
论文题目:ARMA模型的阻尼最小二乘法班级: 姓名: 学号: 指导教师:
摘要 ARMA模型是将实际问题利用时间序列建立起的模型,只要把ARMA模型的参数估计出来,实际问题就能解决了。本文只对讨论了ARMA模型参数的优化理论估计方法的一种:阻尼最小二乘法。非线性时间序列ARMA模型参数的优化估计法一阻尼最小二乘法,它结合了Newton法和最速下降法的优点,既保证了迭代计算的收敛性,又加快了收敛的速度。当初值的精度较差时,更宜采用阻尼最小二乘法。本文给出实例的MATLAB程序,并利用t统计量检验出阻尼最小二乘法要比最小二乘法的参数估计值更为显著,拟合模型更优。 关键词:非线性;阻尼最小二乘法;ARMA;MATLAB Abstract ARMA model is to establish a real problem using time series models, As long as the ARMA model parameters estimated from the actual problem can be solved. Nonlinear time series ARMA model parameter optimization estimation method—Damped least squares method, It combines the advantage of Newton method and the steepest descent method, It not only ensures the convergence of iterative calculations, but also accelerate the speed of convergence. When the accuracy of the original value is poor, it better to using qualified damped least squares method. This paper gives examples of the MATLAB program,And use the t-statistic tests the damped least squares method more significant than the method of least squares parameter estimates, and better fitting model. Keywords: Nonlinear; Damped least squares method; ARMA; MATLAB
逻辑斯蒂方程及经济学应用 梁美娟,生物0801,20080205035 摘 要:逻辑斯蒂方程是一种非线性微分方程,其数学模型S 型曲线模型被广泛应用于描述事物的增长,本文系统的阐述了该方程的历史和演变,分析其生态学意义,并说明了该模型在经济学上的应用。 关键词:逻辑斯蒂方程;Lotka-V olterra 模型;前景理论;S 型曲线 一 前言 逻辑斯蒂方程广泛应用于描述客观事物的S 型变化现象。逻辑斯蒂数学模型是一条单调递增的,单参数k 为渐近线的S 型曲线。基本数量特征是当t 很小的时,呈指数增长,而当t 很大时,增长速度下降,且接近一个值(k )趋于平稳。利用它我们可以表征种群的数量动态,描述客观事物增长过程,还可以对满足该方程的现象进行预测,有助于相关政策的制定。另外,logistic 方程还可以作为其它模型如Lotka-V olterra 竞争模型的理论基础。 二.逻辑斯蒂方程的历史和演变 最早在1798年,英国统计学家Malthus(1766-1843)的《人口原理》中提出闻名于世的Malthus 人口模型。假设:在人口自然增长过程中,相对净增长率(出生率减死亡率)为常数,即单位时间人口的增长是与人口正比例,比例系数r 。 ?????==0 )(0N N rN dt dN t (1) 该模型准确反映了1700-1964年的人口增长,表明人口以指数规律随时间无限增长。但不是适应与以后的增长。因地球上各资源只可供一定数量的人生活,人口增加,环境的限制越来越明显,r 减少。1838年,比利时数学家P.F.Verhulst 引入N m ,表示自然条件所能容纳的最大人口数,Verhulst 假设的有限环境的物种相对增长率为 ?????=-=0 )(0)1(N N N K N r dt dN t (2) 由曲线得出以下结论:不管初值为多少,人口总量最终接近于极限值K ,极限值的一半(即r/2K )前,是加速生长的时候,过了这一点以后,增长速度减少,并且迟早会达到零。 三、逻辑斯蒂方程的生态学应用 1、在种群生态学中,种群的增长是一个复杂的问题,,由于种群手到诸多因素的影响,如环境条件、营养条件、出生率、死亡率、个体基数及时代特征等。
自然界的多种群模型分析 摘要:在我们生活的大自然中,有着太多太多的秩序和规则。种群之间的你争我斗,弱肉强食也是非常激烈。种群,顾名思义就是指同一种生物的一个集合。不同种群之间的关系大致分为四种:捕食与被捕食关系,互利共生关系,相互竞争关系和寄生与寄主关系。我们这次的建模就是围绕着种群之间的关系来展开的,下面我将从这几个方面来进行分类讨论,由于寄生与寄主的关系不是很常见,关系也比较简单,在此便不再赘述。 捕食与被捕食关系:这种关系很简单,大家也能很容易地理解,通俗地解释,就是指一种生物以另一种生物为食,举个例子大家也许会更容易地理解。比如说狼和羊的关系,狼是捕食者,羊是被捕食者,狼以羊为食,是羊的天敌。 互利共生关系:指两种生物共同生活在一个区域有助于提高另一种生物的种群密度,假如其中一种生物的数量减少,也会影响另一种生物的数量,使其数量减少。比如草地和森林优势植物的根多与真菌共生形成菌根,多数有花植物依赖昆虫传粉,大部分动物的消化道也包含着微生物群落,最典型的就是大豆与根瘤菌。大豆给根瘤菌提供养分,根瘤菌给大豆提供氮元素。 相互竞争关系:有种内和种间两种竞争方式。这里是指两种共居一起,为争夺有限的营养、空间和其他共同需要而发生斗争的种间关系。竞争的结果,或对竞争双方都有抑制作用,大多数的情况是对一方有利,另一方被淘汰,一方替代另一方。举个例子,牛和羊生活在共同的一片草地上,因为这两种生物都以草为食,它们之间不存在其他关系,所以它们之间是竞争关系。 以上就是三种种群之间的关系,下面我们就从这三个方面对物种种群密度的变化进行分析。在以下的讨论中我们将建立微分方程的数学模型,对生物多种群之间各种关系进行 关键词:生物种群,数量,关系,互相作用,竞争 问题重述: 生物学的研究对维持地球生态平衡有着不可替代的作用,是可持续发展的重要组成部分!地球上的物种一直只在减少,现在也有很多物种濒临灭绝,因此对
讨论问题:在按年龄分组的种群增长模型中,设一群动物的最高年龄为15岁,每5岁一组,分成3个年龄组,各组的繁殖率为b1=0,b2=4,b3=3,存活率为s1=1/2,s2=1/4,开始时3组各有1000只。求15年后各组分别有多少只,以及时间充分长以后种群的增长率(即固有增长率)和按年龄的分布。 成员: 按年龄分组的种群增长 不同年龄组的繁殖率和死亡率不同 以雌性个体数量为对象 建立差分方程模型,讨论稳定状况下种群的增长规律 模型建立 种群按年龄大小等分为n 个年龄组,记i=1,2,… , n 时间离散为时段,长度与年龄组区间相等,k=1,2,… 第i 年龄组1雌性个体在1 时段内的繁殖率为bi 第i 年龄组在1时段内的死亡率为di, 存活率为si=1- di xi(k)~时段k 第i 年龄组的种群数量 ) ( ) 1 ( 11 k x b k x i n i i ( 设至少 1 个 b i >0)
T n k x k x k x k x )] (),(),([)(21 ~按年龄组的分布向量 X(k+1)=LX(k),k=0,1,2,… 当矩阵L 和按年龄组的初始分布向量x (0)已知时,可以预测任意时段k 种群按年龄组的分布为: 稳定状态分析的数学知识 1 , , 2 , 1 ), ( ) 1 ( 1 n i k x s k x i i i 0 0 0 12 1 12 1 n n n s s s b b b b L ) ( ) ( x L k x k
矩阵存在正单特征根1, >0, 则1 ( lim cx k x k k T n n s s s s s 11 1 2 2 1 2 1 1 1 ,, ,, 1 1 ( lim cx k x k k 0, )( lim 1 1 x Pdiag k x k k
逻辑斯蒂模型(Logistic growth model ) 1.原始逻辑斯蒂模型: 设0t 时刻的人口总数为)(0t N ,t 时刻人口总数为)(t N ,则: ?????==0 0)(N t N rN dt dN 但是这个模型有很大的局限性:只考虑出生率和死亡率,而没有考虑环境因素,实际上人类生存的环境中资源并不是无限的,因而人口的增长也不可能是无限的。此人口模型只符合人口的过去而不能用来预测未来人口总数。 2.改进逻辑斯蒂模型: 考虑自然资源和环境对人口的影响,实际上人类所生存的环境中资源并不是无限的,因而人口的增长也不可能是无限的,因此,将人口增长率为常数这一假设修改为:?????=-=0 02)(N t N KN rN dt dN 其中K r ,称为生命系数 分析如下: rt t t e r K N r K t N -∞→∞→-+=)1(1lim )(lim 0 0)1(1lim 0?-+=∞→r K N r K t = K r N KN r KN r KN r dt dN KN r dt dN KN dt dN r dt N d ))(2)(2()2(222---=-=-= 说明: (1)当∞→t 时,K r t N → )(,结论是不管其初值,人口总数最终将趋向于极限值K r /; (2)当K r N 00时,0)(2 N K r KN KN rN dt dN -=-=,说明)(t N 是时间的单调递增函数; (3)当K r N 2 时,022 dt N d ,曲线上凹,当K r N 2 时,022 dt N d ,曲线下凹。
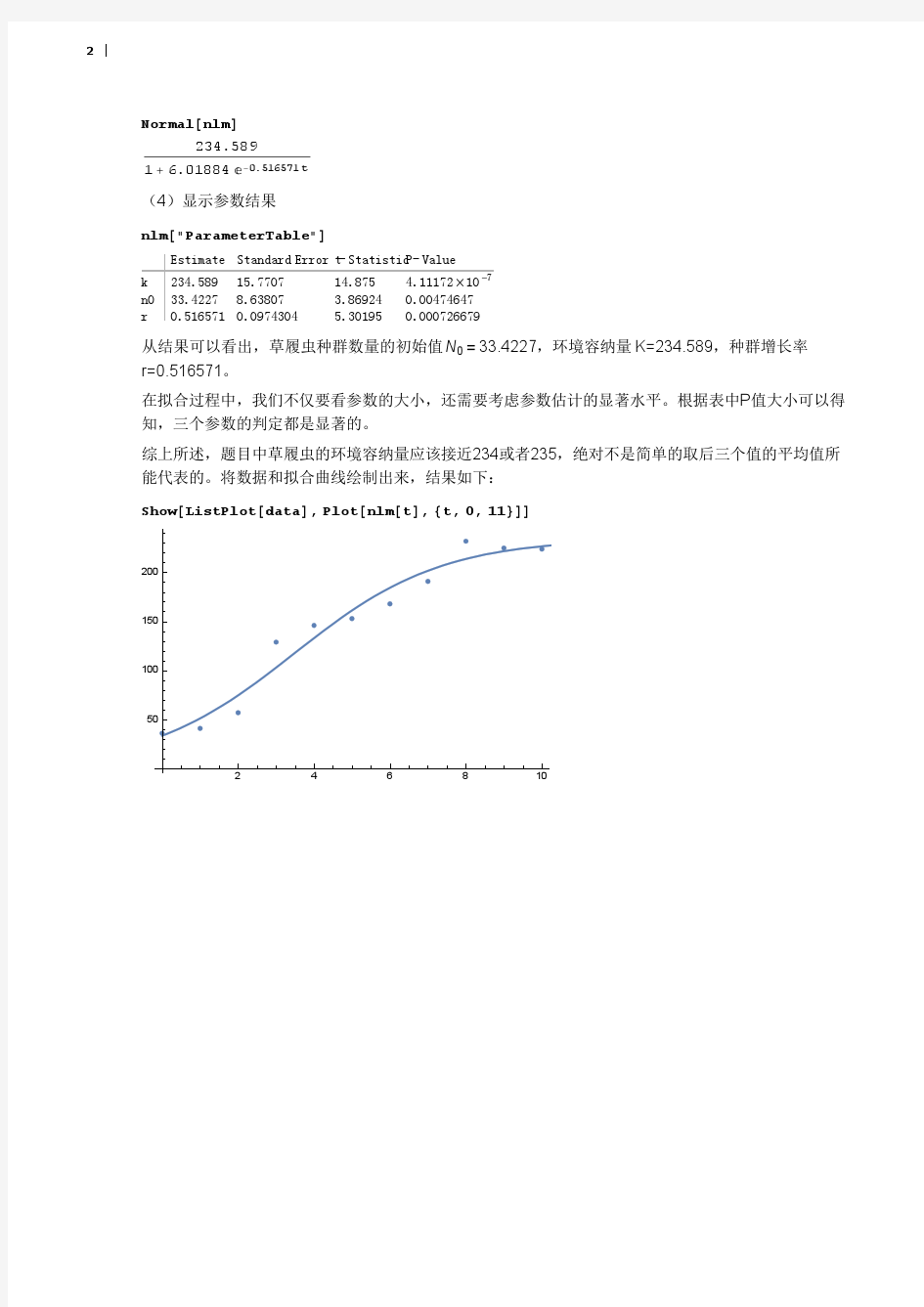
表九用spss软件得到各观察值所对应的拟核值,残差值和标准残差 拟合值97077.7 101458.9 105412.6 108940.84 112057.91 114787.4 117159.2 残差-818.74 -2753.91 438.35 3763.15 2275.08 1035.51 11.73 标准残 -0.7505 -2.0548 0.3051 2.5699 1.5537 0.7098 0.0080 差 拟合值119206.2120962.7122462.4123737.3124817.2125729.2126497.3残差-689.28-1112.76-1341.41-1348.34-1191.28-968.25-711.37标准残 -0.4707-0.7540-0.9009-0.8985-0.7899-0.6410-0.4720差 拟合值127142.9127684.4128138.0128517.4128834.5129099.2 残差-399.93-57.47314.93709.501153.451656.76 标准残 -0.2670-0.03870.21470.49060.81010.941 差 从新数据得到F=372.3471 p值=0.001 从新数据得到相关系数R=0.9888,相关性比较强,说明这种拟合是比较贴切的,本文建立逻辑斯蒂模型:0.8840.185 =+ y e-- 130517.5/(1)x
§4 从倍周期分定走向混沌 4-1 逻辑斯谛(Logistic )映射 我们将以一个非常简单的数学模型来加以说明从倍周期分定走向混沌现象。该模型称为有限环境中无世代交替昆虫生息繁衍模型。若昆虫不加以条件控制,每年增加λ倍,我们将一年作为一代,把第几代的虫日记为,则有: i N o i i i N N N 11++==λλ (4-1) i N ,1>λ增长很快,发生“虫口爆炸”,但虫口太多则会由于争夺有限食物和生存空间, 以及由于接触传染导致疾病曼延,使虫口数目减少,它正比于,假定虫口环境允许的最大虫口为,并令2 i N o N o i i N N x = ,则该模型由一个迭代方程表示: 21i i i N N N λλ?=+ 即为: )1(1i i i x x x ?=+λ (4-2) 其中:]4,0[], 1,0[∈∈λi x 。 (4-2)式就是有名的逻辑斯谛映射。 4-2 倍周期分歧走向混沌 借助于对这一非线性迭代方程进行迭代计算,我们可以清楚地看到非线性系统通过倍周期分岔进入混沌状态的途径。 (一)迭代过程 迭代过程可以用图解来表示。图4-1中的水平轴表示,竖直轴表示,抛物线表示(4-2)式右端的迭代函数。45o线表示n x 1+n x n n x x =+1的关系。由水平轴上的初始点作竖直线,找到与抛物线的交点,A 的纵坐标就是。由点 )0,(0x R ),(10x x A 1x
),(10x x A 作水平直线,求它与45o线的交点,经B 点再作竖直线,求得与抛物 线的交点,这样就得到了。仿此做法可得到所迭代点。 ),(11x x B ),(21x x 2x 从任何初始值出发迭代时,一般有个暂态过程。但我们关心的不是暂态过程,而是这所趋向的终态集。终态集的情况与控制参数λ有很大关系。增加λ值就意味着增加系统的非线性的程度。改变λ值,不仅仅改变了终态的量,而且也改变了终态的质。它所影响的不仅仅是终态所包含的定态的个数和大小,而且也影响到终态究竟会不会达到稳定。 (二)终态性质 ①当31<<γ时,迭代结果的归宿是一个确定值,趋于一个不动点,即抛物线与45o线的交点,这相当于系统处于一个稳定态,如图4-2(a)所示。此值与λ有关,且与λ值有一一对应关系。当4.2=λ时,12/711==+i x x 。迭代的结果为一个不动点的情况,其周期为1,这表示从出发,迭代一次就回到。 i x i x ②当449.33<<γ时,迭代的终态在一个正方形上循环,亦即在两个值之间往复跳跃,与一个i x λ值对应将有两个值,即其归宿轮流取两个值,如图4-2(b)所示。当i x 2.3=λ时,此值为i i x x =?+2,7995.05130.0周期为2,表示从出发,迭代二次后回到。所以,从图3-12(a)到3-12(b)中间发生了一个倍周期分岔,一个稳定态分裂成为两 i x i x 图4-2 叠代过程 种状态,而系统便在两个交替变动的值间来回振荡。 ③当544.3449.3<<λ时,最终在四个值之间循环跳跃,如图4-2(c)所示。 +4,即终态集是个四周期解,表示从出发,迭代四次后回到。所以,从图4-2(b) 到3-12(c),中间又发生了一个倍周期分岔,两种状态分裂成四种状态,而系统便在四个交 i x i i x x i x i x =
第六次建模作业 组员:何睿洁 张鹏 刘顺 一.logistic 模型模拟 【摘要】物种种群数量的变化规律一直是我们所探究的问题,考虑到一些自然灾害和物种间的食物链或竞争关系,我们可以在一定条件下模拟某一种群的变化规律。对于人口的增长一直是一个热门话题,我们通过数据的统计和拟合可以总结出某地区的人口变化规律,并在其他地区进行模型检验,分析该动态机理模型是否在一定程度上成立。 【关键词】人口增长 数据统计 模型检验 动态机理模型 【问题重述】美国人口数据随时间的变化: 1790 1800 1810 1820 1830 1840 1850 3.9 5.3 7.2 9.6 12.9 17.1 23.2 1860 1870 1880 1890 1900 1910 1920 31.4 38.6 50.2 62.9 76.0 92.0 106.5 1930 1940 1950 1960 1970 1980 1990 2000 123 132 151 179 204 227 251 281 【模型建立】首先我们可用微积分的思想将连续的微分方程离散化,不妨设x(n)表示第n 次普查所得人口数,根据logistic 模型 dy/dt=r(1-y/K)y 可得: ())())(1()()1(n x K n x r n x n x -=-+ 进一步化简有 ))(1()()()1(K n x r n x n x n x -=-+
令 )()()1(n x n x n x u -+= ,)(n x v = 可得: K rv r u -= 【求解模型】现在我们可以用线性拟合,借助matlab 来进行运算得到r ,K 运行程序: X=[3.9 ; 5.3 ; 7.2 ; 9.6 ; 12.9; 17.1; 23.2; 31.4; 38.6 ; 50.2 ; 62.9 ; 76.0 ; 92.0 ; 106.5; 123 ; 132 ; 151; 179 ; 204 ; 227 ; 251 ; 281]; Y=[] for i=1:21 Y(i)=(X(i+1,:)-X(i,:))./(X(i,:)); Y=[Y ,Y(i)] End 运行结果运用cftool 工具线性模拟: Result Linear model Poly1: f(x) = p1*x + p2 Coefficients (with 95% confidence bounds): p1 = -0.0009825 (-0.001254, -0.0007108) p2 = 0.3178 (0.2832, 0.3525)
种群增长的Gompertz 模型 摘要 本文根据题目要求,在渔场鱼量的自然生长服从种族增长规律Gompertz 模型的情况下,建立捕捞情况下渔场产量模型。根据模型,对渔场鱼量的平衡点及其稳定性进行讨论,并且在稳定的前提下,使用图解法讨论如何控制捕捞使持续产量达到最大。最后,对模型的优缺点进行了讨论。 关键词:Gompertz 模型 稳定性模型 图解法 正文 1 问题复述 已知某渔场鱼量的自然生长服从种族增长规律Gompertz 模型:().ln N x t rx x =,其中r 是固有增长率,N 是环境容许的最大鱼量。并且单位时间捕捞量为h Ex =,其中比例常数E 表示单位时间捕捞率,又称捕捞强度。现要求: (1)建立在捕捞情况下渔场鱼量的数学模型,讨论渔场鱼量的平衡点及其稳定性; (2)在鱼量稳定的前提下,求最大持续产量m h 及获得最大产量的捕捞强度m E 和 渔场鱼量水平*0x 。 2 模型假设 (1)捕捞过程视为连续性过程; (2)忽略种群间的相互作用及环境突变对渔场鱼量变造成的影响。 3 符号说明 ()x t 表示时刻t 时渔场中的鱼量; ()0,1i x i =表示渔场鱼量平衡点; *0x 表示获得最大持续产量的渔场鱼量水平; r 表示种群的固有增长率; N 表示环境容许的最大鱼量; ()f x 表示单位时间渔场鱼量的增长量; ()h x 表示单位时间的捕捞量; m h 表示单位时间的最大持续产量; ()F x 表示在捕捞情况下渔场的鱼量; ()'F x 表示()F x 的导数;
E 表示单位时间捕捞率,即捕捞强度; m E 表示获得最大持续产量时的捕捞强度; 4 模型建立 (1)在无捕捞条件下,()x t 的增长服从Gompertz 规律,即 ()().ln N x t f x rx x == ① (2)单位时间的捕捞量(即产量)()h x 与渔场鱼量()x t 成正比,比例系数为E ,于是单位时间的捕捞量为 ()h x Ex = ② (3)由①式与②式可以得到捕捞情况下渔场鱼量满足的方程 ()().ln N x t F x rx Ex x ==- ③ 5 模型求解 渔场鱼量平衡点及其稳定性讨论 根据上面得到的在捕捞情况下渔场的鱼量()F x 所满足的方程③式,令 ()ln 0N F x rx Ex x =-= 得到两个平衡点 01,0E r N x x e == ④ 由于()'ln N F x r r E x =--,因此有()'00F x r =-<,故0x 点稳定(与E ,r 的大小无关);同时,可证1x 点不稳定。 渔场鱼量稳定前提下持续产量最大问题的讨论 根据①,②式作曲线()y f x =和直线()y h x Ex ==,如图1所示。由于稳定点0x 与E ,r 的大小无关,因此应用图解法,由图1可知,当y Ex =与()y f x =在顶点*P 相交时可获得最大持续产量,此时的稳定平衡点为 *01N N x e = ⑤ 且单位时间的最大持续产量为
第六章 ARMA 模型的参数估计—主要内容 §6.1 AR(p)模型的参数估计 问题: 已知p 的AR(p): 1 ,0p t j t j t j X a X t ε-==+≥∑,2~WN(0,)t εσ.(1.1) 由12{,,,}N x x x 去估计12(,,,)T p a a a =a 和2σ. 1. AR(p)模型的Yule-Walker 估计 自回归系数p a 由自协方差函数{}k γ惟一确定.
1111210 2212 0p p p p p p a a a γγγγγγγγγγγγ----?????? ?????? ??=??????????????? ??????? 白噪声的方差2σ由2 0T p p σγ=-γa 决定. 现获12{,, ,}N x x x , N p >, 则作 (1) ,1~t t N y x x t N =-=; (2) 1 1 ?,0,1, ,N k k j j k j y y k p N γ-+== =∑;
(3) 只要12,, ,N x x x 不全同, 则?p Γ正定, 得惟一 1???p p p -=a Γγ, 2100????????T T p p p p p σγγ-=-=-γa γΓγ. 实用中, Levinson 递推公式(无需求逆, 快): (1)2 001,1 10222 1,1,11,2 1,1,101,12,2,1,,1,1,1????????(1)???????...????????...????,1,k k k k k k k k k k k k k k k k k k k j k j k k k k j a a a a a a a a a a a a a j k k p σγγγσ σγγγγγγγγ-+-++++++-?=?=??=-??----?=?----? =-≤≤≤??
生态学实验指导 浙江大学生命科学学院 生物科学系 生态学课程组 2012年2月
目录 前言 (1) 实验一、二种群增长观测及Logistic增长模型 (2) 实验三、四代谢生态-群体密度调控指数确定 (4) 实验五、六环境诱导细胞钙信号对气孔开闭的调控 (6) 实验七、八群落物种多样性测定与计算 (8) 实验九、十城市植被与土壤碳特征的关系 (10) 实验十一、十二城市树木年龄与生产力关系 (15) 实验十三、十四植物吸收水中铵氮能力比较分析 (16) 实验十五、十六植物吸收水中硝氮能力比较分析 (18)
前言 “工欲善其事,必先利其器”——在科学研究的构成中,方法占有极其重要的意义,甚至可以说,一部科学发展的历史就是一部科学方法不断发展的历史。生态学是生命科学的一个分支学科,同其他学科一样,实验是其最重要的内容。掌握基本方法,掌握基本技能,对于理解生态学理论,并将之运用于实践,具有重要意义。 在科学方法论中,实验是狭义的。包括我们经常说到的观察和实验两个内容。科学观察是指在自然条件下,人们对自然现象进行搜集、描述和记载的一种手段。在各种科学手段十分发达的今天,观察依然是生态研究中的重要方法。目前生态学中的观察主要有野外观测(包括野外考察和定位观测)和实验室(包括试验田)观察两大类。 依照研究目的,使用科学仪器和设备,有意义地去控制自然过程条件,模拟自然现象,避开次要矛盾,突出主要因素,在特定条件下去探索客观规律,认识客观世界,这种方法即实验方法。生态学中的实验方法主要有原地实验和人工控制实验两类。原地实验是在野外条件下通过某些措施获得某个因素的变化对生物的影响及生物的响应数据。例如,在牧场进行围栏可以分析出食草动物对群落结构和生产力的影响;在田间人工“小岛”上接种昆虫,以观测昆虫出现后的生态关系变化,等等。人工控制实验是在受控条件系统中研究单项或多项因子对目标的作用,如人工气候箱中的实验。 在科学研究中实验只是手段,由观察、实验获得的大量第一手资料,需要经过比较与分类,进一步进行归纳、演绎、分析、综合,进行逻辑思维与抽象(经常要用到模型方法,进行逻辑思维与抽象,形成概念,提出假说,发展成理论。因此,生物科学的研究过程中不能只动手,而应该同时动脑。 生态学实验不是一般的验证实验,而是探索型实验。同学们必须对实验数据进行充分的比较\综合和分析,最后得出自己的结论。这期间需要阅读科学文献,共享其他成员和实验组的数据,并按照科学论文的格式进行写作。 迈开了解和参与科学研究的第一步,你的投入越多,你的收获越丰!
ARMA模型及分析 本次试验主要是通过等时间间隔,连续读取70个某次化学反应的过程数据,构成一个时间序列。试对该时间序列进行ARMA模型拟合以及模型的优化,最后进行预测。以下本次试验的数据: 表1 连续读取70个化学反应数据 47 64 23 71 38 64 55 41 59 48 71 35 57 40 58 44 80 55 37 74 51 57 50 60 45 57 50 45 25 59 50 71 56 74 50 58 45 54 36 54 48 55 45 57 50 62 44 64 43 52 38 59 55 41 53 49 34 35 54 45 68 38 50 60 39 59 40 57 54 23 资料来源:O’Donovan, Consec. Readings Batch Chemical Proces, https://www.doczj.com/doc/959630979.html,ler et al. 下面的分析及检验、预测均是基于上述数据进行的,本次试验是在Eviews 6.0上完成的。 一、序列预处理 由于只有对平稳的时间序列才能建立ARMA模型,因此在建立模型之前,有必要对序列进行预处理,主要包括了平稳性检验和纯随机检验。 序列时序图显示此化学反应过程无明显趋势或周期,波动稳定。见图1。
图2 化学反应过程相关图和Q统计量 从图2的序列的相关分析结果:1. 可以看出自相关系数始终在0周围波动,判定该序列为平稳时间序列2.看Q统计量的P值:该统计量的原假设为X的1期,2期……k期的自相关系数均等于0,备择假设为自相关系数中至少有一个不等于0,因此如图知,该P值在滞后2、3、4期是都为0,所以拒接原假设,即序列是非纯随机序列,即非白噪声序列(因为序列值之间彼此之间存在关联,所以说过去的行为对将来的发展有一定的影响,因此为非纯随机序列,即非白噪声序列)。 二、模型识别 由于检验出时间序列是平稳的,且是非白噪声序列,因此可以建立模型,在建立模型之前需要识别模型阶数即确定阶数。阶数确定要借助于时间序列的相关图,即序列的自相关函数和偏自相关函数,并根据他们之间的理论模式进行阶数最后的确定。 下面给出自相关函数和偏自相关函数之间的理论模式:
传染病问题中的SIR模型 摘要: 2003年春来历不明的SARS病毒突袭人间,给人们的生命财产带来极大的危害。长期以来,建立传染病的数学模型来描述传染病的传播过程,分析受感染人数的变化规律,探索制止传染病蔓延的手段等,一直是我国及全世界有关专家和官员关注的课题。 不同类型的传染病的传播过程有其各自不同的特点,我们不是从医学的角度一一分析各种传染病的传播,而是从一般的传播机理分析建立各种模型,如简单模型,SI模型,SIS 模型,SIR模型等。在这里我采用SIR(Susceptibles,Infectives,Recovered)模型来研究如天花,流感,肝炎,麻疹等治愈后均有很强的免疫力的传染病,它主要沿用由Kermack 与McKendrick在1927年采用动力学方法建立的模型。应用传染病动力学模型来描述疾病发展变化的过程和传播规律,预测疾病发生的状态,评估各种控制措施的效果,为预防控制疾病提供最优决策依据, 维护人类健康与社会经济发展。 关键字:传染病;动力学;SIR模型。 一﹑模型假设 在疾病传播期内所考察的地区范围不考虑人口的出生、死亡、流动等种群动力因素。总 人口数N(t)不变,人口始终保持一个常数N。人群分为以下三类: 易感染者(Susceptibles), 其数量比例记为s(t),表示t时刻未染病但有可能被该类疾病传染的人数占总人数的比例;感染病者(Infectives), 其数量比例记为i(t),表示t时刻已被感染成为病人而且具有传染力的人数占总人数的比例; 恢复者(Recovered),
其数量比例记为r(t),表示t时刻已从染病者中移出的人数(这部分人既非已感染者,也非感染病者,不具有传染性,也不会再次被感染,他们已退出该传染系统。)占总人数的比例。 病人的日接触率(每个病人每天有效接触的平均人数)为常数λ,日治愈率(每天被治愈的病人占总病人数的比例)为常数μ, 显然平均传染期为1/μ,传染期接触数为σ=λ/μ。 该模型的缺陷是结果常与实际有一定程度差距,这是因为模型中假设有效接触率传染力是不变的。 Logistic曲线模型: 如下为拟合的原始数据点:
种群数量增长的几种数学曲线模型例析 种群生态学研究的核心是种群的动态问题。种群增长是种群动态的主要表现形式之一,它是在不同环境条件下,种群数量随着时间的变化而增长的 状态。数学曲线模型能直观反映种群数量增长的规律,它能达到直接观察和实验所得不到的效果。为了更好理解种群数量增长规律,下面结合实例介绍种群数量增长的几种数学曲线模型。 1.种群数量增长曲线模型 种群在“无限”的环境中,即环境中空间、食物等资源是无限的,且气候适宜、没有天敌等理想条件下,种群的增长率不随种群本身的密 度而变化,种群数量增长通常呈指数增长。也就是说,种群数量每年以 一定的倍数增长,第二年的数量是第一年的λ倍,t年后种群数量为N t =N0λt,如果绘成坐标图指数式增长很像英文字母“J”,称之为“J” 型增长曲线。然而自然种群不可能长期地呈指数增长。当种群在一个有 限的环境中,随着密度的上升,个体间对有限的空间、食物和其他生活 条件的种内斗争也将加剧,加之天敌的捕食,疾病和不良气候条件等因 素必然要影响到种群的出生率和死亡率,从而降低了种群的实际增长率,一直到停止增长。种群在有限环境条件下连续增长称之为逻辑斯谛增长,这种增长曲线很像英文字母“S”,称之为“S”型增长曲线。两种类型种群增长模型如右图所示。 例1.右图为某种群在不同环境的增长曲线,据图判断下列说法不正确的是 ( D ) A.A曲线呈“J”型,B曲线呈“S”型 B.改善空间和资源有望使K值提高 C.阴影部分表示有环境阻力存在 D.种群数量达到K值时,种群增长最快 解析:由图可知,A曲线呈“J”型增长,B曲线呈“S”型增长。在种群生态学中,环境容纳量(K值)是指在环境条件不受破坏的情况下,一定空间中所能维持的种群最大数量。环境容纳量是一个动态的变量,只要生物或环境因素发生变化,环境容纳量也就会发生相应的变化。因此,改善空间和资源有望使K值提高。图像中阴影部分表示环境阻力所减少的生物个体数,代表环境阻力的大小。种群数量在k/2时增长速率最大。 2.种群λ值变化曲线模型 在种群指数式增长过程中,λ值表示相邻两年(生物的两代)种群数量的倍数。从理论上讲,λ有以下四种情况:λ>1 种群上升;λ=1 种群稳定;0<λ<1 种群下降;λ=0 种群无繁殖现象,且在一代中灭亡。
一、逻辑斯蒂方程建立的过程及背景 在自然界和社会上存在大量的 s型变化的现象, 逻辑斯蒂Logistic模型几乎是描述 s型增长的唯一数学模型.这是一条连 续的、单调递增的、以参数 k为上渐近线的 s型曲线, 其变化 速度一开始增长较慢, 中间段增长速度加快, 以后增长速度下 降并且趋于稳定. 利用它可以表征种群的数量动态, 描述某一 研究对象的增长过程, 也可作为其它复杂模型的理论基础如 Lotka- Volterra两种群竞争模型. 可以看出逻辑斯蒂方程不管 在自然科学领域还是在社会科学中都具有非常广泛的用途. 1逻辑斯蒂模型的产生与发展 在提出逻辑斯蒂模型之前, 最早给出种群生态学经典数学模型 是 M althus模型, 由英国统计学家 M althus( 1766- 1834)在 1798年人口原理!一书中, 提出了闻名于世的 M althus人口模 型. 设 t0时刻的人口总数为 N ( t0), t时刻人口总数为 N( t), 则: dN/dt=rN N(t0)=N0 但是这个模型有很大的局限性: 只考虑出生率和死亡率, 而没 有考虑环境因素. 实际上人类所生存的环境中资源并不是无限 的, 因而人口的增长也不可能是无限的, 实践证明 M althus人 口模型只符合人口的过去而不能用来预测未来人口总数. 比利 时数学家Verhulst对Malthus模型中关于人口增长率为常数这一 假设修改为 dNdt=rN-KN^2 N(t0)=N0 其中 r,K称为生命系数(VitalCoefficients). (2)式就 是最早的逻辑斯蒂模型. 解之得: N(t) =1/(K/r+(1/N0-K/r)exp(-rt) 二、逻辑斯蒂方程在MATLAB中的实现 function f = curvefun1(x,t) syms x t; k=9000;
第六次建模作业 一.logistic模型模拟 【摘要】物种种群数量的变化规律一直是我们所探究的问题,考虑到一些自然灾害和物种间的食物链或竞争关系,我们可以在一定条件下模拟某一种群的变化规律。对于人口的增长一直是一个热门话题,我 们通过数据的统计和拟合可以总结出某地区的人口变化规律,并在其他地区进行模型检验,分析该动态机理模型是否在一定程度上成立。 【关键词】人口增长数据统计模型检验动态机理模型 【问题重述】美国人口数据随时间的变化: 1790180018101820183018401850 3.9 5.37.29.612.917.123.2 I860187018801890190019101920 31.438.650.262.976.092.0106.5 19 301940195019601970198019902000 123132151179204227251281【模型建立】首先我们可用微积分的思想将连续的微分方程离散化, 不妨设x(n)表示第n次普查所得人口数,根据logistic模型 dy/dt=r(1-y/K)y 可得: (x(n +1) —x(n) )= r(1 —n) K x(n +1) -X(n) _ r(1x(n)) x(n) K 进一步化简有 U = x(n+1) -x(n) x( n) V = x(n)
rv u = r-—— K 可得: 【求解模型】现在我们可以用线性拟合,借助 matlab 来进行运算得 到r ,K X=[3.9 ; 5.3 7.2 ; 9.6 ; 50.2 ; 62.9 ; 76.0 ; 92.0 ; 204 ; 227 ; 251 ; 281]; Y=[] for i=1:21 23.2; 31.4; 38.6 ; 132 ; 151; 179 ; End Y=[Y ,Y(i)] 运行结果运用cftool 工具线性模拟: Result Lin ear model Poly1: f(x) = p 1*x + p2 Coefficie nts (with 95% con fide nee boun ds): p1 = -0.0009825 (-0.001254,-0.0007108) p2 = 0.3178 (0.2832, 0.3525) 运行程序: Y(i)=(X(i+1,:)-X(i,:))./(X(i,:)); 106.5; 123 ; 12.9; 17.1;
迪克西特-斯蒂格利茨模型及其应用 胡 怀 国 新贸易理论和新增长理论是当前经济理论中最热门的研究领域之一,它们的产生和发展在很大程度上则得益于迪克西特-斯蒂格利茨模型(简称D-S 模型)的启发,该模型是由迪克西特(A.K. Dixit,)和斯蒂格利茨在1977年所发表的《垄断竞争和最优产品的多样性》论文中提出来的。本文先简要介绍D-S 模型的核心内容,然后重点分析新贸易理论和新增长理论如何能够以D-S 模型为基础建立起来。 一、迪克西特-斯蒂格利茨模型 不论是在国际贸易理论还是在经济增长理论方面,人们日益发现“规模经济”是问题的核心。尽管人们可以在完全竞争的框架下对外部规模经济问题进行研究,但内部规模经济却无法和竞争性的市场结构相兼容。在D-S 模型提出以前,由于内部规模经济模型的求解极为复杂、且一般不能求出均衡解(从而更无法进行福利分析和比较),人们不得不借助于外部性、溢出效应和边干边学等似是而非的概念,将研究局限于外部规模经济的分析。不论是经济理论本身还是对经济现实的解释,都要求一种能够对内部规模经济和垄断竞争的市场结构进行严格分析的理论框架,D-S 模型则为该问题的解决提供了简洁的基本方法。 迪克西特和斯蒂格利茨在论文中指出,经济学中的一个基本问题是:一个市场解能否导致社会最优的产品数量和产品种类?分配不公、外部效应和规模经济都是导致不完全市场结构、并使得市场均衡解偏离社会最优解的原因。他们的主要目的,是对其中的(内部)规模经济情形进行分析。他们首先构造了著名的迪克西特-斯蒂格利茨效用函数(D-S 效用函数,后被引申为D-S 生产函数),然后依次在效用函数是不变弹性、可变弹性和非对称性的情形下求出其市场均衡解,并分别在每种情形下对市场均衡解和社会最优解进行了比较。 D-S 模型的核心内容体现在消费和生产两个方面:(1)他们通过构造一个包含产品种类的D-S 效用函数,推导出了特定形式的需求函数和张伯伦dd 曲线与DD 曲线;(2)他们假定每种产品的生产都具有不变的固定成本和边际成本(这意味着成本函数具有平均成本递减和边际成本不变的性质,从而呈现出内部规模经济的特征),然后结合需求函数和新厂商自由进入的条件,求得每个厂商的均衡产量、均衡价格和产品种类数量。 为了讨论产品数量和产品种类的关系,迪克西特和斯蒂格利茨首先将整个经济分为两部分:某个行业(集团或部门)和行业之外的其它部分。假定行业内部的各种产品之间具有很好的替代性,但与其它行业产品之间的替代性则很差。如果将行业之外的所有产品用0x 表示,行业内的各种产品用()n i x i ,,2,1 =表示,则代表性消费者的效用函数可以表示为 ?? ??????????? ??=∑ρρ10,i i x x U u ,其中()ρ-11是行业内部各种产品之间的替代弹性(这里假定效用函数对行业内的每种产品是对称的)。此外,为了保证效用函数的凸性,假定10<<ρ。 为了在I x p x n i i i =+∑=10的预算约束下,求出实现上述效用函数最大化的每种产品的最 优消费数量,他们构造了关于产品数量和价格水平的两个指数:ρρ11??????=∑=n i i x y 和
种群数量增长的几种数学曲线模型例析 吉林省梨树县第一高级中学姜万录 种群生态学 研究的核心是种 群的动态问题。种 群增长是种群动 态的主要表现形 式之一,它是在不 同环境条件下,种 群数量随着时间 的变化而增长的 状态。数学曲线模型能直观反映种群数量增长的规律,它能达到直接观察和实验所得不到的效果。为了更好理解种群数量增长规律,下面结合实例介绍种群数量增长的几种数学曲线模型。 1.种群数量增长曲线模型 种群在“无限”的环境中,即环境中空间、食物等资源是无限的,且气候适宜、没有天敌等理想条件下,种群的增长率不随种群本身的密度而变化,种群数量增长通常呈指数增长。也就是说,种群数量每年以一定的倍数增长,第二年的数量是第一年的λ倍,t年后种群数量为N t=N0λt,如果绘成坐标图指数式增长很像英文字母“J”,称之为“J”型增长曲线。然而自然种群不可能长期地呈指数增长。当种群在一个有限的环境中,随着密度的上升,个体间对有限的空间、食物和其他生活条件的种内斗争也将加剧,加之天敌的捕食,疾病和不良气候条件等因素必然要影响到种群的出生率和死亡率,从而降低了种群的实际增长率,一直到停止增长。种群在有限环境条件下连续增长称之为逻辑斯谛增长,这种增长曲线很像英文字母“S”,称之为“S”型增长曲线。两种类型种群增长模型如右图所示。 例1.右图为某种群在不同环境的增长曲线,据图判断下列说法不正确的是 ( D ) A.A曲线呈“J”型,B曲线呈“S”型 B.改善空间和资源有望使K值提高 C.阴影部分表示有环境阻力存在
D.种群数量达到K值时,种群增长最快 解析:由图可知,A曲线呈“J”型增长,B曲 线呈“S”型增长。在种群生态学中,环境容纳 量(K值)是指在环境条件不受破坏的情况下, 一定空间中所能维持的种群最大数量。环境容 纳量是一个动态的变量,只要生物或环境因素 发生变化,环境容纳量也就会发生相应的变化。 因此,改善空间和资源有望使K值提高。图像中阴 影部分表示环境阻力所减少的生物个体数,代表环境阻力的大小。种群数量在k/2时增长速率最大。 2.种群λ值变化曲线模型 在种群指数式增长过程中,λ值表示相邻两年(生物的两代)种群数量的倍数。从理论上讲,λ有以下四种情况:λ>1 种群上升;λ=1 种群稳定;0<λ<1 种群下降;λ=0 种群无繁殖现象,且在一代中灭亡。 例2.某岛屿引入外来物种野兔,研究人员调查了30年间野兔种群数量的变化,并据此绘制了值变化曲线(右图)。以下叙述正确的是(D) A.第1年至第5年间野兔种群数量保持相对稳定 B.第5年起野兔种群数量开始下降 C.第15年至第20年间野兔种群数量呈“J”型增长 D.第20年至第30年间野兔种群数量增长率为0 解析:由图示可知,λ值为当年种群数量与上一年种群数量的比值,种群增长率=λ—1,第1年至第5年间,λ大于1野兔种群数量逐年递增,从第5年起,λ值减少但大于0,增长率下降,野兔种群数量增加幅度减少,但不会下降。第15年至第20年间野兔种群数量随种群增长率先增加后减小,呈现“S”型增长。第20年至第30年间,λ值为1野兔种群数量增长率为0。 3.种群增长率和增长速率变化模型 种群增长率是指单位时间种群增长数量,种群增长率=出生率—死亡率=(出生数-死亡数)/(单位时间×单位数量)。种群在“J”型曲线中,增长率保持不变(如图A);而种群在“S”型曲线中,每年的增长率由最初的最大值,在环境阻力(空间压力、食物不足等)的作用下,导致出生率下降、死亡率上升,增长率越来越小,种群数量到达最大值(K值)时,其增长率为0(如图C);