
遮光后叶绿素含量升高和叶绿素a/b降低的原因 试题:如图,叶绿素的含量随着遮光比例的升高而升高,遮光后叶绿素a/b 降低,捕光能力上升。原因。 因为学生知道,光是叶绿素形成的必需条件,所以大部分学生都错误认为叶绿素含量随光照增强而增加。 从资料中可以看出,这些变化都是为了适应植物在遮光条件下的生长。 一、遮光后叶绿素含量为什么会升高 叶绿素含量受到光照、温度、矿质元素、逆境等外界因素及核基因、质基因等内在因素的共同影响,在外部因素中光对叶绿素的合成与分解起主导作用。植物体中叶绿素的合成和分解处于一个动态平衡中,叶片光照后,才能顺利地合成叶绿素,但形成叶绿素所要求光照强度相对较低,当然过弱也不利于叶绿素的生物合成,除680nm以上波长以外,可见光中各种波长的光照都能促使叶绿素形成,光过强反而会发生光氧化而受破坏。 植物中叶绿素和蛋白质结合为结合态叶绿素才能发挥作用,而自由态的叶绿素则会对细胞造成光氧化损伤。为了避免自由态叶绿素对细胞造成的光氧化损伤,植物必须快速降解这些物质。 在遮光条件下,集光色素蛋白在光合单位中的相对含量会增加,从而导致结合态叶绿素增加。与此同时,降低了叶绿素的降解和光氧化,所以遮光后叶绿素的含量会增加。 遮荫环境下,植物通过增加单位叶面积色素密度和叶绿素含量,有利于提高植株的捕光能力,吸收更多的光,提高光能利用率,是对弱光环境的一种适应。 二、遮光后叶绿素a/b降低 在不同生理条件下,叶绿素a和叶绿素b的合成、分解速度影响了叶绿素a/b的比值,但调节叶绿素a/b的比值主要通过“叶绿素循环”实现。叶绿素a 和叶绿素b的相互转化称为“叶绿素循环”。 在遮光条件下,叶绿素a向叶绿素b的转化加快,叶绿素a水解形成脱植基叶绿素a,脱植基叶绿素a再转化为脱植基叶绿素b,最后合成叶绿素b,从而降低了叶绿素a/b的比值。弱光下叶绿素b的相对含量增高是有其生理适应,有利于对弱光的利用。
部分叶绿素荧光动力学参数的定义: F0:固定荧光,初始荧光(minimalfluorescence)。也称基础荧光,0水平荧光,是光系统Ⅱ(PSⅡ)反应中心处于完全开放时的荧光产量,它与叶片叶绿素浓度有关。 Fm:最大荧光产量(maximalfluorescence),是PSⅡ反应中心处于完全关闭时的荧光产量。可反映经过PSⅡ的电子传递情况。通常叶片经暗适应20 min后测得。 F:任意时间实际荧光产量(actualfluorescence intensity at any time)。 Fa:稳态荧光产量(fluorescence instable state)。 Fm/F0:反映经过PSⅡ的电子传递情况。 Fv=Fm-F0:为可变荧光(variablefluorescence),反映了QA的还原情况。 Fv/Fm:是PSⅡ最大光化学量子产量(optimal/maximal photochemical efficiency of PSⅡin the dark)或(optimal/maximalquantum yield of PSⅡ),反映PSⅡ反应中心内禀光能转换效率(intrinsic PSⅡefficiency)或称最大PSⅡ的光能转换效率(optimal/maximalPSⅡefficiency),叶暗适应20 min后测得。非胁迫条件下该参数的变化极小,不受物种和生长条件的影响,胁迫条件下该参数明显下降。 Fv’/Fm’:PSⅡ有效光化学量子产量(photochemicalefficiency of PSⅡin the light),反映开放的PSⅡ反应中心原初光能捕获效率,叶片不经过暗适应在光下直接测得。 (Fm’-F)/Fm’或△F/Fm’:PSⅡ实际光化学量子产量(actual photochemical efficiency of PSⅡin the light)(Bilger和Bjrkman,1990),它反映PSⅡ反应中心在有部分关闭情况下的实际原初光能捕获效率,叶片不经过暗适应在光下直接测得。 荧光淬灭分两种:光化学淬灭和非光化学淬灭。光化学淬灭:以光化学淬灭系数代表:qP=(Fm’-F)/(Fm’-F0’);非光化学淬灭,有两种表示方法,NPQ=Fm/Fm’-1或qN=1-(Fm’-F0’)/(Fm-F0)=1-Fv’/Fv。 表观光合电子传递速率以[(Fm’-F)Fm’]×PFD表示,也可写成:△F/Fm’×PFD×0.5×0.84,其中系数0.5是因为一个电子传递需要吸收2个量子,而且光合作用包括两个光系统,系数0.84表示在入射的光量子中被吸收的占84%,PFD是光子通量密度;表观热耗散速率以(1-Fv’/Fm’)×PFD表示。 Fmr:可恢复的最大荧光产量,它的获得是在荧光P峰和M峰后,当开放的PSⅡ最大荧光产量平稳时,关闭作用光得到F0’后,把饱和光的闪光间隔期延长到180s/次,得到一组逐渐增大(对数增长)的最大荧光产量,将该组最大荧光产量放在半对数坐标系中即成直线,该直线在Y轴的截距即为Fmr。以(Fm-Fmr)/Fmr可以反映不可逆的非光化学淬灭产率,即发生光抑制的可能程度。 FO(初始荧光),Fm(最大荧光),Fv= Fm-FO(可变荧光),Fv /Fm(PSII最大光化学效率或原初光能转换效率),Fv /FO(PSII的潜在活性),Yield(PSII总的光化学量子产额),ETR(表观电子传递速率),PAR(光合有效辐射),LT(叶面温度)。其中FO、Fm、Fv /FO测定前将叶片暗适应20 min。各参数日变化从6: 00~18: 00,每2h测定一次。 (Fv /Fm)和(Fv /FO)分别用于度量植物叶片PSII原初光能转换效率和PSII潜在活性,-(Yield)是PSII的实际光化学效率,反映叶片用于光合电子传递的能量占所吸收光能的比例,是PSII反应中心部分关闭时的光化学效率,其值大小可以反映PSII反应中心的开放程度。常用来表示植物光合作用电子传递的量子产额,可作为植物叶片光合电子传递速率快慢的相对指标。即在光合作用进程中,PSII每获得一个光量子所能引起的总的光化学反应。因此,较高的Yield值,有利于提高光能转化效率,为暗反应的光合碳同化积累更多所需的能量,以促进碳同化的高效运转和有机物的积累。同样毛蕊红山茶和长毛红山茶的Yield值也较高。
实验报告 课程名称: 植物生理学(乙)指导老师: 廖敏 成绩: 实验名称: 叶绿素理化性质和含量 实验类型: 定量探究型 同组学生姓名: 方昊 一、实验目的和要求(必填) 三、主要仪器设备(必填) 五、实验数据记录和处理 七、讨论、心得 二、实验内容和原理(必填) 四、操作方法和实验步骤 六、实验结果与分析(必填) 一、实验目的和要求 掌握植物中叶绿体色素的分离和性质鉴定、定量分析的原理和方法; 二、实验内容和原理 以青菜为材料,提取和分离叶绿体色素并进行理化性质测定和叶绿素含量 分析。原理如下: 1. 叶绿素和类胡萝卜素均不溶于水而溶于有机溶剂,常用95%的乙醇或80%的丙酮提取; 2. 叶绿素是二羧酸酯,与强碱反应,形成绿色的可溶性叶绿素盐,就可与有机溶剂中的类胡萝卜素 分开; 3. 在酸性或加温条件下,叶绿素卟啉环中的Mg++可依次被H+和Cu++取代形成褐色的去镁叶绿素和绿色的铜代叶绿素; 4. 叶绿素受光激发,可发出红色荧光,反射光下可见红色荧光; 5. 叶绿素吸收红光和蓝紫光,红光区可用于定量分析,其中645和663用于定量叶绿素a 、b 及总量,而652可直接用于总量分析。 专业:农业资源与环境 姓名: 吴主光 学号: 3110100403 日期: 2013.10.17 地点: 生物实验中心 装 订 线
三、主要仪器设备 1. 天平(万分之一)、可扫描分光光度计、离心机、研具、各种容(量)器、洒精灯等 四、操作方法、实验步骤以及实验现象 定性分析: 鲜叶5g+95%30ml(逐步加入),磨成匀浆 过滤入三角瓶中,观察荧光现象:透射光绿色,反射光红色。 皂化反应(3ml):加KOH数片剧烈摇均,加石油醚5ml和H2O1ml分层后观察:上层呈黄色,为类胡萝卜素,吸收蓝紫光;下层呈绿色,为叶绿素,吸收红光和蓝紫光。 取代反应(1):加醋酸约2ml,变褐(去镁叶绿素);取1/2加醋酸铜粉加热,变鲜绿色,为铜代叶绿素。 取代反应(2):鲜叶2-3cm2,加Ac-AcCu 20ml加热,观察: 3 min变为褐绿色的去镁叶绿素, 5 min后,变为深绿色的铜代叶绿素。 叶绿素和类胡萝卜素的吸收光谱测定: 皂化反应的上层黄色石油醚溶液(稀释470nm OD 0.5-1) 反复用石油醚粹取,直到无类胡萝卜素,离心得叶绿素(盐)(稀释663nm OD 0.5-1) 在400-700nm处扫描光谱,分别测定类胡萝卜素和叶绿素的吸收峰. 叶绿素定量分析:鲜叶0.1g,加1.9mlH2O,磨成匀浆,取0.2ml加80%丙酮4.8ml,摇匀,4000转离心3min,上清液在645,652,663测定OD,计算Chla,Chlb 和Chl总量的值。 五、实验数据记录和处理
一、实验课题名称:不同环境条件下植物叶绿素a、b含量的比较 二、选题背景或文献综述: 《植物生理学实验指导》(第四版)、《植物生理学》(第六版)、上网查阅相关资料 阴生植物也称“阴性植物”,是在较弱的光照条件下生长良好的植物,但并不是阴生植物对光照强度的要求越弱越好,而是必须达到阴生植物的补偿点,植物才能正常生长,阳生植物也称“阳性植物”,光照强度对植物的生长发育及形态结构的形成有重要作用,在强光环境中生长发育健壮,在阴蔽和弱光条件下生长发育不良的植物称阳性植物,这类植物要求全日照,并且在水分、温度等条件适合的情况下,不存在光照过强的问题。 阳生植物和阴生植物的区别:关于光的饱和点和补偿点光是光合作用的能量来源,光照强度直接影响光合速率,在其它条件都适宜的情况下,在一定范围内,光合速率随光照强度提高而加快,当光照强度高到一定数值后,光照强度再提高而光合速率不再加快,这种现象叫光饱和现象。开始达到光饱和现象的光照强度称为光饱和点,在光饱和点以下,随着光照强度减弱,光合速率减慢,当减弱到一定光照强度时,光合作用吸收二氧化碳量与呼吸释放二氧化碳的量处于动态平衡,这时的光照强度称为光补偿点。此时植物制造有机物量和消耗有机物量相等,不同类型植物的光饱和点和
补偿点是不同的,阳性植物的光饱和点和补偿点一般都高于阴性植物。 结构和特性的区别:阴生植物的叶片的疏导组织比阳生植物稀疏,以叶绿体来说,阳生植物有较大的基粒,基粒片层数目多的多,叶绿素含量也高,阴生植物在较低的光照条件下充分的吸收光线,叶绿素a/叶绿素b的比值小,能够强烈的利用蓝紫光,阳性植物叶片小而厚,表面具蜡质或绒毛,叶脉密,单位面积内气孔多,叶绿素含量高,体内含盐分多,渗透压高,可以抗高温干旱,阳生植物的气孔一般在叶片下表皮分布的数量多于上表皮,这样可以避免阳光直晒而减少水分散失,阳生植物的呼吸速率高于阴生植物。 区分阳生植物与阴生植物,主要是根据植物对光照强度需要的不同,阳生植物要求充分直射日光才能生长或生长良好,阴生植物适宜于生长在荫蔽环境中,它们在完全日照下反而生长不良或不能生长,阳生植物和阴生植物之所以能适应不同光照,是与它们的生理特征和形态特征不同有关,以光饱和点来说,阳生植物的光饱合点是全光照(即全部太阳光照)的100%,而阴生植物是全光照的10%~50%。因为阴生植物叶片的输导组织比阳生植物的稀疏,当光照强度增大时,水分对叶片的供给不足,阴生植物便不再增加光合速率,以叶绿体来说,阴生植物与阳生植物相比,前者有较大的基粒,基粒片层数目多,叶绿素含量较高,能在较低光照强度下充分
盐胁迫对植物的影响 植物的抗盐性: 我国长江以北以及沿海许多地区,土壤中盐碱含量往往过高,对植物造成危害。这种由于土壤盐碱含量过高对植物造成的危害称为盐害,植物对盐害的适应能力叫抗盐性。根据许多研究报道,土壤含盐量超过0.2%~0.25%时就会造成危害。钠盐是形成盐分过多的主要盐类,习惯上把硫酸钠与碳酸钠含量较高的土壤叫盐土,但二者同时存在,不能绝对划分,实际上把盐分过多的土壤统称为碱土。世界上盐碱土面积很大,估计占灌溉农田的1/3,约4×107ha,而且随着灌溉农业的发展,盐碱面积将继续扩大。我国盐碱土主要分布于西北、华北、东北和海滨地区,盐碱土总面积约2~7×107ha,而且这些地区都属平原,盐地土层深厚,如能改良盐碱危害,发展农业的潜力很大,特别应值得重视。 土壤盐分过多对植物的危害: 1.生理干旱:土壤中可溶性盐类过多,由于渗透势增高而使土壤水势降低,根据水从高水势向低水势流动的原理,根细胞的水势必须低于周围介质的水势才能吸水,所以土壤盐分愈多根吸水愈困难,甚至植株体内水分有外渗的危险。因而盐害的通常表现实际上是旱害,尤其在大气相对湿度低的情况下,随蒸腾作用加强,盐害更为严重,一般作物在湿季耐盐性增强。 2.离子的毒害作用:在盐分过多的土壤中植物生长不良的原因,不完全是生理干旱或吸水困难,而是由于吸收某种盐类过多而排斥了对另一些营养元素的吸收,产生了类似单盐毒害的作用。 3.破坏正常代谢:盐分过多对光合作用、呼吸作用和蛋白质代谢影响很大。盐分过多会抑制叶绿素生物合成和各种酶的产生,尤其是影响叶绿素-蛋白复合体的形成。盐分过多还会使PEP羧化酶与RuBP 羧化酶活性降低,使光呼吸加强。生长在盐分过多的土壤中的作物(棉花、蚕豆、番茄等),其净光合速率一般低于淡土的植物,不过盐分过多对光合作用的影响是初期明显降低,而后又逐渐恢复,这似乎是一种适应性变化。盐分过多对呼吸的影响,多数情况下表现为呼吸作用降低,也有些植物增加盐分具有提高呼吸的效应,如小麦的根。呼吸增高是由于Na+活化了离子转移系统,尤其是对质膜上的Na+、K+与A TP活化,刺激了呼吸作用。盐分过多对植物的光合与呼吸的影响尽管不一致,但总的趋势是呼吸消耗增多,净光合速度降低,不利于生长。 一、实验目的 盐胁迫对植物生长发育的各个阶段都有不同程度的影响,如种子萌发、幼苗生长、成株生长等。不同种类的植物受盐胁迫影响的程度也各不相同。本实验主要观察Na2CO3对小麦种子萌发过程的影响,探讨小麦种子在盐胁迫下的萌发特性,对小麦的耐盐能力做出了初步评价。通过实验了解盐胁迫对植物(种子萌发)的影响;掌握种子萌发过程中发芽率、发芽势、发芽指数、芽长、总长、芽重、总重等各项指标的观察和计算方法;各项指标在盐胁迫条件下的变化趋势,绘制盐浓度与生长指标相关曲线,并分析盐胁迫对种子萌发的影响。 二、仪器设备和材料 电子天平;培养皿(直径120mm),滤纸(直径125mm定量滤纸若干),500ml、200ml烧杯,250ml 容量瓶,10ml移液管,玻璃棒,镊子,毫米刻度尺,剪刀;次氯酸钠、碳酸钠;小麦种子等。 三、实验方法和步骤 1.预处理 (1)种子的预处理:用10%的次氯酸钠消毒10min,蒸馏水冲洗数次后,于培养皿中做发芽实验。
第一节 叶绿素荧光参数及其意义 韩志国,吕中贤(泽泉开放实验室,上海泽泉科技有限公司,上海,200333) 叶绿素荧光技术作为光合作用的经典测量方法,已经成为藻类生理生态研究领域功能最强大、使用最 广泛的技术之一。由于常温常压下叶绿素荧光主要来源于光系统II 的叶绿素a ,而光系统II 处于整个光合 作用过程的最上游,因此包括光反应和暗反应在内的多数光合过程的变化都会反馈给光系统II ,进而引起 叶绿素a 荧光的变化,也就是说几乎所有光合作用过程的变化都可通过叶绿素荧光反映出来。与其它测量 方法相比,叶绿素荧光技术还具有不需破碎细胞、简便、快捷、可靠等特性,因此在国际上得到了广泛的 应用。 1 叶绿素荧光的来源 藻细胞内的叶绿素分子既可以直接捕获光能,也可以间接获取其它捕光色素(如类胡萝卜素)传递来 的能量。叶绿素分子得到能量后,会从基态(低能态)跃迁到激发态(高能态)。根据吸收的能量多少, 叶绿素分子可以跃迁到不同能级的激发态。若叶绿素分子吸收蓝光,则跃迁到较高激发态;若叶绿素分析 吸收红光,则跃迁到最低激发态。处于较高激发态的叶绿素分子很不稳定,会在几百飞秒(fs ,1 fs=10-15 s )内通过振动弛豫向周围环境辐射热量,回到最低激发态(图1)。而最低激发态的叶绿素分子可以稳定 存在几纳秒(ns ,1 ns=10-9 s )。 波长吸收荧光红 B 蓝 荧光 热耗散 最低激发态较高激发态基态吸收蓝光吸收红光能量A 图1 叶绿素吸收光能后能级变化(A )和对应的吸收光谱(B )(引自韩博平 et al., 2003) 处于最低激发态的叶绿素分子可以通过几种途径(图2)释放能量回到基态(韩博平 et al., 2003; Schreiber, 2004):1)将能量在一系列叶绿素分子之间传递,最后传递给反应中心叶绿素a ,用于进行光化 学反应;2)以热的形式将能量耗散掉,即非辐射能量耗散(热耗散);3)放出荧光。这三个途径相互竞 争、此消彼长,往往是具有最大速率的途径处于支配地位。一般而言,叶绿素荧光发生在纳秒级,而光化 学反应发射在皮秒级(ps ,1 ps=10-12 s ),因此在正常生理状态下(室温下),捕光色素吸收的能量主要用 于进行光化学反应,荧光只占约3%~5%(Krause and Weis, 1991; 林世青 et al., 1992)。 在活体细胞内,由于激发能从叶绿素b 到叶绿素a 的传递几乎达到100%的效率,因此基本检测不到 叶绿素b 荧光。在常温常压下,光系统I 的叶绿素a 发出的荧光很弱,基本可以忽略不计,对光系统I 叶 绿素a 荧光的研究要在77 K 的低温下进行。因此,当我们谈到活体叶绿素荧光时,其实指的是来自光系 统II 的叶绿素a 发出的荧光。
叶绿素含量的测定 一、原理 根据叶绿体色素提取液对可见光谱的吸收,利用分光光度计在某一特定波长测定其吸光度,即可用公式计算出提取液中各色素的含量。 根据朗伯—比尔定律,某有色溶液的吸光度A 与其中溶质浓度C 和液层厚度L 成正比,即A =αCL 式中:α比例常数。当溶液浓度以百分浓度为单位,液层厚度为1cm 时,α为该物质的吸光系数。各种有色物质溶液在不同波长下的吸光系数可通过测定已知浓度的纯物质在不同波长下的吸光度而求得。 如果溶液中有数种吸光物质,则此混合液在某一波长下的总吸光度等于各组分在相应波长下吸光度的总和。这就是吸光度的加和性。今欲测定叶绿体色素混合提取液中叶绿素a 、b 和类胡萝卜素的含量,只需测定该提取液在三个特定波长下的吸光度A ,并根据叶绿素a 、b 及类胡萝卜素在该波长下的吸光系数即可求出其浓度。在测定叶绿素a 、b 时为了排除类胡萝卜素的干扰,所用单色光的波长选择叶绿素在红光区的最大吸收峰。 已知叶绿素a 、叶绿素b 的80%丙酮溶液在红外区的最大吸收峰分别位于663、645nm 处。已知在波长663nm 下叶绿素a 、叶绿素b 在该溶液中的吸光系数的分别为82.04和9.27;在波长645nm 处的吸光系数分别为16.75和45.60。根据加和性原则列出以下关系式: A663=82.04Ca+9.27Cb (1) A645=16.76Ca+45.60Cb (2) 式(1) (2)A 663nm 和A645nm 为叶绿素溶液在663nm 和645nm 处的吸光度,C a C b 分别为叶绿素a 、叶绿素b 的浓度,以mg/L 为单位。 解方程(1) (2)组得 C a =12.72 A 663—2.59 A 645 (3) C b =22.88 A 645—4.67 A 663 (4) 将C a +C b 相加即得叶绿素总量C T C T = C a 十C b =20.29A 645—8.05 A 663 (5) 从公式(3)、(4)、(5)可以看出,,就可计算出提取液中的叶绿素a 、b 浓度另外,由于叶绿素a 叶绿素b 在652nm 的吸收峰相交,两者有相同的吸光系数(均为30.5),也可以在此波长下测定一次吸光度(A 652)而求出叶绿素a 、叶绿素 b 总量 所测定材料的单位面积或单位重量的叶绿素含量可按下式进行计算: C T = 5 .341000 652 A (6) 有叶绿素存在的条件下,用分光光度法可同时测出溶液中类胡萝卜素的含量。Licht-enthaler 等对Arnon 进行了修正,提出了 80%丙酮提取液中3种色素含量的计算公式: C a =12.21A 663—2.59 A 646 (7)
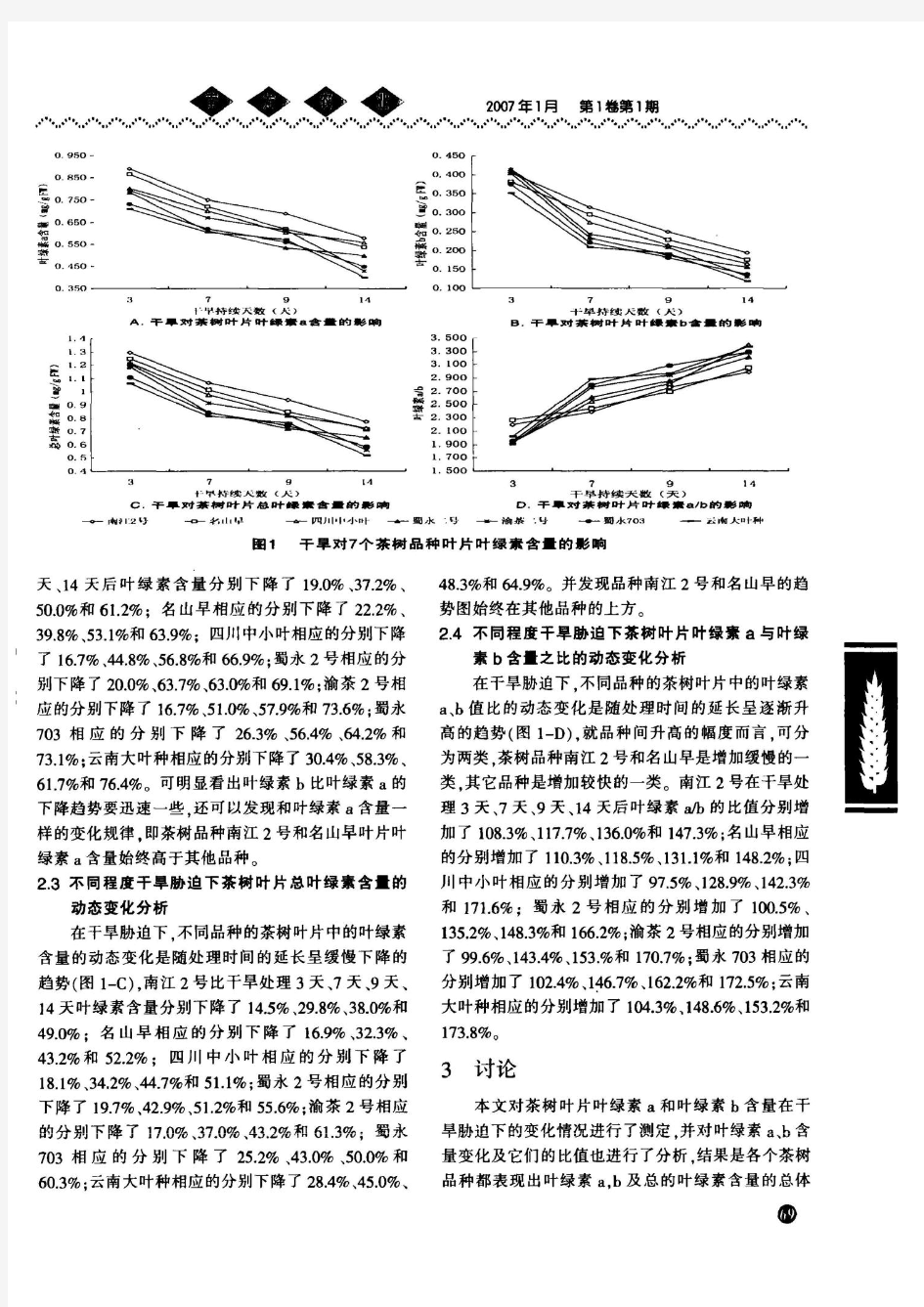
一、实验课题名称 不同环境条件下植物叶绿素a、b含量的比较(分光光度法测定) 二、文献综述 1.叶绿素a的生物合成过程 起始物是谷氨酸,之后为5-氨基酮戊酸,两分子的ALA缩合形成胆色素原(PBG),4分子PBG相互连结形成原中卟啉IX.原卟啉IX与Mg结合形成Mg-原卟啉原IX,光下E环的环化形成,D环的还原作用和叶绿醇尾部的连接完成了整个合成过程,合成过程中的许多步骤在图中已省略 2.影响叶绿素形成的条件 (1)光光是影响叶绿素形成的主要条件。从原叶绿素酸酯转变为叶绿酸酯需要光,而光过强,叶绿素又会受光氧化而破坏。黑暗中生长的幼苗呈黄白色,遮光或埋在土中的茎叶也呈黄白色。这种因缺乏某些条件而影响叶绿素形成,使叶子发黄的现象,称为黄化现象(etiolation)。 也有例外情况,例如藻类、苔藓、蕨类和松柏科植物在黑暗中可合成叶绿素,其数量当然不如在光下形成的多;柑橘种子的子叶及莲子的胚芽在无光照的条件下也能形成叶绿素,推测这些植物中存在可代替可见光促进叶绿素合成的生物物质。 (2)温度叶绿素的生物合成是一系列酶促反应,受温度影响。叶绿素形成的最低温度约2℃,最适温度约30℃,最高温度约40℃。秋天叶子变黄和早春寒潮过后秧苗变白,都与低温抑制叶绿素形成有关。高温下叶绿素分解大于合成,因而夏天绿叶蔬菜存放不到一天就变黄;相反,温度较低时,叶绿素解体慢,这也是低温保鲜的原因之一。 (3)营养元素叶绿素的形成必须有一定的营养元素。氮和镁是叶绿素的组成成分,铁、锰、铜、锌等则在叶绿素的生物合成过程中有催化功能或其它间接作用。因此,缺少这些元素时都会引起缺绿症(chlorosis),其中尤以氮的影响最大,因而叶色的深浅可作为衡量植株体内氮素水平高低的标志。 (4)氧缺氧能引起Mg-原卟啉IX或Mg-原卟啉甲酯的积累,影响叶绿素的合成。 (5)水缺水不但影响叶绿素生物合成,而且还促使原有叶绿素加速分解,所以干旱时叶片呈黄褐色。 通过对室外旱池处理条件下的甘薯叶片叶绿素含量变化的研究,结果表明,水分胁迫下甘薯品种叶片中叶绿素a、b及总叶
叶绿素荧光研究背景知识介绍 前言 近些年来,叶绿素荧光技术已经逐渐成为植物生理生态研究的热门方向。荧光数据是植物光合性能方面的必要研究内容。目前这种趋势由于叶绿素荧光检测仪的改进而得到发展。然而荧光理论和数据解释仍然比较复杂。就我们所了解的情况来看,目前许多研究者对荧光理论不是很清楚,仪器应用仅仅限于简单的数据说明的基础上,本文在此基础上,目的在于简单明晰地介绍相关理论和研究要点,以求简单明确地使用叶绿素荧光检测设备,充分分析实验数据,重点在于植物生理生态学技术的应用和限制。 荧光测量基础 植物叶片所吸收的光的能量有三个走向:光合驱动、热能、叶绿素荧光。三个过程之间存在竞争,其中任何一个效率的增加都将造成另外两个产量的下降。因此,测量叶绿素荧光产量,我们可以获得光化学过程与热耗散的效率的变化信息。尽管叶绿素荧光的总量很小(一般仅占叶片吸收光能总量的1-2%),测量却非常简单。荧光光谱不同于吸收光谱,其波长更长,因此荧光测量可以通过把叶片经过给定波长的光线的照射,同时测量发射光中波长较长的部分光线的量来实现。有一点需要注意的是,这种测量永远是相对的,因为光线不可避免会有损失。因此,所有分析必须把数据进行标准化处理,包括其进一步计算的许多参数也是如此。 调制荧光仪的出现是荧光研究技术的革命性的创新。在这类仪器中,测量光源是调制(高频率开关)的,其检测器也被调谐来仅仅检测被测量光激发的荧光。因此,相对的荧光产量可以在背景光线(主要是指野外全光照的条件下)存在的条件下进行测量。目前绝大多数的荧光仪采用了调制系统,同时也强烈建议选择调制荧光仪(Kate Maxwell,2000)。 为什么荧光产量会发生改变?Kautsky效应和Beyond 叶绿素荧光产量的变化最早在1960年被Kautsky和其合作者发现。他们发现,当把植物叶片从黑暗中转入光下,荧光产量瞬间上升(大约在1秒左右)这种上升可以解释为光合途径中电子受体的还原(可接受电子的受体的减少)。一旦PSII吸收光能,初级电子受体Q A(质体醌)接受了电子,它将不能再接受电子,直到它把电子传递给下一级电子载体Q B。此期间,反应中心是关闭的,反应中心关闭的比
叶绿素含量的测定 绿素含量的绿定叶 一、绿绿目的 1.了解分光光度绿的工作原理~ 2.掌握不同型分光光度绿的操作方绿~号 3.通绿本绿绿的绿掌握绿素含量绿定的一绿常绿的方法学叶------分光光度法。 二、绿绿原理 叶叶体体体叶绿素是脂溶性色素~主要存在于以绿绿首的色素中。在活中~绿绿 素脂蛋白绿合受到绿原系绿的保绿~绿和光是绿定的。与并氧 叶绿素的80%丙绿提取液在波绿663nm~645nm有吸收峰~绿素叶a和绿素叶b 的绿度符合以下公式, C=0.0127A-0.00259A a663645 C=0.0229A-0.00467A绿度绿位是,g/Lb645663 C=12.7A-2.59Aa663645 C=22.9A-4.67A绿度绿位是,mg/Lb645663 叶绿素绿绿度绿, C=C+CTab 若以绿液中色素含量表示~绿来 三、绿器、绿绿和材料 1.绿器 紫外-可绿分光光度绿、、研体25ml容量、璃漏斗、璃棒、皮绿滴管瓶玻玻2. 绿绿
丙绿;分析绿,、85%丙绿、80%丙绿 2.材料 绿绿、石英砂、酸绿碳 四、操作步绿 1. 在遮光件下取出等绿绿品~剪碎~混~取绿绿条匀称0.1-0.5g~ 2. 绿品置于绿~加入少量酸绿和石英砂~加入一定绿的丙绿磨绿绿~再加研内碳体研匀 85%丙绿适量绿绿磨至绿绿白色~研 3. 绿绿有绿绿的漏斗绿液绿入将匀25ml的容量中~用瓶并80%的丙绿分次洗绿和绿绿清研~ 最后用80%的丙绿定容。 4. 以80%的丙绿绿比液~在参663和645nm波绿绿绿定吸光绿;A绿在0.2-0.8范绿~内 绿度绿大绿用80%丙绿适稀绿,。当 五、绿果绿理 按照公式绿算出绿素叶a和绿素叶b的绿度~再绿算出绿素的含量。叶六、 注意事绿 1. 在活~绿合绿绿素是绿定的~绿绿一绿破~绿素易被光解。因此~抽提和绿体内叶坏叶 定工作绿可能避光快速完成。尽 2. 绿含有大量酸性液泡的绿品~绿首先加入微性的绿液~仔绿磨后加入丙绿绿行碱冲研抽提。 3. 分光光度绿的精度绿绿定的绿果有至绿重要的影~使用前绿绿器绿行校正。响七、思考绿
盐分胁迫对植物生长生理的影响 张华新,刘正祥等研究了光叶漆、银水牛果等11种树种后发现,盐胁迫后,各树种的苗高生长量下降、生物量累积减少,且随着处理浓度的增加均呈下降趋势,,各树种的根冠比值增大1 王润贤,周兴元,葛晋纲等人对草的研究后发现,在草坪草适应范围之内,根系活力和蛋白质含量呈先升后降的趋势,如超过忍受范围则持续下降。随盐分胁迫强度的增加和胁迫时间的延长,草坪草叶片的WSD上升,脯氮酸含量均表现为先升后降的趋势,但因胁迫程度和草种的不同,其峰值和下降幅度有较大差异。各项生理指标变化的趋势因草种的不同而有较大的差异,与其耐盐性有关,可以作为判定草坪草抗盐能力的评定依据。2 孙方行,李国雷对刺槐进行3天和17天盐胁迫处理后发现,MDA含量和细胞膜透性存在极显著正相关。叶绿素浓度和可溶性蛋白含量也存在极显著关。SOD活性和叶绿素浓度成负相关。从逐步回归分析可以看出细胞膜透性是影响高生长的主要指标3 张金香,钱金娥等人发现,经过前处理的1/2海水区中生长的苗木其叶、茎、根的生长量均超过淡水区中生长的苗木。说明一定程度的耐盐锻炼能够增强苗木对盐碱、干旱环境的适应能力4 张士功,高吉寅,宋景芝发现,6-苄基腺嘌呤、水杨酸、阿斯匹林,硝酸钙能够在一定程度上限制幼苗对Na+的吸收,阻滞其向地上部分运输的数量和速度。提高体内K+含量、向上运输效率,降低地上部分对Na+、K+的选择性(SNa+、K+>,同时6-苄基腺嘌呤还能够促进幼苗根系对Cl-的吸收,并有效地将Cl-限制在根部,阻滞Cl-向上运输,相对降低地上部分的Cl,这些都有利于
提高小麦幼苗抗盐性和对盐分胁迫的适应性5 王强,石伟勇,符建荣,指出,叶面喷施海藻液肥能提高黄瓜根冠比和干物质含量,提高根系总吸收面积和活跃吸收面积。不同浓度的海藻液肥均能降低盐胁迫对叶片质膜的伤害,提高SOD、POD等酶的活性,降低膜脂过氧化产物MDA的积累,提高脯氨酸、可溶性糖、可溶性蛋白等渗透调节物质的含量6 许兴,郑国琦.等指出,在等渗条件下,NaCl胁迫引起的小麦叶片组织含水量的下降、胁迫伤害率的增大及叶片和根部的脯氨酸、可溶性糖、Na+、K+含量的增加,均大于PEG胁迫引起的变化7 郑国琦,许兴,徐兆桢研究了盐分胁迫对植物的伤害和探讨了植物的耐盐的生物学机理以及通过基于改良作物耐盐性的研究进程。8 吴忠东,王全九.研究发现,在不同的生育期降水量条件下,冬小麦对盐分胁迫有着不同的响应。生育期一般年和湿润年可以采用的最高矿化度为3 g/L,而在生育期偏旱年,如果不采取其他措施的条件下,可以采用的最高矿化度为2 g/L,该结果为合理开发利用当地的地下咸水资源提供了一定的依据。9 郭淑霞,龚元石在研究盐分胁迫对菠菜生长和吸氮量的影响后发现,对菠菜进行盐分胁迫,前 44 天,随着盐分胁迫程度增加,菠菜相对生长速率 叶绿素含量的测定 一.实验原理 根据叶绿体色素提取液对可见光谱的吸收,利用分光光度计在某一特定波长测定其吸光度,即可用公式计算出提取液中各色素的含量。 根据朗伯—比尔定律,某有色溶液的吸光度A与其中溶质浓度C和液层厚度L成正比,即A=αCL.式中:α比例常数。当溶液浓度以百分浓度为单位,液层厚度为1cm时,α为该物质的吸光系数。各种有色物质溶液在不同波长下的吸光系数可通过测定已知浓度的纯物质在不同波长下的吸光度而求得。 如果溶液中有数种吸光物质,则此混合液在某一波长下的总吸光度等于各组分在相应波长下吸光度的总和。就是吸光度的加和性。如欲测定叶绿体色素混合提取液中叶绿素a、b和类胡萝卜素的含量,只需测定该提取液在三特定波长下的吸光度A,并根据叶绿素a、b 及类胡萝卜素在该波长下的吸光系数即可求出其浓度。在测定叶绿素a、b时为了排除类胡萝卜素的干扰,所用单色光的波长选择叶绿素在红光区的最大吸收峰。 植物叶绿素含量测定----丙酮提取法 高等植物光合作用过程中利用的光能是通过叶绿体色素(光合色素)吸收的。叶绿体色素由叶绿素a、叶绿素b、胡萝卜素和叶黄素组成。叶绿体色素的提取、分离和测定是研究它们的特性以及在光合中作用的第一步。叶片叶绿素含量与光合作用密切相关,是反眏叶片生理状态的重要指标。在植物光合生理、发育生理和抗性生理研究中经常需要测定叶绿素含量。叶绿素含量也是指导作物栽培生产和选育作物品种的重要指标。 ● 叶绿素不溶于水,溶于有机溶剂,可用多种有机溶剂,如丙酮、乙醇或二甲基亚砜等研磨提取或浸泡提取。叶绿色素在特定提取溶液中对特定波长的光有最大吸收,用分光光度计测定在该波长下叶绿素溶液的吸光度(也称为光密度),再根据叶绿素在该波长下的吸收系数即可计算叶绿素含量。 ●利用分光光计测定叶绿素含量的依据是Lambert-Beer定律,即当一束单色光通过溶液时,溶液的吸光度与溶液的浓度和液层厚度的乘积成正比。其数学表达式为: ●A=Kbc 式中:A为吸光度;K为吸光系数;b为溶液的厚度;c为溶液浓度。 ●叶绿素a、b的丙酮溶液在可见光范围内的最大吸收峰分别位于663、645nm处。叶绿素a 和b在663nm处的吸光系数(当溶液厚度为1cm,叶绿素浓度为g·L-1时的吸光度)分别为82.04和9.27;在645nm处的吸光系数分别为16.75和45.60。根据Lambert-Beer定律,叶绿素溶液在663nm和645nm处的吸光度(A663和A645)与溶液中叶绿素a、b和总浓度(a+b)(Ca、Cb 、Ca十b,单位为g·L-1),的关系可分别用下列方程式表示: ●A663=82.04C a+9.27C b (1) ●A645=16.76C a+45.60C b(2) ●C a=12.7 A663—2.59 A645(3) ●C b=22.9 A645—4.67 A663 (4) ●C a十b=20.3 A645—8.04 A663 (5) ● 植物对盐胁迫的反应 植物对盐胁迫的反应及其抗盐机理研究进展 杨晓慧1,2,蒋卫杰1*,魏珉2,余宏军1 (1.中国农业科学院蔬菜花卉研究所,北京100081;2.山东农业大学园艺科学与工程学院,山东泰安271018) REVIEW ON PLANT RESPONSE AND RESISTANCE MECHANISM TO SALT STRESS YANG Xiao-hui1,2,JIANG Wei-jie1*,WEI Min2,YU Hong-jun1( 1.Institute of Vegetables and Flowers,Chinese Academy of Agricultural Science,Beijing100081,China;2.College of Horticulture Science and Engineering,Shandong Agriculture University,Taian 271018,China) Key words:Iron stress,Osmotic stress,Salt resistant mechanism,Plant 摘要:本文从植物形态发育、质膜透性、光合和呼吸作用以及能量代谢等方面概述了盐胁迫下植物的生理生化反应,分析了盐害条件下离子胁迫和渗透胁迫作用机理以及植物的耐盐机制:植物小分子物质的积累、离子摄入和区域化、基因表达和大分子蛋白质的合成等,并简要综述了植物抗盐的分子生物学研究进展。 关键词:离子胁迫;渗透胁迫;耐盐机制;植物 中图分类号:S601文献标识码:A文章编号:1000-2324(2006) 叶绿素a,b含量测定 [实验目的]熟悉在未经分离的叶绿素溶液中测定叶绿素a和b的方法及其计算。 [实验原理]在叶绿素a和b的吸收光谱曲线中,红波波长范围内,叶绿素a的最大吸收峰在663nm,叶绿素b的最大吸收峰在645nm。吸收曲线彼此又有重叠。 根据Lambert—Beer定律,最大吸收峰不同的两个组分的混合液,它们的浓度C与光密度OD之间有如下关系:OD1=Ca·ka1+Cb·kb1 (1) OD2=Ca·ka2+Cb·kb2 (2) Ca为组分a的浓度(g/L) Cb为组分b的浓度(g/L) OD1为在波长λ1(即组分a的最大吸收峰波长)时,混合液的光密度OD值。 OD2为在波长λ2(即组分b的最大吸收缝波长)时,混合液的光密度OD值。 ka1,kb1,ka2,kb2分别为组分a,b的比吸收系数,即组分a(b)的浓度为(1g/L)时,其在相应波长(λ1,λ2)时的光密度OD值。 叶绿素A和B的80%丙酮溶液,当浓度为1时,比吸收系数K值如下表: 将表中数值代入上式(1),(2)并整理的: Ca=0.0127OD663-0.00269OD645 Cb=0.0229OD645-0.00468OD663 若把Ca,Cb的浓度单位从原来的g/L改为mg/L,则上式可改写为下列形式: Ca=12.7OD663-2.69OD645 (3) Cb=22.9OD645-4.68OD663 (4) Ct=Ca+Cb=8.02OD663+20.21OD645 (5) Ct为叶绿素总浓度,单位为g/L。 利用(3),(4),(5)式即可计算出叶绿素A和B及总叶绿素的浓度(g/L)。 [器材与试剂] 1.实验仪器:高级型分光光度计,离心机,台天平,剪刀,研钵,漏斗,移液管 2.实验试剂:丙酮,碳酸钙 3.实验材料:植物叶片 [实验步骤] 1.色素的提取:取新鲜叶片,剪去粗大的叶脉并剪成碎块,称取0.5G放入研钵中加纯丙酮5ML,少许碳酸钙和石英砂,研磨成匀浆,再加80%丙酮5ML,将匀浆转入离心管,并用适量80%丙酮洗涤研钵,一并转入离心管,离心后弃沉淀,上清液用80%丙酮定容至20ML。 2.测定光密度:取上述色素提取液1ml,加80%丙酮4ml稀释和转入比色杯中,以80%丙酮为对照,分别测定663nm,645nm处的光密度值。 3.按公式分别计算色素提取液中叶绿素A,B及叶绿素总浓度。再根据稀释倍数分别计算每克鲜重叶片中色素的含量。 [注意事项] 1.由于植物子叶中含有水分,故先用纯丙酮进行提取,以色素提取液中丙酮的最终浓度近似80%。 2.由于叶绿素A,B的吸收峰很陡,仪器波长稍有偏差,就会使结果产生很大的误差,因此最好能用波长较正确的高级型分光光度计。 [实验作业] 1.试比较阴生植物和阳生植物的叶绿素A和叶绿素B的比值有无不同。 2.分光光度法和比色法有何不同? 3.叶绿素A和叶绿素B在红光区和蓝光区都有最大吸收峰,能否用蓝光区的最大吸收峰波长进行叶绿素A和叶绿素B 的定量分析,为什么? 叶绿素含量测定方法---丙酮法 由于微藻的生长周期比较复杂,包括无性繁殖阶段和有性繁殖阶段,其在不同阶段的生理形态不同,有时藻细胞会聚集在一起,以片状或团状形式存在,在显微镜下难以确定其所包含的细胞数量。 藻细胞中叶绿素的含量(特别是叶绿素a的含量)通常随与细胞的生长呈较好的线性关系,因此可通过测定藻细胞中叶绿素含量变化来反映微藻的生长情况。叶绿素测定采用丙酮研磨提取法。 取适量藻液于10 mL离心管中在4000 rpm转速下离心10 min,弃去上清液,藻泥中加入适量的100 %的丙酮。采用丙酮提取法时在试管研磨器中冰浴研磨5 min,4000 rpm离心后,上清液转入10 mL容量瓶中。按上述方法对藻体沉淀进行萃取,直至藻体沉淀呈白色为止。定容后,采用722S型可见分光光度计分别测定645 nm和663 nm下萃取液的吸光值,叶绿素含量用以下公式进行计算(Amon,1949): 叶绿素a含量用以下公式进行计算: Chlorophyll a (mg/L) = (12.7×A663 nm-2.69×A645 nm)×稀释倍数 叶绿素b含量用以下公式进行计算: Chlorophyll b (mg/L) = (22.9×A645 nm-4.64×A663 nm)×稀释倍数 叶绿素总含量用以下公式进行计算: Chlorophyll a+b (mg/L) = (20.2×A645 nm+8.02×A663 nm)×稀释倍数 由于丙酮的沸点较低,较高温度下挥发很快。此外,叶绿素稳定性较差,见光易分解,因此,本实验中叶绿素的提取和测定均在低温黑暗条件下进行,以减少提取过程中的损失。 叶绿素提取方法 提取液:本试验用DMSO/80%丙酮(l/2,v/v)提取的叶绿素,谭桂英周百成底栖绿藻叶绿素的二甲基亚砜提取和测定法* 海洋与湖沼 1987 18(3)295--300. 一、直接浸提法: 1、准确量取10ml藻液,加到15ml离心管中,放在台式离心机离心,3500r/min (根据不同的藻选择不同那个的离心转速)离心5min倒上清;留藻泥。随后在盛有藻泥的离心管中加入蒸馏水,与藻泥混匀后再次离心,目的是除去藻细胞表面的盐份,此清洗过程重复三次。 2、往藻泥中加二甲基亚砜3.33ml,65℃水浴9h,20h; 3、然后离心,将上清转移到10ml棕色瓶中, 4、添加6.67ml80%丙酮到离心管中,混匀,离心,再将上清转移到10ml棕色瓶中。 5、定容,待测。 测定叶绿素a和b的方 法及其计算 Document serial number【NL89WT-NY98YT-NC8CB-NNUUT-NUT108】 实验二十五测定叶绿素a和b的方法及其计算 一目的要求: 熟悉在未经分离的叶绿体色素溶液中测定叶绿素a和b的方法及其计算。 二实验原理: 如果混合液中的两个组分,它们的光谱吸收峰虽然有明显的差异,但吸收曲线彼此有些重叠,在这种情况下要分别测定两个组分,可根据Lambert-Beer定律,通过代数方法,计算一种组分由于另一种组分存在时对光密度的影响,最后分别得到两种组分的含量。 如图z-4叶绿素a和b的吸收光谱曲线,叶绿素a的最大吸收峰在663nm,叶绿素b在645nm,吸收曲线彼此又有重叠。 图z-4 叶绿素a和b的吸收光谱曲线 横坐标为波长(nm),纵坐标为比吸收系数 根据Lambert-Beer定律,最大吸收光谱峰不同的两个组分的混合液,它们的浓度C与光密度OD之间有如下的关系: OD1=Ca·ka1+Cb·kb1 (1) OD2=Ca·ka2+Cb·kb2 (2) 式中:Ca为组分a的浓度,g/L。 Cb为组分b的浓度,g/L。 OD1为在波长λ1(即组分a的最大吸收峰波长)时,混合液的光密度OD值。 OD2为在波长λ2(即组分b的最大吸收峰波长)时,混合液的光密度OD值。 ka1为组分a的比吸收系数,即组分a当浓度为1g/L时,于波长λ1时的光密度OD值。 kb2为组分b的比吸收系数,即组分b当浓度为1g/L时,于波长λ2时的光密度OD值。 ka2为组分a(浓度为1g/L),于波长λ2时的光密度OD值。 kb1为组分b(浓度为1g/L),于波长λ1时的光密度OD值。 从文献中可以查到叶绿素a和b的80%丙酮溶液,当浓度为1g/L时,比吸收系数k值如下: 将表中数值代入上式(1)、(2),则得: OD663=×Ca+×Cb OD645=×Ca+×Cb 经过整理之后,即得到下式: Ca= OD645 Cb= OD663 如果把Ca,Cb的浓度单位从原来的g/L改为mg/L,则上式可改写为下列形式: Ca= OD645 (3) Cb= OD663 (4) CT= Ca+ Cb= OD663+ OD645 (5) (5)式中CT为总叶绿素浓度,单位为mg/L。 利用上面(3)、(4)、(5)式,即可计算出叶绿素a和b及总叶绿素的浓度 (mg/L)。 [附注]一般大学教学实验室所用的分光度计多为721型,属低级类型,其单色光的半波宽要比中级类型的751型宽得多,而叶绿素a和b吸收峰的波长相差仅18nm(663-645nm),难以达到精确测定。此外有时还由于仪器本身的标称波长与实际波长不符,叶绿素含量的测定
植物对盐胁迫的反应
植物生理学实验-叶绿素a b测定
叶绿素含量测定方法(精)
测定叶绿素a和b的方法及其计算完整版
相关主题
文本预览