
Odor Coding in a Model Olfactory Organ:The Drosophila Maxillary Palp
Marien de Bruyne,Peter J.Clyne,and John R.Carlson
Department of Molecular,Cellular,and Developmental Biology,Yale University,New Haven,Connecticut06520-8103
Odor coding relies on the activity of different classes of receptor neurons,each with distinct response characteristics.We have examined odor coding in a model olfactory organ,the maxillary palp of Drosophila.This organ contains only120olfactory receptor neurons,compartmentalized in sensory hairs called sensilla,and provides an opportunity to characterize all neurons in an entire olfactory organ.Extensive extracellular recordings from single sensilla reveal that the neurons fall into six func-tional classes.Each of the60sensilla houses two neurons, which observe a pairing rule:each sensillum combines neurons of two particular classes,thereby yielding three sensillum types. The sensillum types are intermingled on the surface of the palp, but their distribution is not random.The neurons exhibit diverse response characteristics,providing the basis for an olfactory code.A particular odor can excite one neuron and inhibit another,and a particular neuron can be excited by one odor and inhibited by another.Some excitatory responses continue beyond the end of odor delivery,but responses to most odors terminate abruptly after the end of odor delivery,with some followed by a period of poststimulus quiescence.The speci?c-ity of odor response is examined in detail for the neurons of one sensillum,which were found to differ in their relative responses to a homologous series of esters.Adaptation and cross-adaptation are documented,and cross-adaptation experiments demonstrate that the two neurons within one type of sensillum can function independently.The analysis of all neuronal types in this model olfactory organ is discussed in terms of its functional organization and the mechanisms by which it encodes olfactory information.
Key words:Drosophila;olfaction;maxillary palps;odor cod-ing;single-unit electrophysiology;sensory?eld;adaptation; inhibition
Olfactory systems detect and differentiate among many kinds of odor stimuli.Olfactory receptor neurons encode qualitative, quantitative,temporal,and spatial information about odors.Elu-cidating the mechanisms by which olfactory information is en-coded is an intriguing problem in contemporary neurobiology (Derby and Ache,1984;Buck,1996;Hildebrand and Shepherd, 1997).A major dif?culty has been the complexity of most olfac-tory systems.In the case of mammals,for example,the number of olfactory receptor neurons(ORNs)is very large,as is the number of distinct functional classes into which these neurons may fall (Buck and Axel,1991).Moreover,in many systems it is dif?cult to measure systematically the physiological properties of individual ORNs in vivo(Sicard and Holley,1984).
Insect ORNs are distributed in sensilla,usually in the form of sensory hairs that protrude from the cuticle,providing accessibil-ity and ease of identi?cation(Altner and Prillinger,1980;Boeckh, 1981).Odorants pass through tiny pores in the walls of these sensilla and stimulate dendrites bathing in the lymph inside.Most sensilla are on the antenna,and elegant analyses have character-ized their physiological properties in moths,cockroaches,and other insects with large antennae(Boeckh et al.,1987;Hansson, 1995).
The?y Drosophila melanogaster has a relatively simple olfac-tory system.Olfactory response can be measured in vivo,via either physiological or behavioral means,and a variety of genetic and molecular approaches are available to study its olfactory system(Siddiqi,1991;Carlson,1996).There are only1200ORNs in the Drosophila antenna,housed in three morphologically dis-tinct sensillum categories:trichoid,coeloconic,and basiconic sen-silla(Stocker,1994)(Fig.1A).Electrophysiological recordings from each of these categories indicate that they can be subdivided into different functional types(Siddiqi,1991;Clyne et al.,1997), but an exhaustive description is not available.
Even simpler than the antenna is the maxillary palp,an olfac-tory organ that extends from the proboscis and that contains an order of magnitude fewer neurons than does the antenna(Fig. 1B,C)(Singh and Nayak,1985).Each of the paired maxillary palps contains only120ORNs,housed in60sensilla of a single category,sensilla basiconica.The sensitivity of the Drosophila maxillary palp to a variety of odors has been demonstrated by ?eld recordings(Ayer and Carlson,1992).The numerical sim-plicity of the maxillary palp makes it possible to perform an exhaustive study of its neuronal composition and to characterize in detail its functional organization.
In this article we provide an extensive description of the olfac-tory receptor neurons of the maxillary palp,a simple model olfactory https://www.doczj.com/doc/484304006.html,ing single-unit recordings(Kaissling,1995) and a panel of test odorants,we characterize the response pro?les of the neurons and?nd that they fall into distinct functional classes,with different classes presumably re?ecting distinct recep-tor–ligand interactions.We consider the issue of how narrowly some of the neurons are tuned.The neurons are shown to be housed in stereotyped pairs in three functional types of sensilla,
Received Jan.13,1999;revised March12,1999;accepted March18,1999.
This work was supported by the Human Frontier Science Program Fellowship LT
277/97to M.d.B.,by a National Science Foundation predoctoral fellowship to P.J.C.,
and by National Institutes of Health Grant DC-02174and a grant from the Human
Frontier Science Program to J.R.C.
Correspondence should be addressed to Dr.John R.Carlson,Department of
Molecular,Cellular,and Developmental Biology,Yale University,Kline Biology
Tower,P.O.Box208103,New Haven,CT06520-8103.
Copyright?1999Society for Neuroscience0270-6474/99/194520-13$05.00/0
The Journal of Neuroscience,June1,1999,19(11):4520–4532
whose distributions are characterized.Physiological analysis of the neurons reveals a diversity of response mechanisms.We also document adaptation and cross-adaptation of these neurons,and via cross-adaptation experiments,we ?nd evidence that the two neurons that cohabit in a sensillum can function independently.
MATERIALS AND METHODS
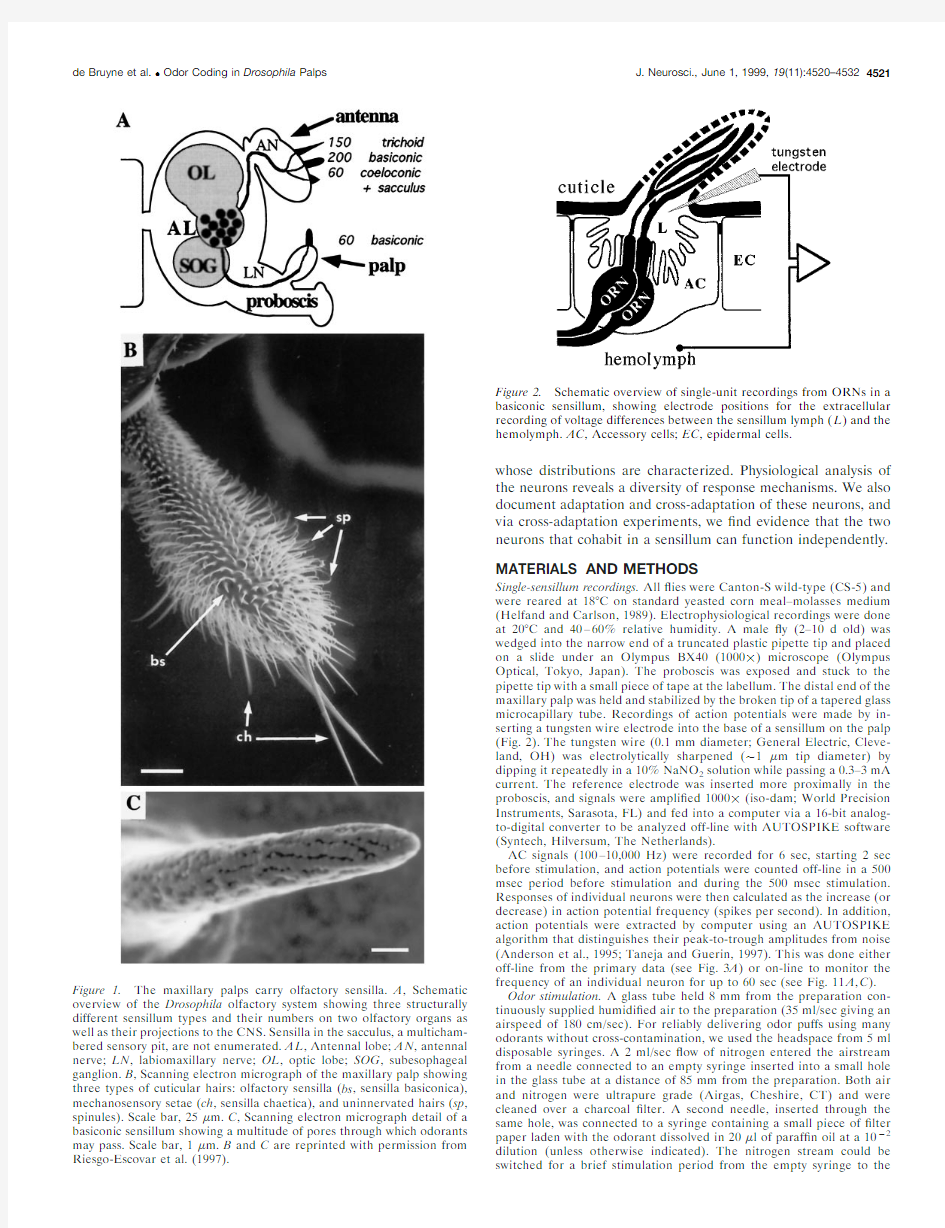
Single-sensillum recordings.All ?ies were Canton-S wild-type (CS-5)and were reared at 18°C on standard yeasted corn meal–molasses medium (Helfand and Carlson,1989).Electrophysiological recordings were done at 20°C and 40–60%relative humidity.A male ?y (2–10d old)was wedged into the narrow end of a truncated plastic pipette tip and placed on a slide under an Olympus BX40(1000?)microscope (Olympus Optical,Tokyo,Japan).The proboscis was exposed and stuck to the pipette tip with a small piece of tape at the labellum.The distal end of the maxillary palp was held and stabilized by the broken tip of a tapered glass microcapillary tube.Recordings of action potentials were made by in-serting a tungsten wire electrode into the base of a sensillum on the palp (Fig.2).The tungsten wire (0.1mm diameter;General Electric,Cleve-land,OH)was electrolytically sharpened (?1?m tip diameter)by dipping it repeatedly in a 10%NaNO 2solution while passing a 0.3–3mA current.The reference electrode was inserted more proximally in the proboscis,and signals were ampli?ed 1000?(iso-dam;World Precision Instruments,Sarasota,FL)and fed into a computer via a 16-bit analog-to-digital converter to be analyzed off-line with AUTOSPIKE software (Syntech,Hilversum,The Netherlands).
AC signals (100–10,000Hz)were recorded for 6sec,starting 2sec before stimulation,and action potentials were counted off-line in a 500msec period before stimulation and during the 500msec stimulation.Responses of individual neurons were then calculated as the increase (or decrease)in action potential frequency (spikes per second).In addition,action potentials were extracted by computer using an AUTOSPIKE algorithm that distinguishes their peak-to-trough amplitudes from noise (Anderson et al.,1995;Taneja and Guerin,1997).This was done either off-line from the primary data (see Fig.3A )or on-line to monitor the frequency of an individual neuron for up to 60sec (see Fig.11A ,C ).Odor stimulation.A glass tube held 8mm from the preparation con-tinuously supplied humidi?ed air to the preparation (35ml/sec giving an airspeed of 180cm/sec).For reliably delivering odor puffs using many odorants without cross-contamination,we used the headspace from 5ml disposable syringes.A 2ml/sec ?ow of nitrogen entered the airstream from a needle connected to an empty syringe inserted into a small hole in the glass tube at a distance of 85mm from the preparation.Both air and nitrogen were ultrapure grade (Airgas,Cheshire,CT)and were cleaned over a charcoal ?lter.A second needle,inserted through the same hole,was connected to a syringe containing a small piece of ?lter paper laden with the odorant dissolved in 20?l of paraf?n oil at a 10?2dilution (unless otherwise indicated).The nitrogen stream could be switched for a brief stimulation period from the empty syringe to
the
Figure 1.The maxillary palps carry olfactory sensilla.A ,Schematic overview of the Drosophila olfactory system showing three structurally different sensillum types and their numbers on two olfactory organs as well as their projections to the CNS.Sensilla in the sacculus,a multicham-bered sensory pit,are not enumerated.AL ,Antennal lobe;AN ,antennal nerve;LN ,labiomaxillary nerve;OL ,optic lobe;SOG ,subesophageal ganglion.B ,Scanning electron micrograph of the maxillary palp showing three types of cuticular hairs:olfactory sensilla (bs ,sensilla basiconica),mechanosensory setae (ch ,sensilla chaetica),and uninnervated hairs (sp ,spinules).Scale bar,25?m.C ,Scanning electron micrograph detail of a basiconic sensillum showing a multitude of pores through which odorants may pass.Scale bar,1?m.B and C are reprinted with permission from Riesgo-Escovar et al.
(1997).
Figure 2.Schematic overview of single-unit recordings from ORNs in a basiconic sensillum,showing electrode positions for the extracellular recording of voltage differences between the sensillum lymph (L )and the hemolymph.AC ,Accessory cells;EC ,epidermal cells.
de Bruyne et al.?Odor Coding in Drosophila Palps J.Neurosci.,June 1,1999,19(11):4520–45324521
odor-?lled syringe by a solenoid valve expelling part of the headspace into the airstream.Thus,all elements of the airstream were kept constant except for a brief pulse of odor.A500msec stimulus period was used in all recordings using this“syringe puff”method.
A series of odors was administered three times,with the odors pre-sented one after the other and an interval of at least60sec between the delivery of each odor.For structure–response studies with a series of nine esters,the order of presentation was randomized.For dose–response relations,odorants were presented with increasing doses in log or half-log steps.All odors were from Aldrich(Milwaukee,WI)and of the highest grade available(97–99%)except cis-vaccenyl acetate(Z11-octadecenyl acetate),which was from Sigma(99%;St.Louis,MO).For ?-pinene,we used the R(?)isomer,and4-methylcyclohexanol was a mix of cis and trans isomers.The paraf?n oil diluent was IR-spectroscopy grade(Fluka,Buchs,Switzerland)
Cluster analysis.An objective classi?cation of data sets into classes of neurons was made with a hierarchical cluster analysis(Anderberg,1973; Derby and Ache,1984;Bieber and Smith,1986)using JMP software (SAS Institute,Cary,NC).This method organizes a data set into discrete clusters on the basis of a number of variables describing each data point. In the case of ORNs,the variables we based the classi?cation on are the responses to a set of?ve odorants(see Fig.7).With the responses to each odorant as parameters,each neuron’s pro?le is a point in a?ve-dimensional space.An iterative process then groups the points into clusters on the basis of their distances,starting with the closest points, until all are grouped.The distances were calculated according to Ward’s method,which minimizes variance(i.e.,ANOVA sum of squares)within clusters(Ward,1963;Getz and Akers,1997).The number of clusters that best represents the organization of the data is determined with the scree procedure(Bieber and Smith,1986).This procedure identi?es a sharp increase in the distances among clusters,indicating a change in the nature of the clustering steps.
Adaptation experiments.For adapting ORNs to continuing stimulation with one particular odorant,we needed to ensure that stimulus intensity stayed constant during a long stimulation period.The syringe puff method only allows reliable delivery of small samples from5ml of odor-saturated air.Instead we established a dynamic equilibrium be-tween ethyl propionate,diluted10?2in200?l of paraf?n oil applied to a?lter paper on the inside wall of a15ml gas-wash?ask,and a continuous air?ow(2ml/sec).A solenoid valve then sent this odor-laden air via a needle either into the airstream toward the preparation or into a vacuum line to prevent odor buildup in the room.The equilibrium in this“?ask?ow”method was established during a period of at least3min before stimulation,and a single?ask was not used for?20min.The response of the two cells in one sensillum was recorded during a500msec stimulation with the syringe puff method.The sensillum was then stim-ulated for25sec with the?ask?ow method,and1sec after this,the response to a second500msec odor puff identical to the?rst was again recorded.
RESULTS
Olfactory receptor neurons of the palp show excitatory and inhibitory responses
ORNs from single sensilla on the Drosophila maxillary palp exhibited spontaneous action potentials,clearly distinguishable from background noise.The spikes from an individual sensillum could be resolved into two distinct populations,based on their amplitudes.A recording from a typical sensillum is shown in Figure3A,with the spikes of each population,large and small, indicated separately below the primary data.This bimodal distri-bution of spike amplitudes is illustrated in Figure3B.We inter-pret the two populations of spikes as representing the activities of two distinct neurons,an interpretation that has been extensively supported in a wide variety of other insect sensory systems (Lof-
Figure3.Single-unit recordings from palpal basiconic sensilla con?rm the presence
of two olfactory receptor neurons.A,Differences in spike amplitudes allow separate
analysis of?ring rates of the two neurons in a single sensillum.Top,Primary data
from a3sec period of spontaneous activity from a sensillum are shown.Middle,
bottom,The extracted data are presented with large spikes shown separately from
small spikes.B,The distribution of amplitudes of individual action potentials
(measured from peak-to-trough)is bimodal.Data shown are for123spikes from6
sec of spontaneous activity of the recording in A.
4522J.Neurosci.,June1,1999,19(11):4520–4532de Bruyne et al.?Odor Coding in Drosophila Palps
tus and Corbie `re-Tichane ′,1981;Fujishiro et al.,1984;Hansson et al.,1994;Anderson et al.,1995;Getz and Akers,1997;Taneja and Guerin,1997).In the case of the Drosophila maxillary palp,the presence of two neurons has been independently demonstrated in an ultrastructural analysis by Singh and Nayak (1985),who found two sensory cells with branched dendrites in individual basiconic sensilla.We will refer henceforth to the two neurons of a sensil-lum as the A and B cells.
Diversity among the olfactory neurons of the maxillary palp can be observed by analysis of their spontaneous ?ring rates.The spontaneous action potential frequency of individual cells varies,with most neurons exhibiting frequencies between 3and 13spikes/sec.The spontaneous rate of one of these individual cells is relatively constant,however;it stayed within a range of ?3spikes/sec over the course of recording periods lasting as long as 60min.In addition to these neurons,some neurons had notice-ably higher spontaneous rates of ?30spikes/sec (see Fig.6).Odor stimulation elicited a marked increase in ?ring frequency in many cases (Fig.4).In most cases one of the two neurons in a sensillum was clearly more excited than was the other by stimu-lation with a particular odor.The excited neuron can generally be identi?ed by its spike height at the start of the response.For example,in Figure 4A only the A cell,that with greater ampli-tude,shows an increased frequency of ?ring in response to odor stimulation.The B cell,whose smaller spikes are indicated by dots ,shows no excitation.In some cases the spike amplitude of the responding neuron gradually changes during the course of stim-ulation,a phenomenon widely observed in single-sensillum re-cordings from insects (discussed in Guillet and Bernard,1972;Fujishiro et al.,1984;Rumbo,1989).In such cases,the
identities
Figure 4.Sensilla house receptor neurons with a variety of different response characteristics to different odors.A–E ,Five 1500msec traces of recordings from two different sensilla showing excitatory and inhibitory responses of the two cells to 500msec stimulations (horizontal lines )with different odorants.For odor stimulation,air was expelled from 5ml syringes over ?lter paper laden with 20?l of odorant.The odorants were diluted 10?2in paraf?n oil.We do not know the exact concentrations of the odor present in the air reaching the preparation.A ,B ,Recordings from one sensillum,later classi?ed as https://www.doczj.com/doc/484304006.html,rge action potentials,from the A neuron,increase their frequency in response to ethyl acetate.Dots indicate smaller action potentials from the B neuron,which is not excited by ethyl acetate but which responds to 4-methylphenol.C–E ,Recordings from another sensillum,later classi?ed as https://www.doczj.com/doc/484304006.html,rge spikes are from the B neuron,which is excited by 4-methylphenol and inhibited by other odors.Smaller spikes are from the A neuron,excited by benzaldehyde.
de Bruyne et al.?Odor Coding in Drosophila Palps J.Neurosci.,June 1,1999,19(11):4520–45324523
of the neurons can often be con?rmed if necessary by examination of the spike shapes,which differ between the two neurons.Exci-tatory responses started?80msec after odor was administered; calculations reveal that50msec is required for odor to reach the preparation,indicating a response latency of?30msec.The spike frequency rose to a maximum within50–100msec after response initiation,depending on the odor and dose,and then declined as described in detail below.
Excitatory responses terminated abruptly after the end of the odor stimulation period in most cases,such as that shown in Figure4A.This is consistent with the expected sharp decline in odor levels after the500msec delivery period.In these cases we often observed a brief period in which no action potentials were recorded from this neuron.The duration of this poststimulus quiescence appeared to be dose-dependent,with higher doses producing longer periods of quiescence(data not shown),al-though we have not examined this relationship quantitatively. Poststimulus quiescence was not,however,observed for all odors. In fact,in some cases excitation continues long past the end of odor stimulation;Figure4B shows a recording from the same sensillum analyzed in Figure4A but stimulated with 4-methylphenol instead of ethyl acetate.In this case stimulation continues past the end of the odor delivery period and declines to spontaneous levels over the course of several seconds.The same stimulus elicits a similar response,of smaller magnitude,from a different sensillum and again shows a prolonged pattern of?ring (Fig.4C).Thus the variations in response at the end of odor stimulation show that different odors may have different effects on the kinetics of excitation,thereby providing one putative mecha-nism for odor discrimination.
Inhibitory responses were also observed in some sensilla.Fig-ure4D shows inhibition of?ring in one neuron of such a sensil-lum after stimulation with3-octanol.Moreover,this neuron can be excited by one odor and inhibited by others.For example,the neuron shown to be inhibited by3-octanol in Figure4D is the same one excited by4-methylphenol in Figure4C.Inhibition was observed only in those cells that showed the higher rate of spontaneous action potentials(30spikes/sec).Interestingly,in these sensilla some odors inhibit one cell but excite the other(Fig. 4E);the cell with the large spikes is inhibited,and the cell with the small spikes is excited.Thus an individual odor can have
opposite effects on the?ring frequency of different neurons,and an individual neuron can respond oppositely to different odors.
The olfactory neurons fall into six functional classes and are housed in stereotyped pairs within three sensillum types
To de?ne the basic elements of the olfactory code,we sought to determine whether ORNs fall into discrete functional classes and, if so,to determine the number and odor speci?city of such classes. We initiated this analysis with a chemically diverse group of16 odorants selected on the basis of three criteria.Some odorants (ethyl acetate,3-octanol,and benzaldehyde)were selected be-cause they had been used extensively in previous research on Drosophila olfaction and are known to induce strong behavioral responses.Others were selected because they play important roles in the ecology of related dipteran insects.For example, 4-methylphenol is present in cattle urine and attracts tsetse?ies to their hosts(Bursell et al.,1988);E2-hexenal(leaf aldehyde)is a plant odor that attracts many insect species(Visser,1986). Finally,odors were selected to represent certain chemical groups (e.g.,ketones,aldehydes,alcohols,esters,and aromatics).
Our initial recordings from individual sensilla clearly indicated that sensilla could be divided into three functional types,which we have termed palpal basiconic1(pb1),pb2,and pb3.Although of the same morphological category,the three types of sensilla contain neurons with response spectra that are distinct from each other and from neurons of the other sensillum types.Figure5 shows responses to the16test odors recorded from the two neurons of the pb1sensillum,neurons that we denote pb1A and pb1B.The pb1B cell responds strongly to only one of the tested odorants,4-methylphenol,to which it shows an increase in spike frequency of178?47spikes/sec(?SD;n?13).The only other response from pb1B that was signi?cantly different from that of the paraf?n oil control is the response to4-methylcyclohexanol, an odor molecule structurally similar to4-methylphenol.The pb1A cell,by contrast,shows a broader range of responses to the tested stimuli;it responds most strongly to ethyl acetate,showing an increase of138?32spikes/sec(?SD;n?13),but also responds to several other stimuli.
The different neurons of the maxillary palp could be distin-guished using a diagnostic subset of7of the16odors.
We Figure5.Responses of the two pb1neurons to a set of16odorants(error bars indicate SD;n?13).pb1B responds strongly only to one of the tested odors.Responses of pb1A to several odorants are signi?cantly larger than the control response to the paraf?n oil diluent alone.The indicated ORN response is measured as the increase in spikes per second over the spontaneous frequency.
4524J.Neurosci.,June1,1999,19(11):4520–4532de Bruyne et al.?Odor Coding in Drosophila Palps
therefore used these 7odors to extend our analysis to a larger number of sensilla.The results con?rm the presence of exactly three sensillum types,each containing two neurons,yielding a total of six types of neurons with distinguishable response prop-erties:pb1A,pb1B,pb2A,pb2B,pb3A,and pb3B (Fig.6).The
pb1sensilla contain an A cell that responds strongly to ethyl acetate and a B cell that responds strongly to 4-methylphenol.In the pb2sensilla,the A cell responds strongly to benzaldehyde.The other cell,pb2B,is excited by 4-methylphenol,although not as strongly as is pb1B.In addition,pb2B is strongly inhibited by 3-octanol and several other odors.Speci?cally,the spontaneous ?ring frequency of pb2B is 32?7spikes/sec and is reduced 80–100%by 3-octanol.The pb3sensillum contains two neurons that are both excited by 3-octanol and isoamyl acetate,but pb3B is more strongly stimulated by isoamyl acetate than is pb3A.In summary,this analysis revealed that the maxillary palp contains six distinguishable neuronal types.The responses of some neu-ronal types are overlapping (e.g.,4-methylphenol excites both pb1B and pb2B neurons),but comparisons of responses to mul-tiple odors clearly showed the pro?les to be distinct.We note ?nally that in pb1and pb3sensilla,A neurons consistently have larger spike amplitudes than B neurons.pb2sensilla differ from pb1and pb3sensilla not only in that the B neuron consistently has higher spontaneous activity than the other ?ve neuronal types but also in that pb2A has smaller spikes than pb2B in some sensilla.However,spikes of pb2A are consistently different in shape,having larger negative phases relative to the positive phase.The two neurons in one sensillum observe a pairing rule;for example,the cell that is excited by benzaldehyde (pb2A)is always paired with a cell that is inhibited by 3-octanol (pb2B).Among 232sensilla examined,we recorded the activity of two cells in 225cases (in the 7exceptional sensilla,one cell was absent or unre-sponsive,perhaps because of damage caused by the recording electrode in at least some cases);in 222of the 225cases,we found one of the three characteristic combinations of cells.
Coding of odor quality and quantity across cell classes
Although the existence of exactly six neuronal classes seemed clear from this physiological analysis (Fig.6),we sought to test our classi?cation scheme more rigorously.We therefore per-formed a cluster analysis of the responses of 54neurons in 27sensilla to ?ve of the odors (Fig.7).This analysis yielded ?ve clusters based on the response pro?les.Cluster 1groups cells with a strong response to ethyl acetate (corresponding to pb1A),and cluster 2unites cells responding strongly to benzaldehyde (pb2A).Clusters 5and 4contain cells that respond to 4-methylphenol but that either are or are not inhibited by 3-octanol (pb2B and pb1B,respectively).Cluster 3includes cells that are excited by both isoamyl acetate and 3-octanol but that show no strong response to the other odors;this cluster includes both pb3A and pb3B cells.This analysis con?rms our initial observation that ORNs on the palp can reliably be assigned to cell classes that have clearly distinct response characteristics.
This cluster analysis,however,did not resolve pb3A and pb3B.Therefore we sought further evidence that pb3A and pb3B are distinguishable by testing their responses to an additional set of odors.Signi?cant responses from pb3cells were only observed to 3of the initial 16odorants (data not shown).We chose a homol-ogous series of esters because our previous analysis had revealed a difference in the responses of pb3A and pb3B to isoamyl acetate (Fig.6).We tested aliphatic esters (Fig.8A )with varying chain lengths of both the alcohol and acid moieties.Figure 8,B and C ,shows that the responses of pb3A and pb3B are clearly distin-guishable,in two respects.First,for most of the esters there is a signi?cant difference in the absolute responses of the two neu-rons;for example,for pentyl acetate (5:2),the response of pb3B is 99?12spikes/sec,whereas the response of pb3A is
only
Figure 6.Six classes of olfactory receptor neurons are found in charac-teristic pairs (A and B neurons)in three functional types of sensilla on the Drosophila maxillary palp.Response pro?les of ORNs are shown for the three sensillum types:pb1(n ?17),pb2(n ?15),and pb3(n ?14).The spontaneous frequencies of the two neurons (?SD)are indicated in the top right corner for each type.The ORN response is measured as the increase (or decrease)in spikes per second over the spontaneous frequency.Error bars indicate SEM and are too small to be seen in some cases.
de Bruyne et al.?Odor Coding in Drosophila Palps J.Neurosci.,June 1,1999,19(11):4520–45324525
9?2spikes/sec (note the different scales of the y -axes in Fig.8B,C ).Second,the relative responses of the two neurons to different odors vary;for example,the pb3A neuron responds dramatically better to ethyl butyrate (2:4)than to hexyl acetate (6:2),whereas for pb3B the converse is true.This analysis con-?rms that the response spectra of pb3A and pb3B are clearly different and that the pb3sensillum,like pb1and pb2,contains two distinguishable neurons.We note with interest that for
pb3A,
Figure 7.Dendrogram of a hierarchical cluster analysis of 54ORNs in 27sensilla,comparing their responses to ?ve odors.Branch length is proportional to distance (see Materials and Methods),and ?ve clusters are indicated by the numbered points .Individual ORNs are indicated as the A or B cell of a single sensillum,and their responses,measured as the increase in spikes per second over the spontaneous frequency (y -axis),are shown.Six classes of neurons are indicated at the bottom of the ?gure.Clusters 1,2,4,and 5correspond to pb1A,pb2A,pb1B,and pb2B,respectively;cluster 3includes two cell types within the pb3sensillum,neurons pb3A and pb3B,that can be clearly distinguished by their response to other odors (see Fig.8)and by differences in spike amplitude (data not shown).
4526J.Neurosci.,June 1,1999,19(11):4520–4532de Bruyne et al.?Odor Coding in Drosophila Palps
the length of the odorant molecule correlates with potency;odors that elicit the strongest mean responses are those with a total of 6carbons,with the next most potent being those with 4,5,or 7carbons,followed by those with 8or 10carbons.However,none of the tested odors elicits more than ?30spikes/sec from this neu-ron,leading us to suspect strongly that there are other molecules in odor space that are more effective stimuli for this neuron.The six neuron classes differ in their response patterns;that is,odor quality may be encoded by spike-frequency differences across cell classes.However,ORNs also need to be able to encode different levels of stimuli (i.e.,odor quantity).Are differences in response magnitudes constant across a range of doses of the different odors?We tested pb1A with the homologous series of esters and found that the greatest mean response was provoked by ethyl propionate (2:3)(Fig.8D ).We extended this observation by generating dose–response curves (Fig.9).The response to ethyl propionate appears as a sigmoid curve,spanning several orders of magnitude and reaching saturation at ?225spikes/sec.Dose–response curves for pb1A are also shown for three other odors.The differences in response magnitudes for the different esters vary widely as a function of odor concentration.When tested with 10?4dilutions,pb1A appears narrowly tuned for ethyl propi-onate;at higher concentrations of odors,the differences are less striking.
The three sensillum types have a mixed,but not random,distribution across the surface of the palp
Is there a form of topographical encoding of odor quality on the maxillary palp?The distinct response characteristics allow rapid identi?cation of the three sensillum types.We recorded from 131sensilla at various positions on the dorsal and lateral surfaces of one of the two palps of 43?ies,sampling almost the entire population of olfactory sensilla.There are no basiconic sensilla on the ventral and extreme proximal surfaces (Singh and
Nayak,
Figure 8.Responses of olfactory receptor neurons on the Drosophila maxillary palp to aliphatic esters con?rm and characterize the distinct identities of pb3A and pb3B.A ,Nomenclature illustrated with the struc-ture of butyl acetate.i 5:2is isoamyl acetate,a branched analog of pentyl acetate (5:2).B –D ,Responses of pb3A,pb3B,and pb1A,respectively,to a set of esters with varying chain lengths and structures.The indicated ORN response is measured as the increase in spikes per second over the spontaneous frequency (error bars indicate SEM;n ?8).n.d.,Not
done.
Figure 9.Dose–response relations for pb1A to four odorants.Stimuli were presented as dilutions in 20?l of paraf?n oil on ?lter paper as described in Figure 4.The indicated ORN response is measured as the increase in spikes per second over the spontaneous frequency (error bars indicate SEM;n ?13).The dotted horizontal line indicates the mean response of pb1A to the paraf?n oil diluent alone.
de Bruyne et al.?Odor Coding in Drosophila Palps J.Neurosci.,June 1,1999,19(11):4520–45324527
1985).The sensillum type was easily determined using a few odors;4-methylphenol distinguished pb1and pb2from pb3,and then pb1sensilla were identi?ed by their response to ethyl ace-tate,whereas pb2sensilla were inhibited by 3-octanol.Their distribution over the surface is shown in Figure 10.Their topog-raphy reveals partially overlapping distributions.Over most of the surface the three types are intermingled;thus they are not in mutually exclusive zones.However,pb2sensilla are virtually excluded from a proximolateral region (Fig.10,arrow ),indicating that the distribution of sensillum types is not random.
In this analysis the numbers of sensilla of each type were approximately equal:38,30,and 32%for pb1,pb2,and pb3,respectively (n ?131sensilla).However,to con?rm these results,we performed a second,independent experiment in which we conceptually divided the olfactory surface of the palp into ?ve arbitrary regions of comparable area and recorded from three sensilla in each region,for each of ?ve ?ies (75sensilla total).The proportions of each sensillum type were 36?2,31?3,and 33?4%for pb1,pb2,and pb3,respectively.
Can the functional type of a sensillum be predicted from its coordinates on the surface of the maxillary palp?We addressed this question by analyzing a group of three sensilla that can be readily identi?ed by virtue of their proximity to a cluster of large mechanosensory bristles on the proximolateral edge of the sen-sory ?eld.Two organizational principles emerged from this anal-ysis.First,the precise positions of individual sensilla were not ?xed;the relative positions of the three sensilla to each other and their positions relative to the mechanosensory bristles varied among the four animals examined.Second,the identities of indi-vidual sensilla of this group were not the same among the four
?ies examined;although all three sensilla were either pb1or pb3in every case (the region lies within the pb2exclusion zone),the fraction of sensilla that were of the pb1type ranged from zero of three to three of three.These results indicate that the identity of a sensillum is not strictly determined by its position.
All three sensillum types are found on both male and female palps,and we found no evidence of sexual dimorphism of any kind.A quantitative analysis of spike frequency showed no dif-ferences between the sexes in the response of pb1sensilla (n ?9sensilla of each sex)to any of seven odors tested (ethyl acetate,isoamyl acetate,4-methylphenol,benzaldehyde,3-octanol,E2-hexenal,and cyclohexanone);limited data revealed no differences between the sexes for pb2or pb3sensilla.These results are consistent with the lack of sexual dimorphism in the glomeruli of the antennal lobes (Stocker,1994)and form a striking contrast with the dimorphism observed in the olfactory sensilla and glo-meruli of moths (Schneiderman and Hildebrand,1985).
Adaptation and the independence of neurons within a sensillum
The response of an olfactory neuron may depend not only on the chemical structure (quality)and dose (quantity)of an odor stim-ulus but also on the previous experience of the neuron,via the process of adaptation.The effect of experience on the pb1A neuron is shown in Figure 11.First,during the course of a sustained odor stimulus,the frequency of action potentials changes (Fig.11A ).The spike frequency quickly rises to a peak of 200spikes/sec and then rapidly declines to 25%of its peak value,over a period of 1sec;it then slowly declines over the ensuing 24sec to 15%of its peak value.Second,this 25sec stimulus affects the response to a subsequent stimulus.A second stimulation with the same odor elicited a peak response only 60%of the initial peak response (Fig.11A ).The magnitude of the second response depends critically on the recovery time,that is,the time between the end of the ?rst 25sec stimulus and the onset of the second stimulus.The cell recovers from adaptation relatively fast (Fig.11B ),and sensitivity is approximately a linear function of the logarithm of the recovery time.After 1sec,the responsiveness is ?50%of that before adaptation.Full recovery is achieved after 100sec.We note that full recovery from 500msec stimuli is much faster than that from 25sec stimuli (?5sec;data not shown)Olfactory neurons of the maxillary palp also exhibit cross-adaptation.The peak response of an unadapted pb1A to an ethyl acetate stimulus is 250spikes/sec (Fig.11C ),but when the ethyl acetate stimulus is administered 1sec after a prolonged stimulus of ethyl propionate,the response to ethyl acetate is reduced to 125spikes/sec,50%of the value for the unadapted neuron.
Having documented adaptation and cross-adaptation in these neurons,we were then in a position to investigate whether the experience of one neuron in a sensillum affects the neighboring neuron in the same sensillum.The anatomical relationship of these two neurons is intimate.Their outer dendrites are inter-twined in a sensillum whose diameter is only 10?m,and their inner dendrites and cell bodies are closely apposed,wrapped within a single sheath cell (Singh and Nayak,1985).It has been suggested that there may be communication between adjacent neurons,perhaps mediated by the intercellular second messenger nitric oxide (Breer and Shepherd,1993).To test the possibility of one form of intercellular communication,we stimulated pb1sen-silla with ethyl propionate,an odor that stimulates pb1A but not pb1B,and then tested whether this stimulation of pb1A affected the subsequent response of pb1B to 4-methylphenol,an odor
that
Figure 10.The distribution of the three sensillum types,all of which reside on the dorsal (top)and lateral surfaces of the maxillary palp.Each colored circle represents one recording from a basiconic sensillum.Clusters of circles indicate recordings made from sensilla in corresponding posi-tions on different ?ies.The total number of recordings from each sensillum type are indicated.The arrow points to a zone where pb2sensilla are virtually absent.
4528J.Neurosci.,June 1,1999,19(11):4520–4532de Bruyne et al.?Odor Coding in Drosophila Palps
stimulates pb1B but not pb1A.Figure 11D shows that the pb1B neuron was unaffected by previous stimulation of pb1A;its re-sponse to 4-methylphenol was 101?7%of the pb1B value before the prolonged exposure of the sensillum to ethyl propionate.By contrast,adaptation of pb1A to ethyl propionate affected the subsequent response of pb1A to ethyl propionate,ethyl acetate,E2-hexenal,and isoamyl acetate (all of which excite pb1A but not pb1B);the subsequent responses were all reduced to ?40%of the level before adaptation.We conclude that there is both adapta-tion and cross-adaptation within the pb1A neuron.However,this form of adaptation does not extend from the pb1A neuron to the pb1B neuron;the two neurons in a sensillum function as inde-pendent units in this respect.
DISCUSSION
We have performed a high-resolution functional analysis of an entire olfactory organ.Extensive recordings from the 60sensilla of the Drosophila maxillary palp have revealed that its 120neu-rons fall into six functional classes,each with a different response
spectrum.It seems likely that all the neuronal types of this model olfactory organ have now been de?ned.Analysis of these neurons has revealed some basic principles of their function and organi-zation,suggesting some of the fundamental elements with which olfactory coding can be achieved.
Odor coding in a model olfactory organ
We have de?ned the six neuronal classes with a panel of odors that,although chemically diverse,represents only a very small sampling of odor space.Our purpose was to distinguish among neuronal types,not to identify those ligands that elicit the stron-gest responses from individual neurons.ORNs in other systems have been characterized as having either broadly or narrowly tuned responses (Derby and Ache,1984;Sicard and Holley,1984;Boeckh et al.,1987).We have shown that in the case of the pb1A neuron the apparent breadth of tuning depends on the concen-trations of odorants.When tested with odorants at higher doses (10?2dilutions),pb1A appears to be broadly tuned (Fig.5),but when tested with odorants at lower doses (e.g.,10?4),it
appears
Figure 11.Adaptation and cross-adaptation in pb1sensilla show convergence of olfactory information in pb1A neurons but independence of two neurons in a sensillum.A ,Frequency of action potentials (averaged over 120msec intervals)during prolonged exposure of pb1A to ethyl propionate is shown.The long stimulation (25sec)was generated using the ?ask ?ow method (10?2dilution),whereas the ensuing short stimulus (500msec)was delivered with the syringe puff method (10?3.5dilution;see Materials and Methods).Both stimulations elicit the same spike frequency during the ?rst 500msec from unadapted neurons.Note that frequencies over 500msec are not the same as peak responses averaged over 120msec.B ,The response to the second stimulation with ethyl propionate increases,approximately following a linear function of the logarithm of the recovery time after adaptation.Responses are presented as a percentage of the response to an identical 500msec stimulus given before adaptation.C ,The pb1A neuron cross-adapts to ethyl acetate,after stimulation with ethyl propionate.A 10?2dilution of ethyl acetate and a 10?3.5dilution of ethyl propionate were used,both using the syringe puff method,because they elicit the same response from pb1A (see Fig.9).D ,The pb1A neuron was adapted to ethyl propionate as in A .Then the response was measured from pb1B to 4-methylphenol (10?2dilution;n ?13)or from pb1A to ethyl propionate (10?3.5dilution;n ?10),ethyl acetate (10?2dilution;n ?8),E2-hexenal (10?1.5dilution;n ?8),or isoamyl acetate (10?1dilution;n ?7).All the doses used were determined from Figure 9to give approximately equal responses before adaptation.Error bars indicate SEM.Cross-adaptation is observed for all odorants exciting pb1A but not for 4-methylphenol,which stimulates pb1B.
de Bruyne et al.?Odor Coding in Drosophila Palps J.Neurosci.,June 1,1999,19(11):4520–45324529
narrowly tuned and responds only to ethyl propionate among the tested odors(Fig.9).It is likely that there exist other odorants to which some of the cells described here,particularly pb3A,are more sensitive.We note that we do not know how the stimulus concentrations used in this study compare with those experienced in the wild.At the doses we used,some odors,such as 4-methylphenol,strongly stimulate two neuronal types.The stim-ulation of multiple neurons by a single odor is consistent with a model in which odor quality is assessed by integrating informa-tion from multiple neuronal types.
We have also shown that odor coding at the periphery in Drosophila involves both excitatory and inhibitory responses.One of the neuronal types,p2B,exhibits two modes of response:it is excited by some stimuli and inhibited by others.Moreover,the extent of inhibition varies with different odor stimuli.It seems likely that the excitation and inhibition that we have observed re?ect depolarizing and hyperpolarizing ion currents in the den-drites of the ORNs.The occurrence of such responses in an individual ORN has been demonstrated in insects(Boeckh,1967; Dubin and Harris,1997)and crustaceans(Michel and Ache, 1994).In the latter,depolarizing and hyperpolarizing currents are mediated via the IP3and cAMP transduction pathways, respectively(Ache and Zhainazarov,1995).The existence of two modes of response suggests the possibility that multiple messages can be sent from the same neuron;it provides an additional degree of freedom and expands the possible means by which odor coding can be achieved with a limited number of sensory neurons. Not only can an individual neuron be excited by one odor and inhibited by another,but an individual odor can excite one cell and inhibit another.For example,benzaldehyde stimulates pb2A but inhibits pb2B.The simultaneous production of opposite ef-fects on two neurons may provide a means of generating contrast, which may enhance the recognition of speci?c odors.
We have also found that different odors can have different effects on the kinetics of the response.For example,?ring of the pb1b and pb2b neurons stimulated with4-methylphenol continues long past the end of the stimulus(Fig.4B,C),whereas?ring of other neurons with other odors terminates at the end of the stimulus period(Fig.4A,E).Phenols also elicit prolonged?ring of receptor cells in tsetse?y antennae(Den Otter and Van der Goes van Naters,1993),but such extended tonic responses are not limited to phenols;they have also been observed in moth neurons after stimulation with pheromones(Almaas et al.,1991).Thus the temporal pattern of action potentials provides another degree of freedom:the time course of spikes may contain information aiding in odor recognition.
Neural organization of the maxillary palp and
its development
The number of functional types of neurons on the maxillary palp, six,is of the same order as the estimated number of glomeruli that receive afferent?bers from it:?ve,as reported by Singh and Nayak(1985),or three,as estimated by Stocker(1994).This approximate numerical equivalence,in which the number of neuronal classes is determined by direct physiological analysis,is consistent with the approximate equivalence of the number of glomeruli with the number of neuronal types in the mammalian main olfactory epithelium,in which the number of neuronal types is estimated on the basis of an analysis of receptor gene expres-sion(Mombaerts et al.,1996).We note that in moths,there is also a well documented pattern of projection from particular func-tional classes of neurons,pheromone-sensitive neurons,to a clus-ter of specialized glomeruli,the macroglomerular complex (Hansson et al.,1992;Hildebrand and Shepherd,1997).However, it remains to be seen whether the two neurons in each sensillum type project to one glomerulus,giving three palpal glomeruli,or to two glomeruli,which would give six palpal glomeruli.The total number of olfactory glomeruli reported for the antennal lobe of Drosophila is?40(Laissue et al.,1999).Hence the total periph-eral input to the olfactory-processing centers in the CNS may consist of?40basic types of input elements.
A major way in which the insect olfactory system differs from that of vertebrates is that the neurons of the sensory?eld are compartmentalized in sensilla.The insect olfactory system can in this respect be considered as a“compound nose,”by analogy to the compound eye(Hekmat-Scafe et al.,1998).We have found that neurons of the palp are ordered with respect to this level of organization;the six types of neurons are distributed within sensilla in stereotyped pairs,with a neuron of one particular response spectrum cohabiting in a sensillum with a neuron of another particular response spectrum.Despite the intimate co-habitation of neurons within a sensillum,our cross-adaptation experiments show that they are able to function primarily as independent units of perception.This independence and their distinct response spectra are observed despite the fact that both neurons in a sensillum share a common pool of binding proteins (Hekmat-Scafe et al.,1997)and a common electrical circuit (Kaissling,1987).In this sense,olfactory tissues in insects may be similar to those in mammals.
The largely overlapping distribution of the three sensillum types across the sensory?eld shows that in the maxillary palp there is not an obvious odotopic layout at the primary neuron level.However,the presence of a pb2“exclusion”zone provides some heterogeneity and likely points to zones that are develop-mentally distinct.Our analysis of sensillum function and organi-zation raises interesting questions about how such a sensory?eld develops.Olfactory sensilla develop from founder cells(Ray and Rodrigues,1995)that build different sensillum categories.The mixed distribution of functional types on the palp and the vari-ability in the positions of individual sensilla suggest that sensilla are not committed to one of the three alternative fates strictly according to their position in the?eld.We do not know how the expression of genes encoding the choice between pb1,pb2,and pb3is regulated.Moreover,further studies will be necessary to reveal the developmental logic by which the stereotyped pairing of neurons is produced.What is the mechanism that regulates expression of class-speci?c elements such as receptors and coor-dinates it between the two neurons of one sensillum?
The palp as an accessory olfactory organ
The olfactory system of adult Drosophila contains two organs,the antenna and the maxillary palp.Each antenna is covered with ?500sensilla of three morphological types,whereas the maxillary palp is covered with60sensilla of a single morphological type.It is tempting to compare this arrangement with the presence of two olfactory organs in mammals,the main olfactory epithelium (MOE)and the vomeronasal organ(VNO).Like the VNO,the maxillary palp has been associated with pheromone response and the modulation of sexual behavior(Stocker and Gendre,1989). The VNO neurons project to an accessory olfactory bulb that does not overlap with the main olfactory bulb(Farbman,1992; Dulac and Axel,1995);likewise,projections from the palp have been reported to map to a subset of glomeruli distinct from those that receive input from the antenna(Stocker,1994;cf.Singh and
4530J.Neurosci.,June1,1999,19(11):4520–4532de Bruyne et al.?Odor Coding in Drosophila Palps
Nayak,1985).However,VNO neurons differ from MOE neurons in that they are microvillous rather than ciliate(Farbman,1992), whereas both palpal and antennal ORNs are ciliate(Singh and Nayak,1985).In addition,the VNO neurons probably perceive a different type of odorants(less volatile and larger molecules) from those perceived by the MOE,whereas the present study shows that the maxillary palp is sensitive to many small,volatile molecules that also stimulate the antenna(this study)(Siddiqi, 1991;Ayer and Carlson,1992).We also have found no evidence of a special role of the palp in pheromone perception.First, cis-vaccenyl acetate,a compound reported previously as an inhib-itory sex pheromone(Jallon,1984),induces a response from pb1A.However,this neuron responds more strongly to many other odors,and cis-vaccenyl acetate has also been found to stimulate an antennal neuron(Clyne et al.,1997).Second,we did not?nd differences between male and female ORN populations, making it unlikely that sex-speci?c pheromonal receptor cells occur on the palps.
In summary,we have provided an extensive description of an olfactory organ,compiling what is likely a complete catalog of its neural elements.We have shown that olfactory information in this organ is encoded by a limited number of ORN classes. Furthermore,this study,taken together with previous studies (Siddiqi,1991;Clyne et al.,1997),suggests that the total number of ORN response classes in the entire olfactory system is limited and identi?able.Although a detailed roster of olfactory neurons of the worm Caenorhabditis elegans has been made available via behavioral analysis(Bargmann et al.,1993),it has not been possible to identify exhaustively all the neuronal types of olfac-tory systems of higher organisms on account of their complexity. Physiological analysis of the receptor neurons of the Drosophila palp has also revealed a rich diversity of response dynamics, which together with the existence of neuron classes establishes a basis for olfactory coding.In addition to principles of functional organization,the present study provides a foundation for analysis of the molecular and genetic mechanisms that underlie the func-tion and development of this model olfactory organ. REFERENCES
Ache BW,Zhainazarov A(1995)Dual second-messenger pathways in olfactory transduction.Curr Opin Neurobiol5:461–466.
Almaas TJ,Christensen TA,Mustaparta H(1991)Chemical communi-cation in heliothine moths.I.Antennal receptor neurons encode several features of intra-and interspeci?c odorants in the male corn earworm moth Helicoverpa zea.J Comp Physiol[A]169:249–258.
Altner H,Prillinger L(1980)Ultrastructure of invertebrate chemo-, thermo-and hygroreceptors and its functional signi?cance.Int Rev Cytol67:69–139.
Anderberg MR(1973)Cluster analysis for applications.New York: Academic.
Anderson P,Hansson BS,Lo¨fqvist J(1995)Plant-odour-speci?c recep-tor neurones on the antennae of female and male Spodoptera littoralis. Physiol Entomol20:189–198.
Ayer R,Carlson JR(1992)Olfactory physiology in the Drosophila an-tenna and maxillary palp:acj6distinguishes two classes of odorant pathways.J Neurobiol23:965–982.
Bargmann CI,Hartwieg E,Horvitz HR(1993)Odorant-selective genes and neurons mediate olfaction in C.elegans.Cell74:515–527. Bieber SL,Smith DV(1986)Multivariate analysis of sensory data:a comparison of methods.Chem Senses11:19–47.
Boeckh J(1967)Inhibition and excitation of single insect olfactory re-ceptors,and their role as a primary sensory code.In:Olfaction and taste(Hayashi T,ed),pp721–735.Oxford:Pergamon.Boeckh J(1981)Chemoreceptors:their structure and function.In: Sense organs(Laverack MS,Cosens PJ,eds),pp86–99.London: Blackie.
Boeckh J,Ernst K-D,Selsam P(1987)Neurophysiology and neuroanat-omy of the olfactory pathway in the cockroach.In:Olfaction and taste, Vol IX(Roper SD,Atema J,eds),pp39–43.New York:New York Academy of Sciences.
Breer H,Shepherd GM(1993)Implications of the NO-cGMP system for olfaction.Trends Neurosci16:5–9.
Buck LB(1996)Information coding in the vertebrate olfactory system. Annu Rev Neurosci19:517–544.
Buck LB,Axel R(1991)A novel multigene family may encode odorant receptors:a molecular basis for odor recognition.Cell65:175–187. Bursell E,Gough AJE,Beevor PS,Cork A,Hall DR,Vale GA(1988) Identi?cation of components of cattle urine attractive to tsetse?ies, Glossina spp.(Diptera:Glossinidae).Bull Entomol Res78:281–291. Carlson JR(1996)Olfaction in Drosophila:from odor to behavior. Trends Genet12:175–180.
Clyne P,Grant AJ,O’Connell RJ,Carlson JR(1997)Odorant response of individual sensilla on the Drosophila antenna.Invertebrate Neurosci 3:127–135.
Den Otter CJ,Van der Goes van Naters WM(1993)Responses of individual olfactory cells of tsetse?ies(Glossina m.morsitans)to phenols from cattle urine.Physiol Entomol18:43–49.
Derby CD,Ache BW(1984)Quality coding of a complex odorant in an invertebrate.J Neurophysiol51:906–924.
Dubin AE,Harris GL(1997)Voltage-activated and odor-modulated conductances in olfactory neurons of Drosophila melanogaster.J Neu-robiol32:123–137.
Dulac C,Axel R(1995)A novel family of genes encoding putative pheromone receptors in mammals.Cell83:195–206.
Farbman AI(1992)Cell biology of olfaction.Cambridge,UK:Cam-bridge UP.
Fujishiro N,Kijima H,Morita H(1984)Impulse frequency and action potential amplitude in labellar chemosensory neurons of Drosophila melanogaster.J Insect Physiol30:317–325.
Getz WM,Akers RP(1997)Response of American cockroach(Peripla-neta americana)olfactory receptors to selected alcohol odorants and their binary combinations.J Comp Physiol[A]180:701–709.
Guillet JC,Bernard J(1972)Shape and amplitude of the spikes induced by natural or electrical stimulation in insect receptors.J Insect Physiol 18:2155–2171.
Hansson BS(1995)Olfaction in Lepidoptera.Experientia51:1003–1027. Hansson BS,Ljungberg H,Hallberg E,Lo¨fstedt C(1992)Functional specialization of olfactory glomeruli in a moth.Science256:1313–1315. Hansson BS,Hallberg E,Lo¨fstedt C,Steinbrecht RA(1994)Correlation between dendrite diameter and action potential amplitude in sex pher-omone speci?c receptor neurons in male Ostrinia nubilalis(Lepidop-tera:pyralidae).Tissue Cell26:503–512.
Hekmat-Scafe DS,Steinbrecht RA,Carlson JR(1997)Coexpression of two odorant-binding protein homologs in Drosophila:implications for olfactory coding.J Neurosci17:1616–1624.
Hekmat-Scafe DS,Steinbrecht RA,Carlson JR(1998)Olfactory coding in a compound nose,coexpression of odorant-binding proteins in Dro-sophila.In:Olfaction and taste,Vol XII,An international symposium (Murphy C,ed),pp311–315.New York:New York Academy of Sciences.
Helfand SL,Carlson JR(1989)Isolation and characterization of an olfactory mutant in Drosophila with a chemical speci?c defect.Proc Natl Acad Sci USA86:2908–2912.
Hildebrand JG,Shepherd GM(1997)Mechanisms of olfactory discrim-ination:converging evidence for common principles across phyla.Annu Rev Neurosci20:595–631.
Jallon J-M(1984)A few chemical words exchanged by Drosophila during courtship and mating.Behav Genet14:441–478.
Kaissling K-E(1987)R.H.Wright lectures on insect olfaction.Burnaby, British Columbia,Canada:Simon Fraser University.
Kaissling K-E(1995)Single unit and electroantennogram recordings in insect olfactory organs.In:Experimental cell biology of taste and olfaction,current techniques and protocols(Spielman AI,Brand JG, eds),pp361–377.Boca Raton,FL:CRC.
Laissue PP,Reiter C,Hiesinger PR,Halter S,Fishbach K-F,Stocker RF (1999)3D reconstruction of the antennal lobe in Drosophila melano-gaster.J Comp Neurol405:543–552.
Loftus R,Corbie`re-Tichane′G(1981)Antennal warm and cold receptors
de Bruyne et al.?Odor Coding in Drosophila Palps J.Neurosci.,June1,1999,19(11):4520–45324531
of the cave beetle,Spheophyes lucidulus Delar.,in sensilla with a lamellated dendrite.I.Response to sudden temperature change. J Comp Physiol[A]143:443–452.
Michel WC,Ache BW(1994)Odor-evoked inhibition in primary olfac-tory neurons.Chem Senses19:11–24.
Mombaerts P,Wang F,Dulac C,Chao SK,Nemes A,Mendelsohn M, Edmondson J,Axel R(1996)Visualizing an olfactory sensory map. Cell87:675–686.
Ray K,Rodrigues V(1995)Cellular events during development of ol-factory sense organs in Drosophila melanogaster.Dev Biol167:426–438. Riesgo-Escovar JR,Piekos WB,Carlson JR(1997)The maxillary palp of Drosophila:ultrastructural and physiology depends on the lozenge gene.J Comp Physiol[A]180:143–150.
Rumbo ER(1989)The shape of extracellularly recorded nerve impulses from pheromone receptors.Chem Senses14:361–369. Schneiderman AM,Hildebrand JG(1985)Sexually dimorphic develop-ment of the insect olfactory pathway.Trends Neurosci8:494–499. Sicard G,Holley A(1984)Receptor cell responses to odorants:similar-ities and differences among odorants.Brain Res292:283–296.Siddiqi O(1991)Olfaction in Drosophila.In:Chemical senses,Vol3, Genetics of perception and communications(Wysocki CJ,Kare MR, eds),pp79–96.New York:Dekker.
Singh RN,Nayak SV(1985)Fine structure and primary sensory projec-tions of sensilla on the maxillary palp of Drosophila melanogaster Meigen(Diptera:Drosphilidae).Int J Insect Morphol Embryol 14:291–306.
Stocker RF(1994)The organization of the chemosensory system in Drosophila melanogaster:a review.Cell Tissue Res275:3–26. Stocker RF,Gendre N(1989)Courtship behavior of Drosophila geneti-cally or surgically deprived of basiconic sensilla.Behav Genet 19:371–385.
Taneja J,Guerin PM(1997)Ammonia attracts the haematophagous bug Triatoma infestans:behavioural and neurophysiological data on nymphs.J Comp Physiol[A]181:21–34.
Visser JH(1986)Host odour perception in phytophagous insects.Annu Rev Entomol31:121–144.
Ward Jr JH(1963)Hierarchical grouping to optimize an objective func-tion.J Am Stat Assoc58:236–244.
4532J.Neurosci.,June1,1999,19(11):4520–4532de Bruyne et al.?Odor Coding in Drosophila Palps
儿童歌谣大全 各位读友大家好,此文档由网络收集而来,欢迎您下载,谢谢 篇一:儿童歌谣歌词大全 [小螺号]儿歌歌词 小螺号,嘀嘀嘀吹海鸥听了展翅飞小螺号,嘀嘀嘀吹浪花听了笑微微小螺号,嘀嘀嘀吹 声声唤船归啰 小螺号,嘀嘀嘀吹 阿爸听了快快回啰。 茫茫的海洋,蓝蓝的海水 吹起了小螺号,心里美吔 [小鸟小鸟] 蓝天里有阳光,树林里有花香 小鸟小鸟,你自由地飞翔 在田野,在草地,在湖边,在山冈 小鸟小鸟迎着春天歌唱 啦啦啦啦啦。爱春天,爱阳光,爱湖水,爱花香小鸟小鸟,我的好朋友让我们一起飞翔歌唱
一起飞翔歌唱 啦啦啦啦啦 [让我们荡起双桨] 让我们荡起双桨,小船儿推开波浪海面倒映着白塔,四周环绕着绿树红墙小船儿轻轻飘荡在水中 迎面吹来了凉爽的风 红领巾迎着太阳,阳光洒在海面上 水中鱼儿望着我们 悄悄听我们愉快歌唱 小船儿轻轻飘荡在水中 迎面吹来了凉爽的风 做完了一天的功课,我们来尽情欢乐 我问你亲爱的伙伴 谁给我们安排下幸福的生活? 小船儿轻轻飘荡在谁中 迎面吹来了凉爽的风 [小红花]儿歌歌词 金波词尚疾曲 花园里,篱笆下,我种下一朵小红花
春天的太阳当头照,春天的小雨沙沙下 啦啦啦啦啦,啦啦啦啦啦 小红花张嘴笑哈哈 花园里,篱笆下,我种下一朵小红花 春天的太阳当头照,春天的小雨沙沙下 啦啦啦啦啦,啦啦啦啦啦 小红花张嘴笑哈哈[我家几口] 金苗苓词曲我家有几口? 让我扳指头 爸爸,妈妈,还有我 再加一个布娃娃 哟!有四口我家有几口? 让我扳指头 爸爸,妈妈,还有我 再加一个布娃娃 哟!有四口 [谁会这样] 少数民族儿歌潘振声曲 谁会飞呀,鸟会飞
鸟儿鸟儿怎样飞? 拍拍翅膀飞呀飞 谁会游呀,鱼会游 鱼儿鱼儿怎样游? 摇摇尾巴点点头 谁会跑呀,马会跑马儿马儿怎样跑?四脚离地身不摇。[我叫轻轻] 张友珊词汪玲曲 走路轻轻轻轻 上夜班的阿姨还没醒呀 敲门轻轻轻轻 给邻居叔叔送呀送封信 说话轻轻轻轻 姐姐灯下看书多用心呀 大家夸我是好孩子 给我取个名字叫呀叫轻轻 走路轻轻轻轻 上夜班的阿姨还没醒呀 敲门轻轻轻轻 给邻居叔叔送呀送封信 说话轻轻轻轻 姐姐灯下看书多用心呀
56首经典儿歌歌词大全 1、做早操 早上空气真叫好,我们都来做早操。 伸伸臂,弯弯腰,踢踢腿,蹦蹦跳,天天锻炼身体好。 2、饭前要洗手 小脸盆,水清请,小朋友们笑盈盈,小手儿,伸出来, 洗一洗,白又净,吃饭前,先洗手,讲卫生,不得病。 3、小手绢 小手绢,四方方,天天带在我身上。 又擦鼻涕又擦汗,干干净净真好看。 4、搬鸡蛋 小老鼠,搬鸡蛋,鸡蛋太大怎么办?一只老鼠地上躺, 紧紧抱住大鸡蛋。一只老鼠拉尾巴,拉呀拉呀拉回家。 5、大骆驼 骆驼骆驼志气大,风吹日晒都不怕。 走沙漠,运盐巴,再苦再累不讲话。 6、螳螂 螳螂哥,螳螂哥,肚儿大,吃得多。飞飞能把粉蝶捕, 跳跳能把蝗虫捉。两把大刀舞起来,一只害虫不放过 7、大蜻蜓 大蜻蜓,绿眼睛,一对眼睛亮晶晶, 飞一飞,停一停,飞来飞去捉蚊蝇。 8、小鸭子 小鸭子,一身黄,扁扁嘴巴红脚掌。 嘎嘎嘎嘎高声唱,一摇一摆下池塘。 9、拍手歌 你拍一,我拍一,天天早起练身体。 你拍二,我拍二,天天都要带手绢。 你拍三,我拍三,洗澡以后换衬衫。 你拍四,我拍四,消灭苍蝇和蚊子。 你拍五,我拍五,有痰不要随地吐。 你拍六,我拍六,瓜皮果核不乱丢。 你拍七,我拍七,吃饭细嚼别着急。 你拍八,我拍八,勤剪指甲常刷牙。 你拍九,我拍九,吃饭以前要洗手。
你拍十,我拍十,脏的东西不要吃。 10 、小螃蟹 小螃蟹,真骄傲,横着身子到处跑, 吓跑鱼,撞倒虾,一点也不懂礼貌 11 、庆六一 儿童节,是六一,小朋友们真欢喜。 又唱歌来又跳舞,高高兴兴庆六一。 12、花猫照镜子 小花猫,喵喵叫,不洗脸,把镜照, 左边照,右边照,埋怨镜子脏,气得胡子翘。 13、蚂蚁搬虫虫 小蚂蚁,搬虫虫,一个搬,搬不动,两个搬,掀条缝, 三个搬,动一动,四个五个六七个,大家一起搬进洞。 14、小青蛙 小青蛙,呱呱呱,水里游,岸上爬, 吃害虫,保庄稼,人人都要保护它。 15、花儿好看我不摘 公园里,花儿开,红的红,白的白, 花儿好看我不摘,人人都说我真乖。 16 、红绿灯 大马路,宽又宽,警察叔叔站中间, 红灯亮,停一停,绿灯亮,往前行。 17 、七个果果 一二三四五六七,七六五四三二一。 七个阿姨来摘果,七个篮子手中提。七个果子摆七样。 苹果、桃儿、石榴、柿子、李子、栗子、梨。 18、睡午觉 枕头放放平,花被盖盖好。 小枕头,小花被,跟我一起睡午觉,看谁先睡着。 19 、吃荸荠 荸荠有皮,皮上有泥。洗掉荸荠皮上的泥,削去荸荠外面的皮,荸荠没了皮和泥,干干净净吃荸荠。 20 、小云骑牛去打油 小云骑牛去打油,遇着小友踢皮球,皮球飞来吓了牛,摔下小云撒了油。 21 、盆和瓶 车上有个盆,盆里有个瓶,乒乒乒,乓乓乓,不知是瓶碰盆,还是盆碰瓶。
一、餐饮类 1、吃饭歌谣 铃….下课了,小朋友们开饭了,洗洗手,不打闹,安安静静吃个饱,不乱吃,不挑食,身体倍棒好宝宝。 2、自由吃饭 好宝宝,吃饭了,小饭碗,手扶好,小勺子,快拿好,绿青菜,红大虾,细细嚼,慢慢咽,啊呜啊呜吃个饱。 3、吃饼干 饼干圆圆,圆圆饼干,用手掰开,变成小船,你吃一半,我吃一半,啊呜一口,小船真甜。 4、大苹果 我是一个大苹果,小朋友们都爱我,请你先洗洗手,要是手脏别碰我。 5 我的性格有点怪,餐餐吃饭爱挑菜,老师天天教育我,吃饭不能把菜挑,影响健康害处大,现在的我真可爱,吃饭再也不挑菜。 6、宝宝吃饭不用喂 小花碗,竹筷子儿,盛好饭,自己端,不挑食,不浪费,细细嚼,慢慢咽,宝宝吃饭不用喂。 7、小宝宝,坐坐好,妈妈盛饭喂宝宝,细细嚼,慢慢咽,宝宝吃的真叫好。 8吃饭了 小朋友们快坐好,我们马上开饭了,幼儿园里饭菜香,既讲卫生又营养,小朋友们要记住,吃饭不要把话讲,蔬菜猪肉都要吃,豆腐鸡蛋也咽下,细嚼慢咽来吃饭,身体长得高又壮。 9、大老虎 大白米饭,豆腐鸡蛋,青菜肉汤,喷香喷香,我是一只大老虎,啊呜… 10、吃饭不挑剔 小羊爱吃草,小鸡爱吃米,小兔爱吃菜,小猫爱吃鱼,我是好孩子,吃饭不挑食。11、吃饭不挑食 米饭白白馒头香,窝头尖尖面条长,萝卜白 菜营养好,吃得多来长得壮,我们吃饭不挑食,爸爸妈妈都夸奖。 12、兔兔吃饭 小兔兔,吃饭罗,洗小手,高高坐,爸爸来 吃馍馍,妈妈来吃肉肉,我吃青菜又吃肉, 又吃馒头又喝粥,吃得快,真不错,小兔一 家真快活! 13、开饭了 开饭了,开饭了,妈妈盛饭我端碗,桌椅餐 具摆摆好,小心谨慎要安全,香喷喷,白米饭,先给爷爷和奶奶,再给爸爸和妈妈,最 后放到我面前。 14、 小餐桌,四方方,宝宝吃饭坐桌旁,一口饭,一口菜,细嚼慢咽无声响,青菜鱼肉都爱吃,不挑食,身体壮。 15、 吃饭时,勿嬉戏,谨防饭食噎口里。细细嚼,慢慢咽,嘴巴出声不雅观。饭菜好,莫暴食,每餐只吃八分饱,碗里饭菜吃干净。 16、 吃饭前,先邀请,大家一起来开动。第一筷长辈吃,好饭菜要礼让。高举手,轻落下,筷勺上面要干净。夹菜要从面前夹,不要满盘来挑拣。 17、 吃饭时,坐端正,右手拿调羹,左手扶着碗。细细嚼,慢慢咽,不剩饭,不挑菜,自己吃饭真能干,我是园里的好伙伴。 18乖宝宝 好宝宝,乖宝宝,吃饭前,把手洗,正面洗,背面洗,洗干净,讲卫生,宝宝乖,宝宝棒,不挑食,不浪费,人人都喜爱。
《第56号教室的故事》读后感 2020-07-18 当仔细品读一部作品后,你有什么体会呢?此时需要认真思考如何写了哦。那么读后感到底应该怎么写呢?下面是小编为大家收集的《第56号教室的故事》读后感,欢迎阅读,希望大家能够喜欢。 利用闲暇的时间,我拜读了雷夫先生的《第56号教室的故事》,在教育教学中深有启发。 (一)特殊的理财课。 在开学的第一天,雷夫先生告诉孩子们,必须在班里承担一项工作,可以是“银行家”、“清洁工”、“文秘”,等等。承担并完成了这些工作之后,每个月底他给孩子们发“工资”,当然不是真的钱,但可以在教室里使用。 启发:我们班级也在搞班级百分制,虽然百分制和孩子们每周之星评比和综合素质评定等有一定的关系,但随着时间的流逝,孩子们对班级百分制会产生一定的倦怠,百分制对孩子们的刺激也不如初一时来得这么强烈。我在想,如果按照雷夫先生的方法,将千分制换成具体的“工资”形式,是否能让孩子在学习、工作等方面的积极性更强,同时也能指导他们正确理财,既锻炼了孩子们的自立自强能力,又培养了孩子们的责任意识,让孩子们的学习生活和社会生活有了更密切的联系,这真是一举多得啊! (二)抓中间,而非抓两头。 雷夫先生说:“在我的班级里,我总是在脑子里把学生大致分成三类:第一类孩子、第二类孩子和第三类孩子。” 这是一般老师的普遍的分法,而雷夫先生对待这三类孩子的秘诀在于把更多的时间花在第二类孩子身上。这一秘诀让我眼前一亮。的确,在以往的教学中我们喜欢第一类孩子,但把更多的时间花在了第三类孩子身上,而第二类孩子却往往受到忽视。事实上,在第二类孩子身上花时间是最有成效的。 在雷夫老师这一观点的启发下,我想每一个孩子都应该得到发展,在不同的时间,不同的场合,对这三类孩子的关注强度各不一样,为使每一个孩子都得到各自的发展,我认为作为老师,应该为他们的'发展创造条件。比如说位置的安排。七年级,为了班级的稳定与和谐,为了小学的孩子能尽快适应初中的生活,我们的位置习惯采取“互补”的形式,特别是习惯好,成绩优异的同学,他们的同桌相对习惯较差,成绩较弱。在班级初始阶段,这样的安排的确有利于保证课堂的纪律,形成良好的班级学习氛围,对习惯差学习能力弱的同学也有一定的榜样作用。但久而久之,学习成绩优异能力强的同学会找不到合
《樱桃小丸子》 小小年纪谈起理想一串串 想当专家、想做博士、想出唱片老爸老妈老师老友都夸赞 想来容易,说来简单,做做就难要数一百,先数一二三 要过明天先过好今天 瞄准目标看齐 噼里啪啦,噼里啪啦 做事不偷懒 噼里啪啦,噼里啪啦
学习不怕难 我们脚踏实地地干 瞄准目标看齐 噼里啪啦,噼里啪啦 读书真勤快 噼里啪啦,噼里啪啦 今天学得好 噼里啪啦,噼里啪啦 明天理想能实现 《多啦A梦》 如果我有仙女棒变大变小变漂亮
还要变个都是漫画巧克力和玩具的家 如果我有机器猫我要叫他小叮当 竹蜻蜓和时光隧道能去任何的地方 让小孩大人坏人都变成好人 (hi 大家好,我是小叮当) ang ang ang小叮当帮我实现所有的愿望 躺在草地上幻想想动想西想玩耍 想到老师还有考试一个头就变成两个大好在我有小叮当困难时候求求他 万能笔和时间机器能做任何的事情 让我的好朋友一齐分享他 (啊!救命啊!有老鼠!) ang ang ang 小叮当帮我实现所有的愿望 躺在草地上幻想想动想西想玩耍 想到老师还有考试一个头就变成两个大好在我有小叮当困难时候求求他 万能笔和时间机器能做任何的事情 让我的好朋友一齐分享他 (小叮当永远是你们的好朋友喔!) ang ang ang 小叮当帮我实现所有的愿望
ang ang ang 小叮当帮我实现所有的愿望 《铁臂阿童木》 越过辽阔天空,啦啦啦飞向遥远群星,来吧!阿童木,爱科学的好少年。善良勇敢的啦啦啦铁臂阿童木,十万马力七大神力,无私无畏的阿童木。穿过广阔大地,啦啦啦潜入深深海洋,来吧!阿童木,爱和平的好少年。善良勇敢的啦啦啦铁臂阿童木,我们的好朋友啊, 无私无畏的阿童木。 《小龙人之歌》 天上有,无数颗星星,那颗最小的就是我,我不知道我从哪里来,也不知道我在哪里生。地上有,无数个龙人,那个最小的就是我,我不知道我从哪里来,也不知道我在哪里生。啊----这是我将在妈妈怀 抱里,啊寻遍天涯,去找他 《我是一条小青龙》我头上有只角,我身后有尾巴,谁也不知道,我有多少秘密?我头上有只角,我身后有尾巴,谁也不知道,我有多少秘密。我是一条小青龙(小青龙,小青龙)我有许多小秘密(小秘
五十首儿歌童谣歌词大全
经典儿童歌谣大全 必背作品: 1、《爸爸瞧妈妈看》 爸爸瞧,妈妈看,宝宝的小手真好看。 爸爸瞧,妈妈看,宝宝的小手看不见。 爸爸妈妈都来看,宝宝的小手又出现。 2.《小花狗捉螃蟹》 小花狗呀尾巴摇,去捉螃蟹真胡闹。一只螃蟹没捉到哟,脚被夹得嗷嗷叫哟。 哎哟哎哟哎哟哎哟哟,你说好笑不好笑? 3、《狗和猴》 桥东走来一条狗,桥西走来一只猴。来到桥心俩碰头,彼此匆匆跑回头。 猴跑几步望望狗,狗跑几步望望猴,究竟猴怕狗,还是狗怕猴。 4、《小鲤鱼》 大大大,小小小,一二三四五六七。 大大大,小小小。哆唻咪发嗦啦西。 小鲤鱼,模样真神气,活蹦乱跳滚了一身泥。 看江河,一望无边际。它说,它说,“我们游过去。” 小鲤鱼,模样真神气,活蹦乱跳滚了一身泥。 看江河,一望无边际。它说,它说,“我们游过去。” 5、《小青蛙》 小青蛙,呱呱呱;水里游,岸上爬;吃害虫,保庄稼;人人都要保护它。 6、《小板凳》 小板凳,真听话,跟我一起等妈妈,妈妈下班回来了,我请妈妈快坐下。7、《老鼠抬花轿》 八只老鼠抬花轿,两只老鼠放鞭炮,四只老鼠来吹号,呜哩哇啦真热闹。 老猫听见来贺喜,一口一口全吃掉。 8、《小白兔过年》 小白兔,过新年。两串红辣椒,挂在门两边,远远看一看,就像红对联。9、《数一数》 一条虫,两条虫,小虫喜欢钻洞洞。三头猪,四头猪,肥猪睡觉打呼噜。 五匹马,六匹马,马儿一跑呱哒哒。七只鸡,八只鸡,公鸡打鸣喔喔啼。 九朵花,十朵花,桃花树下是我家。
25、《老虎和灰兔》 坡上有只大老虎,坡下有只小灰兔;老虎饿肚肚,想吃灰兔兔, 虎追兔,兔躲虎,老虎满坡找灰兔;兔钻窝,虎扑兔,刺儿扎痛虎屁股。 气坏了老虎,乐坏了兔;饿虎肚里咕咕咕,窝里笑坏了小灰兔。 26、《学扣扣》 小牛学扣扣,用手使劲揪;小柳学扣扣,用手来回抠; 小妞学扣扣,对准扣眼扣。小牛、小柳和妞妞,谁学会了扣纽扣? 27、《一位爷爷他姓顾》 一位爷爷他姓顾,上街打醋又买布。买了布,打了醋,回头看见鹰抓兔。 放下布,搁下醋,上前去追鹰和兔。飞了鹰,跑了兔,打翻醋,醋湿布。 28、《雪娃娃》 门口有个雪娃娃,张着嘴巴不说话。 我拿苹果去喂它,叫它不要想妈妈。 29、《小雨点》 小雨点,沙沙沙,落在小河里,青蛙乐得呱呱呱; 小雨点,沙沙沙,落在大树上,大树乐得冒嫩芽; 小雨点,沙沙沙,落在马路上,鞋子啪叽啪叽啪…… 30、《小雪人》 小雪人,白又胖,大眼睛,红鼻头, 头上戴顶歪歪帽,花花围巾套脖上, 哎呀呀,萝卜头没有了,哪去了,准是淘气的小白兔,咬了一口逃跑了。 31、《两只萤火虫》 两只萤火虫,出门找外公。飞到西,飞到东,一头飞进草丛中。 一个说:我要歇歇脚。一个说:我要睡一觉。两只萤火虫,睡到东方红。 外公找不到,恨得气冲冲,只怨自己是懒虫。 32、《三只猫咪》 三只猫咪一起玩,欢欢喜喜捏面团。 大姐捏的小汤圆,二姐捏的大鸭蛋。 小小三姐最能干,捏个老鼠当晚饭。 33、《鹅大哥》 鹅大哥,鹅大哥,红帽子,白围脖, 哦呜、哦呜上山坡。我要问问你,唱的什么歌?
蚕 桑叶嫩,桑叶香, 蚕儿吃,白又胖, 吐银丝,细又长, 织出绸缎做衣裳。 凑“十”歌谣一加九,十只小蝌蚪, 二加八,十只花老鸭, 三加七,十只老母鸡, 四加六,十只金丝猴, 五加五,十只大老虎。 爸爸好作曲:彭野作词:彭野 爸爸好爸爸好爸爸好 爸爸的力气真不小 做的多说的少 脏活累活他全包了 爸爸好爸爸好爸爸好
爸爸的力气真不小 挣的多说的少 剩菜剩饭他全包了 把舞儿跳起来作曲:彭野作词:彭野 把腿儿抬起来呀抬起来 把手儿摆起来呀摆起来 把腰儿扭起来呀扭起来 把头发甩起来呀甩起来 把歌儿唱起来呀唱起来 把舞儿跳起来呀跳起来 把心情放松起来呀放松起来 把笑容露出来呀露出来 多运动身体好身体好呀快长高 高高兴兴最重要健健康康没烦恼 多运动身体好身体好呀快长高 高高兴兴最重要健健康康没烦恼 色彩 作曲:彭野作词:彭野
红色red白色white 绿色green黑色black 粉色pink紫色purple蓝色blue成色orange 红色red白色white 绿色green黑色black 粉色pink紫色purple蓝色blue成色orange 粉色pink紫色purple蓝色blue成色orange 我叫妈妈mother 作曲:彭野作词:彭野 有时候我叫爸爸father 有时候我叫妈妈mother 有时候我叫哥哥brother 有时候我叫姐姐sister 有时候我叫爷爷grandpa 有时候我叫奶奶grandma 有时候我叫叔叔uncle 有时候我叫婶婶aunty 爸爸和妈妈我们是一家 哥哥和姐姐我们是一家 爷爷和奶奶我们是一家 叔叔和婶婶我们是一家 有时候我叫爸爸father
《第56号教室的奇迹》读后感 有了身教的基础,再加上有理的说教,学生才会服从教师的教育,才会更加尊重你,通过阅读《第56号教室的奇迹》,的确让我受益匪浅。 《第56号教室的奇迹》向我们介绍了一位美国传奇的小学教师——雷夫? 艾斯奎斯。他20多年如一日,一直坚守在第56号教室。他是个在教育中能准确找到目标,从不偏离跑道的人。他对教育和学生有信徒般的坚持、父亲般的亲切,还有哲人的敏锐、专家的自信、战士的勇敢——他拥有智慧,拥有力量,所以他创造出奇迹。他不仅是位好老师,也是位好家长。他用他的文字处处提示人们,家庭教育与学校教育的不可分割性。是的!学校教育离不开家庭教育的配合,对于自己孩子在学校的学习情况,家长要做到心中有数,最好的办法就是与老师经常沟通,然后对症下药,这样才能达到目的。对于老师提出的要求,家长要大力支持,如学校的活动,应积极鼓励孩子参与,若需家长参与的活动绝对不要推迟,这样既给孩子做榜样,也支持了老师的工作,同时让自己的孩子也拥有了一个良好的学习环境。 师者,所以传道授业解惑也。通过阅读《第56号教室的奇迹》,我深感到“为人师表”的重任,传道授业解惑,作为人师的职责不必赘述,在学生人生的十字路口,领航者对其世界观、人生观和价值观的形成和培养起着潜移默化的作用。而今,教师职责得到了深化,引导学生,告诉学生前面有一片绚丽的景色,你去努力吧,学生就朝着这个定位方向去了,继而培养出优秀的学生。 一个人可以一辈子不登山,但他的心中一定要有座山,这就是目标。这座山使你总往高处爬,使你总有个奋斗的方向,使你任何一刻抬起头都能看见自己的希望。而我们要培养出什么样的学生?那就是:科学家的思维,外交家的智慧,军事家的勇敢,政治家的胆魄。第56号教室里的每个孩子都展示了他们的勇气、毅力和热情,而这些正是他们的许多同辈所缺少的。这些孩子勇敢地探索着那些很少有人走过的路,世道险恶时他们与人友善,别人放弃时他们令人难以置信地继续努力。这些孩子们不找借口,他们抓住机会,让遇到他们的所有人都看到了未来的希望。这不正是我们所有教师的期望吗? 一个老师可以影响学生的一生,雷夫?艾斯奎斯让我懂得英才是可以造就,只要方法得当,只要充分挖掘出个人的潜能,每个学生都可能成为英才。读《第56号教室的奇迹》让我受益匪浅,在今后的工作中我希望像雷夫?艾斯奎斯那样,用智慧和爱去经营这份高尚、灿烂而又不乏诗意的教育事业!
幼儿儿歌歌词大全 1、小白兔 小白兔乖乖,把门开开,快点开开,我要进来,不开不开就不开,妈妈没回来,谁叫也不开。小白兔乖乖,把门开开,妈妈回来,我要进来,快开快开快快开,妈妈回来了,我来把门开。 2、小花狗 小花狗,真叫脏,不洗脚丫就上床。问它为什么?它说忘,忘,忘! 3、吃饭 小宝宝,坐坐好,妈妈盛饭喂宝宝。细细嚼,慢慢咽,宝宝吃得直叫好。 4、干净 鼻涕擦干净,指甲剪干净,脸儿洗干净,妈妈和和亲一亲。 5、乒乓球 乒乓球,两人打,他给我,我给他,谁都不愿落在自己家. 6、踢毽子 小毽子,小毽子,飞上天, 落下地,我们都来踢踢它,踢不好儿没关系. 7、走夜路 夜色黑,星星闪,小朋友,把家还,突然窜出一只兔,吓得它,满处窜. 8、小盒子 小盒子,作用大,铅笔橡皮装的下, 还有一支小钢笔,装在里面心欢喜, 9、大马路 马路长又宽,我站在中间, 老师过来拜拜手,让我赶快走,说是有危险. 10、木头人
三三三,我们都是木头人, 不许哭来不许笑,还有一个不许动。 11、小皮球,小小篮,落地开花二十一,二五六,二五七, 二八二九三十一;三五六,三五七,三八三九四十一…… 12、小蜜蜂 小蜜蜂,嗡嗡嗡,嗡嗡嗡,大家一起勤劳动, 来匆匆,去匆匆,走得兴味浓,春暖花开不做工, 将来哪里好过冬?嗡嗡嗡,嗡嗡嗡,不学懒惰虫。 13、请你唱个歌吧 小杜鹃,小杜鹃,我们请你唱个歌,快来呀,大家来呀, 我们静听你的歌,咕咕!咕咕!歌声使我们快乐 14、谁会飞 谁会飞呀,鸟会飞,鸟儿鸟儿怎样飞?拍拍翅膀飞呀飞 谁会游呀,鱼会游,鱼儿鱼儿怎样游?摇摇尾巴点点头 谁会跑呀,马会跑,马儿马儿怎样跑?四脚离地身不摇。 15、好朋友 你帮我来梳梳头,我帮你来扣纽扣 团结友爱手拉手,我们都是好朋友 嘿嘿! 16、数鸭子 门前大桥下,游过一群鸭, 快来快来数一数,二四六七八。 17、小雪花 小雪花,小雪花,飘在空中像朵花,小雪花,小雪花 飘在窗上变窗花,小雪花,小雪花,飘在手上不见了
童谣歌词大全 1慢地一说“斗虫飞”宝宝就会自己对拢食指. 6/// 摇煤球 摇啊摇''摇煤球''煤球大啊装不下'' 煤球小啊装不了。让宝宝躺在毯子上''爸爸妈妈对站''各拎着毯子的两个角''一边摇一边唱。 7/// 找朋友 (边唱边做动作) 找啊找啊找朋友 (摇晃头高兴的样子) 找到一个好朋友 (点头) 敬个礼呀 (把手伸到眼睛旁边) 握握手 (同人握手) 你是我的好朋友 (先指别人再指自己) 再见! (向人挥手) 8/// 动物叫 先让宝宝根据图片学认各种动物''如小猫''小狗''小鸡''小鸭''小羊等''等宝宝认识了各种动物后可以唱以下儿歌: 小猫怎么叫,喵喵喵小狗怎么叫,汪汪汪小鸡怎么叫,叽叽叽小鸭怎么叫,嘎嘎嘎 小羊怎么叫,咩咩咩老牛怎么叫,哞哞哞老虎怎么叫,噢噢噢青蛙怎么叫,呱呱呱 9/// 学自然界的各种声音 下雨了''哗哗哗;打雷了''轰隆隆;刮风了''呼呼呼;小河流水哗啦啦;汽车响''嘀嘀嘀;飞机飞''嗡嗡嗡;宝宝笑''哈哈哈;拍拍手''叭叭叭。 10/// 我有一个金娃娃。金鼻子金眼睛金头发。 第一天,我到河边去玩耍,丢了我的金娃娃。我哭我哭,我伤心地哭。 第二天,我到河边去洗衣,看见了我的金娃娃。我笑我笑,我哈哈地笑。 第三天,日本鬼子进了我的家,偷了我的鸡,抢了我的鸭,最后给我了三嘴巴。 第四天,红军叔叔来到我的家,还了我的鸡,还了我的鸭,最后给我了大红花。 第五天,阿姨叫我买苹果,我买了一箱烂苹果。阿姨打我,阿姨骂我,阿姨告诉爸爸, 爸爸告诉妈妈,妈妈告诉奶奶,奶奶告诉爷爷,爷爷告诉同学,同学告诉老师,老师告诉校长,校长打电话,说:“一不许动,二不许笑,三不许露出你的小白牙。本来要打你三百六十五下,马马虎虎打三下。 11/// 一二三,剁猪肝;四五六,揪肥肉;七八九,按电钮。 12/// 一根手指头,一根手指头,变成变成变成(小牛牛)毛毛虫二根手指头,二根手指头,变成变成变成(小剪刀)小白兔三根手指头,三根手指头,变成变成变成(印地安人)小花猫四根手指头,四根手指头,变成变成变成(小船船)螃蟹走五根手指头,五根手指头,变成变成变成(望远镜)小鸟飞六根手指头,六根手指头,变成变成变成(金元宝)打电话七根手指头,七根手指头,变成变成变成(长方形)三角形八根手指头,八根手指头,变成变成变成(神枪手)神枪手九根手指头,九根手指头,变成变成变成(小狗狗)小狗狗十根手指头,十根手指头,变成变成变成(拍拍手)拍拍手 13/// 一,一,一什么一,一是小猫穿花衣二,二,二什么二,二是小猫梳小辫三,三,三什么三,三是小猫爬雪山四,四,四什么四,四是小猫吃鱼刺五,五,五什么五,五是小猫打花鼓六,六,六什么六,六是小猫吃肥肉七,七,七什么七,七是小猫下象棋八,八,八什么八,八是小猫吹喇叭九,九,九什么九,九是小猫扭一扭
让孩子变成爱学习的天使 ——《第56号教室的奇迹》读书心得 杨地镇中心学校彭莉娜 非常感谢学校领导能给我这次去参加班主任培训的机会,让我有幸认识了被称为“美国最好的老师”、“传奇教师”雷夫.艾斯奎斯,人们还说“雷夫老师简直不是‘人’,而是‘神’的完美化身。”这次培训给了我很大的触动,最让我期待的是雷夫老师的《第56号教室的奇迹》。带着对雷夫老师的无比崇敬,我利用假期值班的时间,迫不及待地认真阅读了雷夫老师的《第56号教室的奇迹》。 一个由90%家庭贫困,且多出自非英语系的移民家庭的孩子,组成了一个不能再普通的班级,但他们在全国测试成绩高居全美前5%,他们长大后纷纷就读于哈佛、斯坦福等顶尖大学并取得了不凡成就。他们就是来自美国第二大小学——美国洛杉矶市霍巴特小学五年级中由雷夫?艾斯奎斯担任的一个班级。是什么力量创造了这样的奇迹?是什么魔法让孩子变成爱学习的天使?《第56号教室的奇迹》为我们解开了这个谜。一间教室能给孩子们带来什么,取决于教室桌椅之间的空白处流动着什么,相同面积的教室,有的显得很小,让人感到局促和狭隘,有的显得很大,让人觉得有无限伸展的可能,是什么东西在决定教室的尺度——教师,尤其是小学教师。他的面貌,决定了教室的内容;他的气质,决定了教室的容量。雷夫老师用自己都有独有的教学模式,给56号教室一个奇迹。雷夫·艾斯奎斯老师在教育生涯中的教育信念,以及独特的教育思想。同时他独特的个性、截然不同的教育观念、别出心裁的创意也使我心灵大为震撼。 体会一:“爱”的传递
雷夫对待学生,就如同对待他的孩子一样,付出了真爱。他凭着超凡的力量,在一个小时之内,改头换面地来回图书馆,目的就是为了给学生们借一本书!甚至他还坚持每天从早上6点开始,到下午5点一直都与学生共同学习,并完全无偿地利用寒暑假、周末等时间额外给学生补课!他还说“绝不拿某个学生的测验分数跟另一个学生相比,一定用学生自己过去的表现来衡量他现在的进步。” 雷夫说,这年头,大多数的教室都被一种东西控制着,那就是“害怕”。老师们害怕:怕丢脸,怕不受爱戴,怕说话没人听,怕场面失控。学生们更害怕:怕挨骂,怕在同学面前出丑,怕成绩不好,怕面对父母的盛怒。 对照我们的现实情况,完全如此。因为害怕,学习增加了许多的压力,因为害怕,教师感受不到与天真浪漫的孩子打交道时产生的乐趣。 然而,雷夫的第56号教室之所以特别,不是因为它拥有什么,而是因为它缺乏了一样东西——害怕。雷夫提醒我们,不管师教导学生还是子女,一定要时时从孩子的角度看事情,不要把害怕当作教育的捷径。 雷夫认为:只要改善班风,各种寻常的挑战就能迎刃而解。打造无恐惧的教室并非易事,可能的花上好多年时间,但这么做值得。为了在不靠恐惧手段的前提下让孩子们循规蹈矩,并使全班维持优异的学习表现,雷夫做了以下四件事: 一、以信任取代恐惧 在开学的第一天,在开始上课前的两分钟,雷夫与孩子们讨论了“信任”这个议题。在开学的第一天,雷夫和孩子们玩起了信任游戏,
100首童谣歌词大全 1、做早操 早上空气真叫好, 我们都来做早操。 伸伸臂,弯弯腰, 踢踢腿,蹦蹦跳 天天锻炼身体好。 2、饭前要洗手 小脸盆,水清请, 小朋友们笑盈盈, 小手儿,伸出来, 洗一洗,白又净, 吃饭前,先洗手, 讲卫生,不得病。 3、小手绢 小手绢,四方方, 天天带在我身上。 又擦鼻涕又擦汗, 干干净净真好看。 4、搬鸡蛋 小老鼠,搬鸡蛋, 鸡蛋太大怎么办? 一只老鼠地上躺, 紧紧抱住大鸡蛋。 一只老鼠拉尾巴, 拉呀拉呀拉回家。 5、大骆驼 骆驼骆驼志气大, 风吹日晒都不怕。 走沙漠,运盐巴, 再苦再累不讲话。 6、螳螂 螳螂哥,螳螂哥, 肚儿大,吃得多。 飞飞能把粉蝶捕, 跳跳能把蝗虫捉。 两把大刀舞起来, 一只害虫不放过 7、大蜻蜓 大蜻蜓,绿眼睛, 一对眼睛亮晶晶, 飞一飞,停一停, 飞来飞去捉蚊蝇。
8、小鸭子 小鸭子,一身黄,扁扁嘴巴红脚掌。嘎嘎嘎嘎高声唱,一摇一摆下池塘。 9、拍手歌 你拍一,我拍一,天天早起练身体。你拍二,我拍二,天天都要带手绢。你拍三,我拍三,洗澡以后换衬衫。你拍四,我拍四,消灭苍蝇和蚊子。你拍五,我拍五,有痰不要随地吐。你拍六,我拍六,瓜皮果核不乱丢。你拍七,我拍七,吃饭细嚼别着急。你拍八,我拍八,勤剪指甲常刷牙。你拍九,我拍九,吃饭以前要洗手。你拍十,我拍十,脏的东西不要吃。 10 、小螃蟹 小螃蟹,真骄傲,横着身子到处跑,吓跑鱼,撞倒虾,一点也不懂礼貌11 、庆六一 儿童节,是六一,小朋友们真欢喜。又唱歌来又跳舞,高高兴兴庆六一。 12、花猫照镜子小花猫,喵喵叫,不洗脸,把镜照,左边照,右边照,埋怨镜子脏, 气得胡子翘。13、蚂蚁搬虫虫小蚂蚁,搬虫虫,
童谣歌词大全 1/// 一只小蜜蜂呀,飞到花丛中呀,飞呀,飞呀。二只小耗子呀,跑到粮仓里呀,吃呀,吃呀。三只小花猫呀,去抓小耗子呀,追呀,追呀。四只小花狗呀,去找小花猫呀,玩呀,玩呀。五只小山羊呀,爬到山坡上呀,爬呀,爬呀。六只小鸭子呀,跳到水里面呀,游呀,游呀。七只小百灵呀,站在树枝上呀,唱呀,唱呀。八只小孔雀呀,穿上花衣裳呀,美呀,美呀。九只小白兔呀,竖起长耳朵呀,蹦呀,蹦呀。十个小朋友呀,一起手拉手呀,笑呀,乐呀。 2/// 1什么1,棍子1。2什么2,鸭子2。3什么3,耳朵3。4什么4,帆船4。5什么5,钩钩5。6什么6,大肚6。7什么7,拐杖7。8什么8,眼镜8。9什么9,汽球9。10什么10,棍子打棒球。 3/// 一根棍子轻轻打,二双筷子里外扒,三人小组爱说话,四个小兵不害怕,五个朋友力气大。 4/// 起床歌-小宝宝,起得早,睁开眼,眯眯笑,咿呀呀,学说话,伸伸手,要人抱穿衣歌-小胳膊,穿袖子,穿上衣,扣扣子,小脚丫,穿裤子,穿上袜子穿鞋子。小镜子-小镜子,圆又圆,看宝宝,露笑脸。闭上眼,做个梦,变月亮,挂上天。小铃铛-叮铃铃,叮铃铃,一会远,一会近。小宝宝,耳朵灵,听铃声,找到铃。学画画-小宝宝,学画画,大蜡笔,手中拿,画小鸭,叫嘎嘎,画小马,骑回家。大鞋子-大鞋子,象只船,爸爸穿,我也穿,一二一,象前走,走呀走,翻了船。逛公园-逛公园,宝宝笑,东看看,西瞧瞧,花儿香,鸟儿叫,小草绿,小树摇。看画报-小娃娃,看画报,睁大眼,仔细瞧,布娃娃,哈哈笑,伸伸手,要你抱。搭积木-大积木,红黄兰,小宝宝,最爱玩,搭火车,钻山洞,盖高楼,连着天。小汽车-小汽车,嘀嘀嘀,开过来,开过去,小宝宝,当司机,送妈妈,上班去。 5/// 斗虫飞 斗虫飞''斗虫飞''飞到南山吃露水'' 露水吃不到''回来吃青草。妈妈抱宝宝坐在膝上''用手拉起宝宝的两手''将他的两个食指对拢再分开''边唱这个歌谣.慢慢地一说“斗虫飞”宝宝就会自己对拢食指. 6/// 摇煤球 摇啊摇''摇煤球''煤球大啊装不下'' 煤球小啊装不了。让宝宝躺在毯子上''爸爸妈妈对站''各拎着毯子的两个角''一边摇一边唱。 7/// 找朋友(边唱边做动作) 找啊找啊找朋友(摇晃头高兴的样子) 找到一个好朋友(点头) 敬个礼呀(把手伸到眼睛旁边) 握握手(同人握手) 你是我的好朋友(先指别人再指自己) 再见! (向人挥手)
《一闪一闪亮晶晶》 一闪一闪亮晶晶。漫天都是小星星。挂在天空放光明。好像千万小眼睛。太阳慢慢向西沉。乌鸦回家一群群。星星眨着小眼睛。光辉照耀到天明。一闪一闪亮晶晶。满天都是小星星。 《春天在哪里》 春天在哪里呀。春天在哪里。春天在那青翠的山林里。这里有红花呀这里有绿草。还有那会唱歌的小黄鹂。嘀哩哩嘀哩嘀哩哩。嘀哩哩嘀哩嘀哩哩。春天在青翠的山林里。还有那会唱歌的小黄鹂。春天在哪里呀。春天在哪里。春天在那湖水的倒影里。映出红的花呀映出绿的草。还有那会唱歌的小黄鹂。 《卖报歌》 卖报歌。啦啦啦,啦啦啦,我是卖报的小行家,不等天明去等派报,一面走一面叫,今天的新闻真正好,七个铜板就买两份报。啦啦啦,啦啦啦,我是卖报的小行家,大风大雨里满街跑,走不好滑一跤,满身的泥水惹人笑,饥饿寒冷只有我知道。 《两只老虎》 两只老虎。两只老虎。跑的块跑的快。一只没有耳朵。一只没有尾巴。真奇怪。真奇怪。两只老虎。两只老虎。跑的块跑的快。一只没有鼻子。一只没有头发。真奇怪。真奇怪。 《葫芦娃》 葫芦娃葫芦娃。一根藤上七朵花。风吹雨打都不怕。啦啦啦啦。叮当咚咚当当。葫芦娃。叮当咚咚当当。本领大啦啦啦啦。葫芦娃葫芦娃。本领大。葫芦娃葫芦娃。一根藤上七朵花。风吹雨打都不怕。啦啦啦啦。叮当咚咚当当。葫芦娃。叮当咚咚当当。本领大啦啦啦啦。 《歌声与微笑》 请把我的歌带回你的家。请把你的微笑留下。请把我的歌带回你的家。请把你的微笑留下。明天明天这歌声飞遍海角天涯。飞遍海角天涯。明天明天这微笑将是遍野春花。将是遍野春花。 《小螺号》 小螺号嘀嘀嘀吹。海鸥听了展翅飞。小螺号嘀嘀嘀吹。浪花听了笑微微。小螺号嘀嘀嘀吹。声声唤船归啰。小螺号嘀嘀嘀吹。阿爸听了快快回啰。茫茫的海滩。蓝蓝的海水。吹起了螺号。心里美吔。 《小毛驴》 我有一只小毛驴我从来也不骑。有一天我心血来潮骑着去赶集。我手里拿着小皮鞭我心里正得意。不知怎么哗拉拉拉拉我摔了一身泥。我有一只小毛驴我从来也不骑。有一天我心血来潮骑着去赶集。我手里拿着小皮鞭我心里正得意。不知怎么哗拉拉拉拉我摔了一身泥。 《数鸭子》 门前大桥下游过一群鸭。快来快来数一数。二四六七八。嘎嘎嘎嘎。真呀真多呀。数不清到底多少鸭。数不清到底多少鸭。赶鸭老爷爷胡子白花花。唱呀唱着家乡戏。还会说笑话。小孩小孩。快快上学校。别考个鸭蛋抱回家。别考个鸭蛋抱回家。 《一只哈巴狗》 一只哈巴狗。站在大门口。眼睛黑油油。想吃肉骨头。一只哈巴狗。吃完肉骨头。尾巴摇一摇。向我点点头。
[ 百首儿歌童谣歌词大全 1、做早操 ?早上空气真叫好,我们都来做早操。 伸伸臂,弯弯腰,踢踢腿,蹦蹦跳,天天锻炼身体好。 2、饭前要洗手 小脸盆,水清请,小朋友们笑盈盈,小手儿,伸出来,洗一洗,白又净,吃饭前,先洗手,讲卫生,不得病。 3、小手绢 小手绢,四方方,天天带在我身上。 又擦鼻涕又擦汗,干干净净真好看。 4、搬鸡蛋 小老鼠,搬鸡蛋,鸡蛋太大怎么办?一只老鼠地上躺,紧紧抱住大鸡蛋。一只老鼠拉尾巴,拉呀拉呀拉回家。 5、大骆驼 骆驼骆驼志气大,风吹日晒都不怕。 走沙漠,运盐巴,再苦再累不讲话。 6、螳螂 螳螂哥,螳螂哥,肚儿大,吃得多。飞飞能把粉蝶捕,跳跳能把蝗虫捉。两把大刀舞起来,一只害虫不放过
7、大蜻蜓 大蜻蜓,绿眼睛,一对眼睛亮晶晶,飞一飞,停一停,飞来飞去捉蚊蝇。 8、小鸭子 小鸭子,一身黄,扁扁嘴巴红脚掌。嘎嘎嘎嘎高声唱,一摇一摆下池塘。 9、拍手歌 你拍一,我拍一,天天早起练身体。你拍二,我拍二,天天都要带手绢。你拍三,我拍三,洗澡以后换衬衫。你拍四,我拍四,消灭苍蝇和蚊子。你拍五,我拍五,有痰不要随地吐。你拍六,我拍六,瓜皮果核不乱丢。你拍七,我拍七,吃饭细嚼别着急。你拍八,我拍八,勤剪指甲常刷牙。你拍九,我拍九,吃饭以前要洗手。你拍十,我拍十,脏的东西不要吃。 10 、小螃蟹 小螃蟹,真骄傲,横着身子到处跑,吓跑鱼,撞倒虾,一点也不懂礼貌
11 、庆六一 儿童节,是六一,小朋友们真欢喜。 又唱歌来又跳舞,高高兴兴庆六一。 12、花猫照镜子 小花猫,喵喵叫,不洗脸,把镜照, 左边照,右边照,埋怨镜子脏,气得胡子翘。 13、蚂蚁搬虫虫 小蚂蚁,搬虫虫,一个搬,搬不动,两个搬,掀条缝,三个搬,动一动,四个五个六七个,大家一起搬进洞。 14、小青蛙 小青蛙,呱呱呱,水里游,岸上爬, 吃害虫,保庄稼,人人都要保护它。 15、花儿好看我不摘 公园里,花儿开,红的红,白的白, 花儿好看我不摘,人人都说我真乖。 16 、红绿灯 大马路,宽又宽,警察叔叔站中间, 红灯亮,停一停,绿灯亮,往前行。 17 、七个果果 一二三四五六七,七六五四三二一。
儿歌童谣大全拍手歌歌词: 你拍一,我拍一,一个小孩坐飞机,你拍二,我拍二,二个小孩骑马儿,你拍三,我拍三,三个小孩爬高山,你拍四,我拍四,四个小孩写大字,你拍五,我拍五,五个小孩在跳舞,你拍六,我拍六,六个小孩滚雪球,你拍七,我拍七,七个小孩坐滑梯,你拍八,我拍八,八个小孩吹喇叭,你拍九,我拍九,九个小孩玩气球,你拍十,我拍十,十个小孩来剪纸。 你拍一,我拍一,一个小孩坐飞机,你拍二,我拍二,二个小孩手腕,你拍三,我拍三,三个小孩去爬山,你拍四,我拍四,四个小孩写大字,你拍五,我拍五,五个小孩学跳舞,你拍六,我拍六,六个小孩载杨柳,你拍七,我拍七,七个小孩滑滑梯,你拍八,我拍八,八个小孩吹喇叭,你拍九,我拍九,九个小孩拉拉手,你拍十,我拍十,动手动脑长知识
你拍一,我拍一,一个宝宝开飞机, 你拍二,我拍二,二个宝宝梳小辫(注意读儿音), 你拍三,我拍三,三个宝宝去爬山, 你拍四,我拍四,四个宝宝学写字, 你拍五,我拍五,五个宝宝敲锣鼓, 你拍六,我拍六,六个宝宝吃石榴, 你拍七,我拍七,七个宝宝做游戏, 你拍八,我拍八,八个宝宝吹喇叭, 你拍九,我拍九,我们都是好朋友, 你拍十,我拍十,吃饭干净不挑食。 你拍一,我拍一,宝宝小手在哪里。 你拍二,我拍二,妈妈和你说句话。 你拍三,我拍三,宝宝身子翻一翻。 你拍四,我拍四,爸爸教你写大字。 你拍五,我拍五,宝宝肚子鼓一鼓。 你拍六,你拍六,宝宝体操一起做 你拍七,我拍七,宝宝小腿踢一踢。 你拍八,我拍八,宝宝学着爬一爬。 你拍九,我拍九,宝宝屁股扭一扭。 你拍十,我拍十,宝宝身体美又直 部门管理制度1. 规定 1.1
幼儿园儿歌歌词 1《花仙子之歌》 lu lu lu lu lu lu lu lu lu lu lu lu lu lu lu lu lu 能给人们带来幸福的花儿啊。你在哪里悄悄地开放。我到处把你找。脚下的路伸向远方。大波斯菊是我的帽子。蒲公英在我在我枕边飘荡。穿过那阴森的针槐林。奋勇向前向前。幸福的花仙子就是我。名字叫Lulu不寻常。说不定说不定有那么一天。就来到来到你身旁。lu lu lu lu lu。lu lu lu lu lu。lu lu lu lu lu lu lu。大波斯菊是我的帽子。蒲公英在我在我枕边飘荡。穿过那阴森的针槐林。奋勇向前向前。幸福的花仙子就是我。名字叫Lulu不寻常。说不定说不定有那么一天。就来到来到你身旁。lu lu lu lu lu。lu lu lu lu lu。lu lu lu lu lu lu lu。 2《哪咤传奇》 是他。是他。就是他。我们的朋友。小哪咤。就是他。是他。就是他。少年英雄。小哪咤上天他比天要高。下海他比海更大。智斗妖魔勇降鬼怪。少年英雄就是小哪咤。有时他也聪明有时他也犯傻。他的个头和我一般高。有时他很努力有时他也贪玩。他的年纪和我一般大。上天他比天要高。下海他比海更大。智斗妖魔勇降鬼怪。少年英雄就是小哪咤。 3《两个小娃娃打电话》 两个小娃娃呀正在打电话呀。喂喂喂你在哪里呀。诶诶诶我在幼儿园。两个小娃娃呀正在打电话呀。喂喂喂你在做什么。诶诶诶我在学唱歌。两个小娃娃呀正在打电话呀。喂喂喂你在哪里呀。诶诶诶我在幼儿园。两个小娃娃呀正在打电话呀。喂喂喂你在做什么。诶诶诶我在学唱歌。两个小娃娃呀正在打电话呀。喂喂喂你在哪里呀。诶诶诶我在幼儿园。两个小娃娃呀正在打电话呀。喂喂喂你在做什么。诶诶诶我在学唱歌。 4《山猫和吉米》 蓝蓝的天空,青青的草地 快乐的山猫,快乐的吉咪 来到了大自然,呼吸新空气 快乐成长,健康成长, 这里是快乐的,快乐的天地 来吧。。来吧来吧。。。来吧。。来吧来吧。。。来吧。。来到我的身边 来吧。。来吧来吧。。。来吧。。来吧来吧。。。和我一起唱歌跳舞 蓝蓝的天空,青青的草地 帅气的山猫,漂亮的吉咪 来到了我身边,快乐无极限快乐成长,健康成长 这里是快乐的,快乐的天地 5《大头儿子小头爸爸》 大头儿子,小头爸爸.。一对好朋友,快乐父子俩.。儿子的头大手儿小,。爸爸的头小手儿大.。大手牵小手,走路不怕滑.。走着走着走走走走,转眼儿子就长大.。 啦啦啦啦啦啦拉啦,转眼儿子就长大! 6《小羊儿》 小羊咩咩叫妈妈。母羊咩咩也叫他。跟着妈妈一道去。吃饱早回家。小羊咩咩叫妈妈。 母羊咩咩也叫他。跟着妈妈一道去。吃饱早回家。小羊咩咩叫妈妈。母羊咩咩也叫他。 跟着妈妈一道去。吃饱早回家。小羊咩咩叫妈妈。母羊咩咩也叫他。跟着妈妈一道去。 吃饱早回家。跟着妈妈一道去。吃饱早回家。啦......啦......
儿歌大全100首是搜集最受欢迎,简单好学易懂的儿歌100首,儿歌内容是反映儿童生活的乐趣,将知识,道理融为一体,歌词多采用比兴手法,唱起来朗朗上口,节奏轻快,明朗,深受小朋友和家长们的喜欢。 儿歌大全100首: 1.娃哈哈 2.小小少年 3.马兰花 4.丢手绢 5.哆来咪 6.小喇叭 7.英文字母歌 8.泥娃娃英文版 9.学习雷锋好榜样 10.泥娃娃 11.童年 12.找朋友 13.雪绒花英文版 14.健康歌 15.祝你生日快乐 16.幸福的手歌 17.鲁冰花 18.妈妈的吻 19.祝你圣诞快乐 20.闽南语童谣系列
21.小蜜蜂 22.摇篮曲 23.读书郎 24.聪明的一休 25.我的果汁分你一半 26.打电话儿歌 27.小草 28.蜗牛与黄鹂鸟 29.家庭称呼歌 30.快乐的节日 31.春天在哪里 32.葫芦娃 33.虫儿飞 34.生日歌 35.游子吟 36.夜静思 has My Little Dog Gone 38.鹅咏古诗 39.一分钱 40.金兔大拜年 41.新年好 42.响叮当(英文版)
43.少年司马光 44.小白兔乖乖 45.洗澡歌 46.布娃娃弹琴 47.一个师傅仨和尚 48.白雪公主 49.我的糖果在哪里 50.时尚新版圣诞歌 51.一代猪侠 52.我爱洗澡 53.三只小猫 54.做早操歌 个字母歌 56.摇篮曲 57.赶海的小姑娘 58.白龙马 59.小白兔乖乖 60.爸爸妈妈听我说 61.花仙子之歌 62.小老鼠上登台 63.捉泥鳅 64.数鸭子
65.卖报歌 66.学英文歌 67.小白兔过小桥 68.可爱的蓝精灵 69.小燕子 70.小毛驴 71.小红帽 72.拔萝卜 73.少年英雄小哪 74.上学歌 75.小海螺 76.世上只有妈妈好 77.新年好 78.三个和尚 79.两只老虎 80.小兔子乖乖 81.采蘑菇的小姑娘 82.十二生肖歌 83.吉祥三宝 84.我的好妈妈 85.机器猫
《第56号教室的奇迹》读后感精品范文(精 选3篇) 《第56号教室的奇迹》读后感精品范文(精选3篇) 当细细品完一本名著后,相信你一定有很多值得分享的收获,需要好好地就所收获的东西写一篇读后感了。为了让您不再为写读后感头疼,下面是WTT精心整理的《第56号教室的奇迹》读后感精品范文,希望对大家有所帮助。 《第56号教室的奇迹》读后感1 这次寒假学校专门安排的假期共读书目是《第56号教室的奇迹》。对于雷夫·艾斯奎斯的了解并不多,如果不是学校安排,我还真是不会去查阅有关的材料。还没有拜读过外国教师这么专门的教学心得,细读之后,我的内心翻腾起不小的“浪花”,从他坦率和刚正的叙述中获益良多。下面从几个方面简单谈一谈自己的感触。 在文章开始有这么一个案例:一个年轻教师班里出现了一位邋遢的孩子,书包整天都是乱糟糟、脏兮兮的。为了让他改掉这个毛病,年轻教师当着全班学生的面把这位学生的书包抖搂出来,并拿出相机拍照,扬言要在家长返校日公开,还说出了“谁那里有垃圾,就丢到这位同学的桌子上,反正他那里已经是垃圾桶了。”的话语。其结果是,家长十分的气愤,告到了校长处。
最可悲的是当事教师还没有认识到自己的错误,反而认为自己做的对,说现在这个孩子已经指导整理好自己的书包了。雷夫先生认为“不要把害怕当成教育的捷径”。 工作中,我也经常会遇到这样“不遵守秩序”的孩子,通常会软硬兼施、绞尽脑汁地去“对付”,正如雷夫老师所说“为达目的,不择手段”。而第56号教室,孩子很沉静,而且文明、有礼到一个难以置信的程度,这里就像是块绿洲,并不是因为它拥有什么,而是因为它缺乏了这样的东西——害怕。用信任取代恐惧,做孩子可以信赖的依靠,讲求纪律和公平,并且成为孩子的榜样。 与铁腕管理相反,雷夫老师提倡的是“没有害怕的教育”和彼此信任;与“小红花”奖励不同,他则反复强调知识本身就是最好的奖品……优异的教学质量,孩子个个谦逊有礼、诚实善良。这样的成就,追溯其根源则是雷夫反复强调的“道德培养的六阶段”理论:我不想惹麻烦——我想要奖赏——我想取悦某人——我要遵守规则——我能体贴别人——我有自己的行为准则并奉行不悖。近25年的教育实践,雷夫深信:着力孩子的品格培养激发孩子自身的高要求才是成就孩子一生的根本。 雷夫老师认为,老师可以带班级,但决定这个班级优秀或平庸的是班上的学生。他总结出了教育的“六个阶段”:第一阶段:我不想惹麻烦。这一阶段的思维是以恐惧为基础的,而我们要孩子们有良好行为表现的最终目的,是让他们相信这么做是对