具有自我复制、转录和编码功能。人mtDNA由16 569bp组成,双链闭合环状,其中外环DNA单链由于含G较多,C较少,使整个外环DNA分子量较大,称为重链(heavy chain)或H链;而内环DNA单链则C含量高,G含量低,故分子量小,称为轻链(light chain)或L链。mtDNA的两条链都有编码功能,除与复制及转录有关的一小段D环区(displacement loop)无编码基因外,基因间无内含子序列;部分基因有重叠现象,即前一个基因的最后一段碱基与下一个基因的第一段碱基相重叠(图6-1)。因此,mtDNA的任何突变都会累及到基因组中的一个重要功能区域。mtDNA含有37个基因,其中两个rRNA基因 (16SrRNA,12SrRNA),22个tRNA基因,13个蛋白质基因(包括1个细胞色素b基因,2个ATP酶亚单位的基因。 图6-1 人线粒体基因图谱 Figure 6-1 Map of the human mitochondrial genome Box 6.1 The limited autonomy of the mitochondrial genome Encoded by Encoded by Mitochondrial nuclear
genome genome Components of oxidative phosphorylation system Ⅰ NADH dehydrogenase Ⅱ Succinate CoQ reductase Ⅲ Cytochrome b-c1 complex Ⅳ Cytochrome c oxidase complex Ⅴ ATP synthase complex Components of protein synthesis apparatus tRNA components rRNA components Ribosomal proteins Other mitochondrial proteins 13 subunits 7 subunits 0 subunits 1 subunits 3 subunits 2 subunits 24 22 tRNAs 2 rRNAs None None >80 subunits >41 subunits 4subunits 10 subunits 10 subunits 14 subunits ~80 None None ~80 All, e.g. mitochondrial enzymes and proteins 和7个呼吸链脱氢酶亚单位的基因)。位于D环区的HSP(heavy strand promoter)和LSP(light strand promoter)是线粒体基因组转录的两个主要启动子(图6-1)。 mtDNA是裸露的,不与组蛋白结合,存在于线粒体基质内或黏附于线粒体内膜。在一个线粒体内往往有一至数个mtDNA(图6-2)。mtDNA的自我复制也是以半保留复制方式进行。复制先从重链开始,形成一个约680个碱基的7sDNA,称D环。在对鼠细胞研究中发现,大多数的mtDNA均为D环的结构,只有一小部分mtDNA从D环开始合成完整的新生链。轻链的复制要晚于重链,等重链合成过OL之后才开始合成。研究发现mtDNA 的复制可以越过静止期或间期,甚至可以分布在细胞整个周期。mtDNA 的自我转录很似原核生物,即产生一个多顺反子,其中包括多个mRNA和散布于其中的tRNA,剪切位置往往发生在tRNA处,从而使不同的mRNA和tRNA被分离和释放。
2009年 第54卷 第9期: 1250 ~ 1261 https://www.doczj.com/doc/a311105593.html, https://www.doczj.com/doc/a311105593.html, 《中国科学》杂志社 SCIENCE IN CHINA PRESS 论 文 美姑脊蛇Achalinus meiguensis 线粒体基因组全序列及系统发育地位研究 王广力①, 何舜平②, 黄松③, 何苗①, 赵尔宓①④* ① 四川大学生命科学学院, 成都 610064; ② 中国科学院水生生物研究所, 武汉 430072; ③ 黄山学院生命与环境科学学院, 黄山 245041; ④ 中国科学院成都生物研究所, 成都 610041 * 联系人, E-mail: zem006@https://www.doczj.com/doc/a311105593.html, 2008-10-21收稿, 2009-03-13接受 摘要 报道了美姑脊蛇Achalinus meiguensis 线粒体基因组全序列. 美姑脊蛇线粒体全序列长17239 bp, 由22个tRNA, 2个rRNA 和13个蛋白质基因及2个非编码的控制区或D-loop 组成, 存在着基因重排现象. 对已报道蛇类线粒体基因组全序列进行比对分析后, 发现一些蛇类线粒体基因组进化规律: 双控制区现象在爬行动物进化历史中独立地发生, 有不同的演化历史; tRNA 假基因是在真蛇下目(Caenophidia)中进化形成的; T ΨC 臂的相对较短(一般少于 5 bp)和缺失“DHU”臂造成蛇类tRNA 较短. 通过MP 和BI 分析确定美姑脊蛇系统发育位置应该位于瘰鳞蛇Acrochordus granulatus 和其他真蛇下目蛇类之间, 而不应该属于游蛇科Colubridae 或者眼镜蛇科Elapidae. 由于美姑脊蛇与真蛇下目中蝰科Viperidae 、游蛇科以及眼镜蛇科之间的单系性都被统计检验拒绝(P < 0.01), 由此认为美姑脊蛇所在的闪皮蛇亚科(Subfamily, Xenodermatinae)应该提升到科或者更高的分类阶元. 依据系统发育统计检验结果, 我们选择Bayesian 树来进行分歧时间的估算. 原蛇下目与真蛇下目的分化时间为109.50 Mya; 而瘰鳞蛇与其他真蛇下目种类的分化时间为106.18 Mya; 美姑脊蛇的分化时间在103 Mya; 蝰科与眼镜蛇科和游蛇科构成的单系群的分化时间发生在96.06 Mya. 关键词 美姑脊蛇 线粒体全序列系统发育 分化时间 在不同动物类群中, 线粒体基因组在基因结构和基因排列方式等方面均显示了极大的多样性, 这种多样性可能反映了真核细胞不同的进化路线[1]. 就目前的研究而言, 线粒体基因组是一个能够从基因组水平上来分析动物系统发生的分子标记, 可以从线粒体基因组序列信息、基因组成及基因排列方式等进行多方位的分子进化研究. 因而线粒体基因组全序列成为研究动物分子系统发生最有力的证据之一[1] . 蛇的线粒体基因组大体构造与其他脊椎动物相似, 2个rRNA, 22个左右的tRNA 和13个左右的蛋白编码基因, 至少一个非编码区(control region), 但是蛇类线粒体全序列在其组成、结构与功能等方面又有着一 些与众不同的特点, 如紧凑的基因结构、双控制区、 缩短的tRNA 以及较快的进化速度等[2~5]. 脊蛇属Genus Achalinus Peters, 1869在分类上隶属于游蛇科Family Colubridae, 闪皮蛇亚科Subfamily Xenodermatinae [6,7]. 该属的模式种是黑脊蛇(Acha- linus spinalis ). 脊蛇属蛇类已知有9种, 除1种外, 在中国均有分布, 其中5种是中国特有种. 美姑脊蛇(Achalinus meiguensis Hu and Zhao, 1966)是中国的特产蛇类, 区别于其他脊蛇的主要形态特征是具鼻间鳞和一枚极小的眶后鳞[8]. 目前仅见于四川的美姑、安县、洪雅、峨眉、宝兴、汶川、卧龙、屏山等以及云南省水富县, 已报道的分布海拔在1500~3000 m 之
线粒体自噬 线粒体自噬研究概论 关于线粒体自噬 线粒体自噬(mitophagy)是指细胞通过自噬的机制选择性地清除线粒体的过程。选择性清除受损伤或功能不完整的线粒体对于整个线粒体网络的功能完整性和细胞生存来说十分关键。 线粒体自噬主要的作用有几个方面: 1.选择性清除功能受损的线粒体 2.选择性调节细胞内线粒体数量 3.通过线粒体影响诸多生理和病理学过程 Fig:The pathways of mitophagy for quality control and clearance of mitochondria Cell Death and Differentiation(2013)20,31–42
线粒体自噬的信号通路 1)Pink/Parkin pathway 2)Bnip3/Nix pathway 3)FUNDC1pathway Fig.Mitophagy pathway:Pink1/Parkin OR Bnip3/Nix Pink1/Parkin pathway:E3泛素连接酶Parkin和蛋白激酶Pink1一起介导了线粒体膜电位下降,引起的线粒体自噬的发生,当线粒体损伤后,线粒体膜电位下降,引起Pink1蛋白在损伤线粒体上的积累,能够吸引Parkin到损伤的线粒体上。Parkin使得线粒体外膜上的很多蛋白发生泛素化,从而能够募集其他一些相关蛋白,介导线粒体自噬的发生。
线粒体自噬 汉恒线粒体自噬研究工具与研究方法 汉恒生物有多种线粒体自噬病毒研究工具可以提供,便于直接感染目的细胞后直观地观察线粒体自噬的变化 一、汉恒线粒体自噬表型研究工具 1)Ad-GFP-LC3腺病毒病毒系统,可高效感染目的细胞,表达GFP-LC3,感染感染后细胞可在荧光显微镜下实时观察自噬的整体水平(由于GFP荧光偏弱,暂停Ad-GFP-LC3销售, 慢病毒单标LV-GFP-LC3荧光正常,正常销售); 2)Ad-HBmTur-Mito腺病毒系统(红光标记),为汉恒生物自主研发的线粒体特异性定位荧光探针(pHBmTur-Mito)可准确定位标记线粒体,结合汉恒独家推出的双荧光LC3细胞自噬腺病毒的使用,即可准确实时地追踪线粒体自噬的动态过程; 使用方法:Ad-GFP-LC3+Ad-HBmTur-Mito共感染目的细胞,confocal检测双荧光共定位的情况,如果共定位,则存在线粒体自噬!(下图说明:红色标记为线粒体,绿色标记自噬小体,二者有共定位时代表自噬发生) 二、汉恒线粒体自噬通路研究工具 1)Ad-Parkin-EGFP 2)Ad-Bnip3-EGFP+Ad-Nix-EGFP 3)Ad-FUNDC1-EGFP
线粒体的结构与功能 生命科学与食品工程系,050601030, 易永洁 摘要:线粒体是细胞质中重要的细胞器之一,普遍存在于真核细胞中。它是生物氧化和能量转换的主要场所,以氧化磷酸化(OXPHOS)方式将食物内蕴藏的能量转变为可被机体直接利用的ATP高能磷酸键。细胞生命活动所需能量的80%来源于线粒体,因此线粒体在细胞的生长代谢和人类的遗传中都有重要的作用。 关键词:线粒体;;结构;功能;遗传病;mtDNA 自1890年Altaman首次发现线粒体以来,生物学家就一直以极大的热情给予关注,到目前为止,其结构和功能方面的研究已经越来越深入明了。 1线粒体的结构 1.1外膜(out membrane) 含40%的脂类和60%的蛋白质,具有孔蛋白(porin)构成的亲水通道,允许分子量为5KD以下的分子通过,1KD以下的分子可自由通过。标志酶为单胺氧化酶。 1.2内膜(inner membrane) 含100种以上的多肽,蛋白质和脂类的比例高于3:1。心磷脂含量高(达20%)、缺乏胆固醇,类似于细菌。通透性很低,仅允许不带电荷的小分子物质通过,大分子和离子通过内膜时需要特殊的转运系统。如:丙酮酸和焦磷酸是利用H+梯度协同运输。 线粒体氧化磷酸化的电子传递链位于内膜,因此从能量转换角度来说,内膜起主要的作用。内膜的标志酶为细胞色素C氧化酶。 内膜向线粒体基质褶入形成嵴(cristae),嵴能显著扩大内膜表面积(达5~10倍),嵴有两种类型:①板层状、②管状,但多呈板层状。 1.3膜间隙(intermembrane space) 是内外膜之间的腔隙,延伸至嵴的轴心部,腔隙宽约6-8nm。由于外膜具有大量亲水孔道与细胞质相通,因此膜间隙的pH值与细胞质的相似。标志酶为腺苷酸激酶。 1.4基质(matrix) 为内膜和嵴包围的空间。除糖酵解在细胞质中进行外,其他的生物氧化过程都在线粒体中进行。催化三羧酸循环,脂肪酸和丙酮酸氧化的酶类均位于基质中,其标志酶为苹果酸脱氢酶。
电流检测方法 1 传统的电流检测方法 1. 1 利用功率管的RDS进行检测( RDS SENSIN G) 当功率管(MOSFET) 打开时,它工作在可变电阻区,可等效为一个小电阻。MOSFET 工作在可变电阻区时等效电阻为: 式中:μ为沟道载流子迁移率; COX 为单位面积的栅电容;V TH 为MOSFET 的开启电压。 如图1 所示,已知MOSFET 的等效电阻,可以通过检测MOSFET 漏源之间的电压来检测开关电流。 这种技术理论上很完美,它没有引入任何额外的功率损耗,不会影响芯片的效率,因而很实用。但是这种技术存在检测精度太低的致命缺点: (1) MOSFET 的RDS本身就是非线性的。 (2) 无论是芯片内部还是外部的MOSFET ,其RDS受μ, COX ,V TH影响很大。 (3) MOSFET 的RDS随温度呈指数规律变化(27~100 ℃变化量为35 %) 。 可看出,这种检测技术受工艺、温度的影响很大,其误差在- 50 %~ + 100 %。但是因为该电流检测电路简单,且没有任何额外的功耗,故可以用在对电流检测精度不高的情况下,如DC2DC 稳压器的过流保护。 图1 利用功率管的RDS进行电流检测
1. 2 使用检测场效应晶体管(SENSEFET) 这种电流检测技术在实际的工程应用中较为普遍。它的设计思想是: 如图2 在功率MOSFET两端并联一个电流检测FET ,检测FET 的有效宽度W 明显比功率MOSFET 要小很多。功率MOSFET 的有效宽度W 应是检测FET 的100 倍以上(假设两者的有效长度相等,下同) ,以此来保证检测FET 所带来的额外功率损耗尽可能的小。节点S 和M 的电流应该相等,以此来避免由于FET 沟道长度效应所引起的电流镜像不准确。 图2 使用场效应晶体管进行电流检测 在节点S 和M 电位相等的情况下,流过检测FET的电流IS 为功率MOSFET 电流IM 的1/ N ( N 为功率FET 和检测FET 的宽度之比) , IS 的值即可反映IM 的大小。 1. 3 检测场效应晶体管和检测电阻相结合 如图3 所示,这种检测技术是上一种的改进形式,只不过它的检测器件不是FET 而是小电阻。在这种检测电路中检测小电阻的阻值相对来说比检测FET 的RDS要精确很多,其检测精度也相对来说要高些,而且无需专门电路来保证功率FET 和检测FET 漏端的电压相等,降低了设计难度,但是其代价就是检测小电阻所带来的额外功率损耗比第一种检测技术的1/ N 2还要小( N 为功率FET 和检测FET 的宽度之比) 。此技术的缺点在于,由于M1 ,M3 的V DS不相等(考虑VDS对IDS的影响), IM 与IS 之比并不严格等于N ,但这个偏差相对来说是很小的,在工程中N 应尽可能的大, RSENSE应尽可能的小。在高效的、低压输出、大负载应用环境中,就可以采用这种检测技术。
哺乳动物自噬研究方法 【摘要】 自噬涉及到许多的生理和病理过程,因此,我们越来越需要科学地准确认识,并且量化操控自噬的过程。但是,由于自噬涉及了许多动态复杂的过程,关于它的研究分析经常不正确。在本文中,我们探讨监控自噬以及调控自噬活性的各种方法,主要集中关注哺乳动物中的大自噬。 【简介】 过去的十年中,有大量的研究围绕一个基本的细胞生物学通路“自噬”(希腊语中意味自我吞噬)。一系列进化上保守的基因(最初在酵母中被鉴定)被发现是自噬过程中所必需的,这一发现使得科学家能够去发现大量的自噬的功能,如:维持自身平衡稳态、参与细胞发展和其他生理活动。此外,越来越多的证据证明自噬的失控可能与许多哺乳动物的疾病发生有关。因此,科学家们迫切需要能够准确的检测自噬和研究在不同生物学进程中的功能的方法,特别是在哺乳动物系统。 在哺乳动物自噬的研究历史中一直有两个主要的困扰。首先的挑战在于如何将“一个动态的进程”和“静态的测量”进行捕获,并且这个固有的局限性与基于这些测量作出的生物学推论相关。第二个挑战在于将“形式”与“功能”分离,并且避免在给定的生理条件下,基于自身检测到(或没有)导致的将生理功能归至自噬的这个常见陷阱。这两个挑战可能导致了在我们研究哺乳动物自噬功能的历史中的许多误解。例如,一些神经退行性和肌退化性疾病最初被认定的结果,至少一部分被认为是由于自噬的增加(基于显微镜下观察到通路中早期的中间产物的增加)。然而实际上,早期中间产物的蓄积在这些疾病中代表着后阶段自噬通路的阻滞。自噬的一个常见的形态特征是细胞的垂死状态,但这也被错误的认为是一种细胞死亡通路,然而,现在似乎已经很明确自噬的主要功能是帮助细胞抵抗各种“生死攸关”的应激条件并使细胞存活。 这些哺乳动物自噬研究中的历史挑战部分已经通过将阐述自噬分子机制的最新进展运用到新的自噬研究的方法被克服。因此,在过去的十年里,许多新的技术被发明,用于动态监控自噬和通过调控自噬来明确其在给定的细胞状态下的功能。本文的目的在于提供一个重要的关于目前可行的研究哺乳动物自噬的技术和这些技术在解释过程中的限制的概述。更多的各种技术的细节信息可以再其他综述中获得。 自噬的基础知识 自噬是一般用于描述细胞内物质包括细胞器到溶酶体中降解的过程。在自噬的三种类型中(大自噬、小自噬和分子伴侣介导的自噬),研究得最多的就是大自噬。分子伴侣介导的自噬是通过伴侣蛋白展开蛋白质,直接将细胞质蛋白转移,穿过溶酶体膜。小自噬涉及溶酶体膜表面的内陷提供一小部分细胞质进入溶酶体内腔。 大自噬(简称自噬)是本文主要关注的通路。这个通路从酵母到哺乳类都呈现保守,它是通过一些特定的细胞器(自噬体)介导的。在初始的诱导阶段,一个小的囊状的称作隔离膜或者延长的吞噬膜,随后封闭一部分细胞质,导致双层膜结构即自噬体的形成。然后自噬体的外膜融合至一个溶酶体(形成自噬溶酶体),导致所封闭的物质和自噬体的内膜共同降
兰州交通大学化学与生物工程学院 综合能力训练I 文献综述 题目:线粒体疾病的最新研究进展 作者:朱刚刚
学号:201207730 指导教师:谢放 完成日期:2014-7-16 线粒体疾病的最新研究进展 摘要:本文为了对线粒体疾病研究的最新进展进行论述,分别从线粒体功能障碍、线粒体疾病、以及相关线粒体疾病的治疗与干预策略三个方面进行了综述。重点从线粒体的功能障碍进行了介绍。 关键词:线粒体、线粒体tDNA、线粒体疾病。 引言:线粒体疾病主要是指由于线粒体DNA突变所导致的一类疾病。 有许多人类疾病的发生与线粒体功能缺陷相关,如线粒体肌病和脑肌病、线粒体眼病,老年性痴呆、帕金森病、O型糖尿病、心肌病及衰老等,有人统称为线粒体疾病。线粒体疾病的发生被认为与氧化磷酸化过程相关基因的突变有关。 一、线粒体功能障碍 1线粒体结构、基因组特征及主要功能 1.1 线粒体结构及基因组特征电镜下的线粒体是由两层单位膜套叠而 成的封闭囊状结构,从外向内依次分为外膜、膜间隙、内膜、基质。不同于经典的“隔舱板”理论,最新提出的三维重构模型认为:(1)外膜与内质网或细胞骨架连接形成网络;⑵内外膜间随机分布横跨两端,宽20nm的接触点;(3)内膜通过界面与嵴膜接口部分相连,并不直接向内延伸形成嵴膜;(4)嵴膜非“隔舱板”式而是管状或扁平状,相互间可连接或融合,呈现不同的形式。执行线粒体功能的生物大分子分布在不同的空间:外膜上有Bcl-2家族蛋白、膜孔蛋白以及离子 通道蛋白;内膜中有电子传递链(呼吸链)复合物l~IV和复合物V(ATP合成酶); 膜间隙和嵴膜腔分布着细胞色素C、凋亡诱导因子(apoptosis in-dueing factor,AIF)和Procaspase 2、3、9及其他酶蛋白;电压依赖性阴离子通道(VDAC)、ADP/ATP 转换蛋白(ANT)和线粒体膜转运孔
MDC:取12 mg粉末溶于720 nl DMSO使其浓度为50 mmol/L,分装后-20冰箱保存。 临用前用MEM稀释到终浓度50 umol/L; Rapamycin:用MEM培养基配成终浓度为1 umol/L,现用现配; 400ng/ml喹乙醇:称取4 mg喹乙醇,DMSO预溶(体积<0.1%)后加入10 ml MEM培养液至完全溶解,现用现配,避光保存; 3-MA:首先用PBS溶解粉末,临用前加热至完全溶解后再加入MEM培养基至终浓度10mmol/L; PI3K抑制剂(3-MA,Wortmannin)可干扰或阻断自噬体的形成 用RAPAMYCIN诱导自噬我也查过一部分文献,有用无血清的,也有用,一般培养基的,浓度从25nM到100nM都有,用的是50nM的雷帕霉素,加入一般的培养基中,目的是排除无血清所诱导出来的自噬。 文献说饥饿初期激活的是大分子自噬,在4-6小时活力达到最大,24h后以CMA途径为主Earle's balanced salts solution (EBSS) for 48 h sigma的EBSS,货号E2888,有碳酸氢钠,有酚红的,酚红到不是很必须,只是一个PH指示作用,好看些 无血清诱导自噬:EBSS 诱导6个小时就可以了。 EBSS一定可以诱导出来,只是需要说明的是时间点的设置,因为从饥饿诱导开始半个小时就可能开始自噬了,一直到24小时都持续,所以应该设置不同的时间点观察这个作用。另外一个很大的问题是,饥饿诱导的一个很大的弊端是细胞死亡,这也是我面临的问题,就是在细胞收养的时候蛋白浓度太小了。24小时就很少了,更不要说48小时和72小时了 Hank's诱导,也就是通常所说的饥饿诱导,细胞培养到对数生长期后以Hank's替代常规完全培养基,3h后就可诱导出自噬。我用Hank's诱导了3h后电镜观察有30%细胞都有自噬这种现象,但不如国外报道的高。 sigma的氯喹的货号C6628。用氯喹做自噬抑制剂,293T细胞50uM就可以。1. 可以用双蒸水配制2. 配制后4度保存 不同的自噬抑制剂机制不同。抑制的步骤也不同。有的不能抑制lc3的剪切,但能抑制后续的步骤,Chloroquine抑制自噬体与溶酶体的融合过程,autophgy不能完成,所以lc3才会累积。因此加了抑制剂lc3之后会比不加的要高。氯喹能提高溶酶体中的pH值,使溶酶体中的酸性水解酶丧失活性,从而导致“自噬溶酶体”不能降解,因此,位于自噬体和自噬溶酶体膜上的LC3不能按时降解,表现为LC3荧光长时间的保留或WB中LC3条带变粗。 Z-VAD-FMK(caspase-3 抑制剂)抑制EV71感染所引起的细胞凋亡,观察细胞的自噬情况。研究发现,抑制细胞凋亡能增加LC3-I转化为LC3-II以及p62的降解。 1. 雷帕霉素:作为以mTOR 为靶点最经典的诱导剂已经被广为应用,推荐工作浓度为1μmol-10μmol; 2. 氯喹:氯喹(Chloroquine)作为溶酶体的抑制剂,可以抑制自噬体与溶酶体的融合从而可以用来作为自噬以及自噬流的抑制剂用于实验研究,推荐使用浓度:10umol-50umol。 正常培养的细胞自噬活性很低,不适于观察,因此,必须对自噬进行人工干预和调节,经报道的
ISS N 100727626C N 1123870ΠQ 中国生物化学与分子生物学报 Chinese Journal of Biochemistry and M olecular Biology 2008年12月 24(12):1170~1176 ?研究简报? 中国狼(Canis lupus chanco )线粒体全基因组序列分析 孟 超1) , 张洪海 1)3 , 陈玉才 2) (1)曲阜师范大学生命科学学院,曲阜 273165; 2) 济南跑马岭野生动物园,济南 250000) Sequencing and Analysis of Mitochondrial G enome of Chinese G rey Wolf (Canis lupus chanco ) ME NG Chao 1) ,ZH ANG H ong 2Hai 1)3 ,CHE N Y u 2Cai 2) (1)College o f Life Science ,Qufu Normal Univer sity ,Qufu 273165,Shandong ,China ; 2) Jinan Paomaling Wildlife Park ,Jinan 250000,China ) 摘要 参照近缘物种线粒体全基因组序列,设计14对特异引物,采用PCR 产物直接测序法测得中国狼线粒体基因组全序列,并分析其基因组特点和各基因的定位.用pDRAW32软件,预测12种限制性酶的酶切图谱.结果表明,中国狼线粒体基因组全长16774bp ,包括13个蛋白质编码基因、2个rRNA 基因、22个tRNA 基因和1个非编码控制区.除ND 2、ND 3和ND 5基因以AT A 作为起始密码子外,其它基因的起始密码子均为ATG.除COX Ⅲ、ND 4、ND 3基因的终止密码子分别为不完全的T ,T ,T A ;ND 2,COX Ⅱ,Cytb 基因的终止密码子分别是T AG 、T AG 、AG A 外;其余基因均以T AA 作为终止密码子,而欧亚狼和狗COX Ⅱ基因则以T AA 终止.基于近缘哺乳动物15种近缘物种的线粒体基因组的12S rRNA 和16S rRNA 基因全序列,用邻近法、最大简约法和最大似然法构建系统进化树,系统进化关系与传统的系统分类基本一致.并在已有文献的基础上,探讨了中国狼的进化地位.关键词 中国狼;线粒体基因组;序列分析;系统发育中图分类号 Q75 收稿日期:2008205229;接受日期:2008209203 国家自然科学基金资助项目(N o 130370218),山东省自然科学基金 (N o 12002B13),山东省科技攻关项目(N o.2007GG 2009011),山东省教 育厅科技计划重点项目(J07YG 06),教育部新世纪优秀人才支持计划 (NCET 2070507) 3 联系人 T el :0537********;E 2mail :zhanghonghai67@https://www.doczj.com/doc/a311105593.html, Received :M ay 29,2008;Accepted :September 3,2008 Supported by National Natural Science F oundation of China (N o 130370218),Natural Science F oundation of Shandong Province (N o 12002B13),Sci 2tech Development Program of Shandong Province (N o.2007GG 200911), Education Department G rants of Shandong Province (J07YG 06),Program for New Century Excellent T alents in University (N o 1NCET 2070507) 3 C orresponding author T el :0537********; E 2mail :zhanghonghai67@1261com 动物线粒体基因组DNA 是核外遗传物质,结构简单,呈母系遗传,在世代遗传中不会发生重组,广泛用于分析动物类群间的系统关系 [1~5] .线粒体基 因组是现在惟一能够从基因组水平来分析动物系统发生的分子标记,因而成为动物分子系统发生研究的最有力的证据 [6,7] .截至2008年5月,在G enBank 数据库中,食肉目犬科动物中有狗(Canis lupus f amiliaris )、欧亚狼(Canis lupus lupus )、郊狼(Canis latrans )和赤狐(Vulpes vulpes )的线粒体基因组全序列 被测出. 灰狼属于哺乳动物食肉目犬科,划分为22个亚种[8] ,中国只有1个亚种———中国狼或西藏狼(Canis lupus chanco )[9],已被列为国家二级保护动物.由于灰狼和犬形态的相似,国内外学者曾利用考古学、形态学等研究方法对二者的进化关系进行了研究 [10~13],提出狗起源于灰狼的推测.近年来,研究 人员广泛采用线粒体基因组DNA 研究狼和狗的系统进化,不仅支持了狗起源于灰狼的推测 [14,15] ,还推 断出家犬起源的地点在东亚[16] . 为了进一步研究以中国狼为代表的东亚地区的狼和狗的进化关系以及中国狼的系统发育,本文对中国狼(Canis lupus chanco )线粒体全基因组序列进行了测定与分析,基于12S rRNA 和16S rRNA 基因全序列及G enBank 上近缘物种的相应基因序列进行