蛋 白 质 纯 化 仪 使 用 说 明
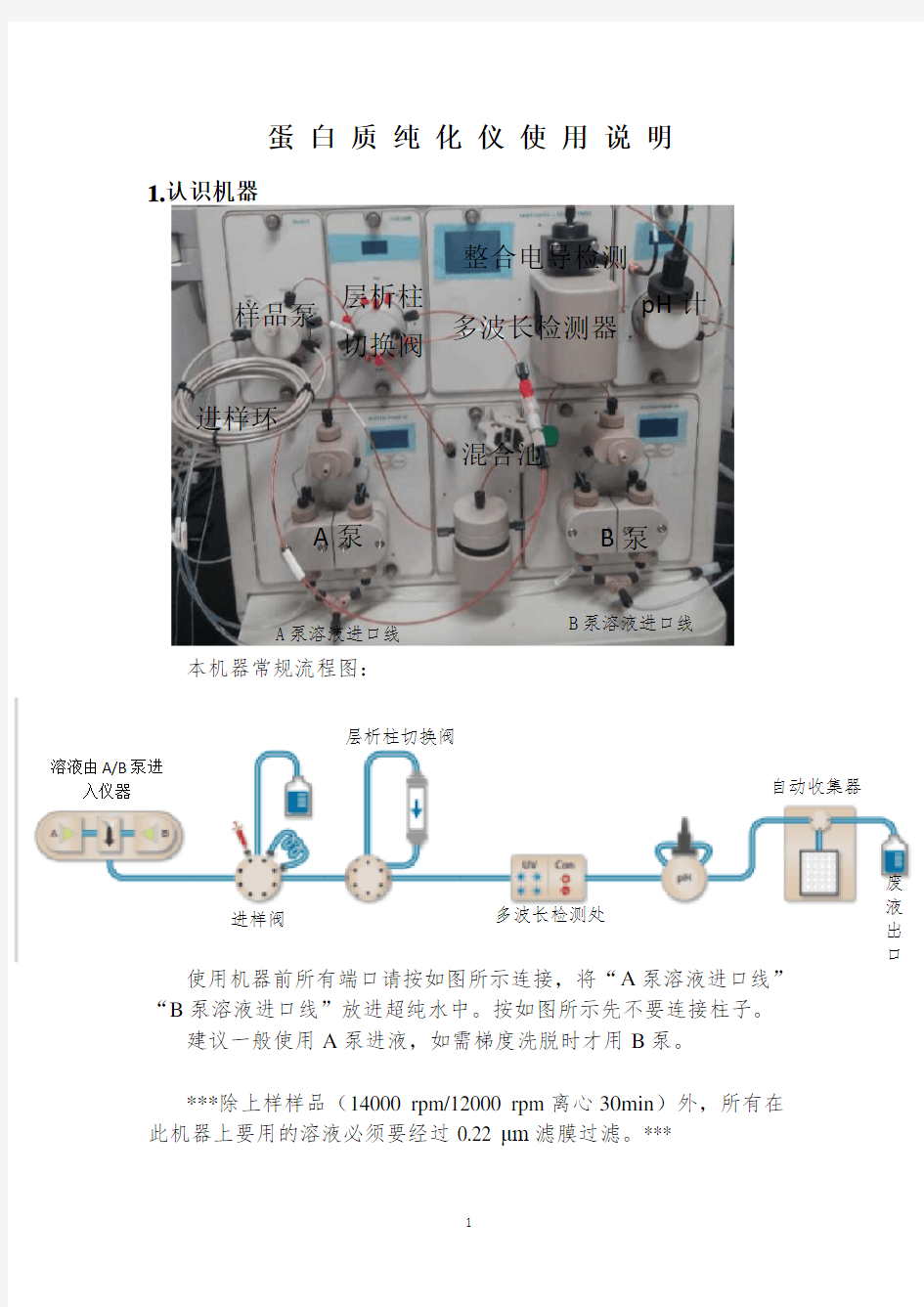
1.认识机器
本机器常规流程图:
使用机器前所有端口请按如图所示连接,将“A 泵溶液进口线”“B 泵溶液进口线”放进超纯水中。按如图所示先不要连接柱子。
建议一般使用A 泵进液,如需梯度洗脱时才用B 泵。
***除上样样品(14000 rpm/12000 rpm 离心30min )外,所有在此机器上要用的溶液必须要经过0.22 μm 滤膜过滤。***
样品泵 A 泵 B 泵
多波长检测器
pH 计
混合池
层析柱
切换阀 进样环 溶液由A/B 泵进
入仪器 进样阀
层析柱切换阀
多波长检测处 自动收集器 废 液 出 口
A 泵溶液进口线
B 泵溶液进口线
整合电导检测
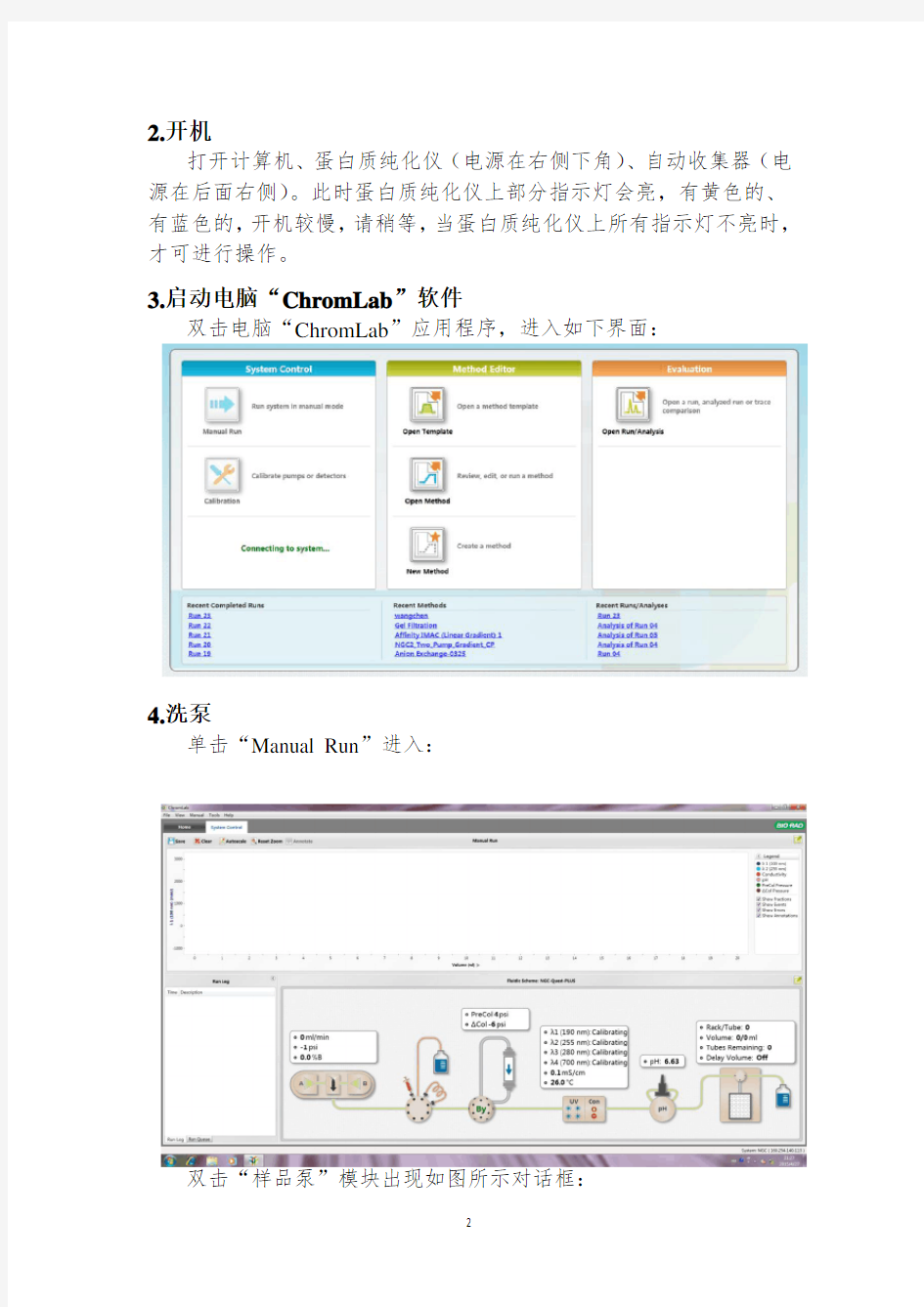
2.开机
打开计算机、蛋白质纯化仪(电源在右侧下角)、自动收集器(电源在后面右侧)。此时蛋白质纯化仪上部分指示灯会亮,有黄色的、有蓝色的,开机较慢,请稍等,当蛋白质纯化仪上所有指示灯不亮时,才可进行操作。
3.启动电脑“ChromLab”软件
双击电脑“ChromLab”应用程序,进入如下界面:
4.洗泵
单击“Manual Run”进入:
我们主要看右下方这块(图中所示模块均可双击设置参数):
双击“样品泵”模块出现如图所示对话框:
双击“层析柱切换阀”模块如下图所示:
此步骤主要是洗泵工作,请确保层析柱不在程序中,点击“Bypass ”
点击此处可让可更改溶液的流动方向。
点击1-5可设置连接不同的层析柱端口
设置流速
设置压力,小于选择的柱子承受最
大压力。在下面“控制流速以防止
超压”前面打对勾,以保护柱子。
不特殊设置,默认A 泵工作 此处可控制机器的运行与停止。
双击“样品泵”模块出现如下图所示:
选择“System Pump Waste”模式后,轻按仪器上“A泵”的“Purge”按钮,稍等片刻,即将进行A泵的洗泵工作,此时流速将会达到最大10 mL/min。如果想洗B泵(凝胶过滤时不需要),轻按仪器上“B 泵”的“Purge”按钮即可。注意此时废液将从“样品泵”模块上的
“Waste 2”出口处留出,拿好废液缸。
目的:洗A、B泵目的:Loop环上样
A、B泵洗完后,选择“Manual Load Loop”点击“Start”让超纯水在机身内流动片刻,此时废液将从自动收集器那边的废液口流出。点击“Stop”再选择“System Pump Inject Loop”此时再开始,将冲洗样品环。最后停止。
将A、B泵溶液进口线分别放入后续实验需要用的缓冲液中,重复以上过程。
嗯,洗完泵以后,把A泵溶液进口线放入超纯水中。
5.Developing a Method
单击“New Method ”进入如下界面:
首先,我们先选择合适模块运行程序,如1中的流程图所示是最佳选择:
如图所示是Ni-Affinity 1mL 柱体积的最佳模式:
选择运行模块按钮 选择柱子类型 选择柱子型号 柱子体积
柱子所承受最大压力 选择柱子连接端口处
自动收集器板块,目前实验室只能选择这个
控制程序默认流速处,一定要勾选第一个,防
止操作不当导致压力过大,对柱子造成损伤。第二个不必选中,本机器没有购买空气检测器。 此处勾选,pH 计才正常工作
设
置
波
长
检测
处 可设置按柱体积(CV )走溶液的用量,还是按ml 数走溶液用量
260
如图所示是Gel-filtration ENrichTM SEC 650的最佳模式:
当创建好上述内容后,我们就开始设置具体的步骤,一般来说一个纯化过程以下步骤是不可少的,如图:(此时可以点击左侧需要步骤按住不放,拖到右侧即可创建,添加洗脱步骤时一般选择“Colum Wash ”创建)
把鼠标放在右侧过程图中,单击右键可修改步骤名称。 下图流程仅供参考。
柱平衡
进样
洗脱样品 洗柱
Ni-Affinity 模式 Gel-filtration 模式:
可根据情况适当调整
下一步就是设置每一步的流速和溶液用量了(点击你所要设置的步骤,在右侧就会出现相应的设置条件):
例如,水洗、挂Ni 2+、柱平衡、不同浓度咪唑洗脱、EDTA 洗脱、20% 乙醇冲柱等过程设置如图:
20 % ethanol 水洗 20 % 乙醇
此处设置流速,打钩为默认流速,可不勾选,设置自需流速 单一溶液洗脱时,双击
此按钮,当B 溶液为
100%时,说明溶液全部从B 泵进入。为0时,
溶液均从A 泵进。 当设置梯度洗脱时双击此按钮。连续梯
度洗脱,不设B 溶液的百分比,只设置
洗脱溶液体积即可;阶段性梯度洗脱,
需设置B 溶液的百分比和洗脱体积。例图所示代表含义为:50%的A 溶液和50%的B 溶液混合后洗脱,洗脱体积为10个
柱体。 勾选此处可设置,自动收集。
全部收集
设置收集体积, 图例为0.2 mL 每管 单击此处,可设置按波峰收集。一般不建
议用。除非已对所纯化蛋白出峰位置非常
了解。
样品进样设置如下:
按如上过程设置好自己需要的流程后,即可开始程序: 点击上方第二行的“Star Run ”→“OK ”即可。
运行以后会出现如下图画面,等待机器运行:
设置进样环体积
1ml/5ml 首先单击这里
选择进样环进样 设置流速,建议不要太快,影响挂柱效率
设置进样环体积1ml/5ml
程序运行以后,先不要急着休息,点击下图左上角的“Hold Step”,此时连接柱子。
将“层析柱交换阀”上相互连接的两根线从中间拧开,接柱子。
注意:
1.注意柱身上标示的溶液流动方向(上端→下端)。
2.旋开柱子顶端和下端的螺母,让纯化仪的出水端先连接柱子的下端,让水从柱子的上端流出,待柱子上端有水滴停留时,将纯化仪的出水端接到柱子的上端,此时再连接柱子的下端。目的:防止有气泡进入柱子内。
3.如果用凝胶过滤柱时,除了要特别注意防止气泡进入,还要,注意的摆放姿势一定要保持垂直,否则会影响纯化的效率。
4.顺便说一下,柱子使用完毕后,应在保持液体流动的情况下,先拆卸柱子的下端,再拆卸柱子的上端,防止柱内保护液体的损失。4℃保存。
柱子安装完毕后,在“Hold Step”点击“Resume”开始运行你预先设置好的程序。
嗯,当然,咱们的机器也不是全自动的,开头和结尾还是需要动身换一下相应的溶液的。1.记好是A泵还是B泵进液。2.换溶液时,
注意将仪器暂停以后再操作,手不要抖,动作尽可能的慢一点。3.换溶液时,还要注意看看溶液进口处有没有空气进入,有的话,可以用注射器加点液体到细管内。
好了,那就可以安心的让它运行了。*记得柱平衡步骤时,调零。
此时如果无聊的话,你就可以将样品倒吸入样品环了,反正早晚也要弄。顺便把注射器、进样针、Loop环和自制的出液口细管(连接在样品泵“Waste 2”的出口上)再用缓冲液多洗两遍以防止污染。
*注意你的样品要提前在14000 rpm(最好,还可冷冻离心)/12000 rpm离心30 min,重新将上清转移到新的离心管中,以确保样品中不含微小颗粒,防止阻塞仪器和柱子。
*上样技巧:
1.确定你的样品量:以1 mL进样环为例,最好样品体积为800-900 μl最佳,此时倒吸一定不要人为引入气泡。
2.然后用200-100 μl的蛋白缓冲涮洗离心管。1 mL你懂得,千万不能超。
3.加样后,进样针就不可以离开进样孔了,否则样品将会从被吸上来的地方流出去。冲洗离心管时让它留在那就行了。
4.倒吸涮洗离心管的溶液时,可将溶液都吸完,还可以稍微多吸那么一点点,但也仅仅那么一点点,为了是不让样品浪费,你要保证空气不能进入进样环哦~
蛋白质纯化的方法 蛋白质的分离纯化方法很多,主要有: (一)根据蛋白质溶解度不同的分离方法 1、蛋白质的盐析 中性盐对蛋白质的溶解度有显著影响,一般在低盐浓度下随着盐浓度升高,蛋白质的溶解度增加,此称盐溶;当盐浓度继续升高时,蛋白质的溶解度不同程度下降并先后析出,这种现象称盐析,将大量盐加到蛋白质溶液中,高浓度的盐离子(如硫酸铵的SO4和NH4)有很强的水化力,可夺取蛋白质分子的水化层,使之“失水”,于是蛋白质胶粒凝结并沉淀析出。盐析时若溶液pH在蛋白质等电点则效果更好。由于各种蛋白质分子颗粒大小、亲水程度不同,故盐析所需的盐浓度也不一样,因此调节混合蛋白质溶液中的中性盐浓度可使各种蛋白质分段沉淀。 影响盐析的因素有:(1)温度:除对温度敏感的蛋白质在低温(4度)操作外,一般可在室温中进行。一般温度低蛋白质溶介度降低。但有的蛋白质(如血红蛋白、肌红蛋白、清蛋白)在较高的温度(25度)比0度时溶解度低,更容易盐析。(2)pH值:大多数蛋白质在等电点时在浓盐溶液中的溶介度最低。(3)蛋白质浓度:蛋白质浓度高时,欲分离的蛋白质常常夹杂着其他蛋白质地一起沉淀出来(共沉现象)。因此在盐析前血清要加等量生理盐水稀释,使蛋白质含量在2.5-3.0%。 蛋白质盐析常用的中性盐,主要有硫酸铵、硫酸镁、硫酸钠、氯化钠、磷酸钠等。其中应用最多的硫酸铵,它的优点是温度系数小而溶解度大(25度时饱和溶液为4.1M,即767克/升;0度时饱和溶解度为3.9M,即676克/升),在这一溶解度范围内,许多蛋白质和酶都可以盐析出来;另外硫酸铵分段盐析效果也比其他盐好,不易引起蛋白质变性。硫酸铵溶液的pH常在4.5-5.5之间,当用其他pH值进行盐析时,需用硫酸或氨水调节。 蛋白质在用盐析沉淀分离后,需要将蛋白质中的盐除去,常用的办法是透析,即把蛋白质溶液装入秀析袋内(常用的是玻璃纸),用缓冲液进行透析,并不断的更换缓冲液,因透析所需时间较长,所以最好在低温中进行。此外也可用葡萄糖凝胶G-25或G-50过柱的办法除盐,所用的时间就比较短。
组氨酸(His)标签蛋白的纯化 His-Tag融合蛋白是目前最常见的表达方式,而且很成熟,它的优点是表达方便而且基本不影响蛋白的活性,无论是表达的蛋白是可溶性的或者包涵体都可以用固定金属离子亲和色谱(IMAC)纯化。 IMAC(Immobilized Metal-ion affinity chromatography)是Porath et 年用固定IDA作为配基的填料螯合过渡金属铜、镍、钴或锌离子,可以吸附纯化表面带组氨酸、色氨酸或半胱氨酸残基的蛋白,1987年Smith et al. 发现带有几个组氨酸或色氨酸小肽和螯合金属离子的IDA-sephadex G-25作用力更强,此前在1986年他和他的合作者用Ni2+-IDA-sephadex G-25亲和纯化在氨基端带组氨酸和色氨酸的胰岛素原。同年1987年Hochuli et al.发现带有相连组氨酸的多肽和Ni2+-NTA填料作用力更强于普通的肽,1988年他第一次用这样的方法纯化了带六个组氨酸标签的多肽,无论是在天然还是变性条件下一次亲和纯化都得到很好效果,此后表达带六个组氨酸标签的蛋白配合IMAC变得非常普遍,相对而言,不带标签的蛋白纯化就非常困难,所以表达带六个组氨酸标签的蛋白配合IMAC 纯化变成最常用而且最有效的研究蛋白结构和功能的有力手段。1986年Porath et al.还发现Fe3+-IDA-sephadex G-25可以用于磷酸化蛋白的纯化,而后发现Ga3+-IDA也有同样的效果,这样螯合这两种金属离子的填料就有效用于磷酸化多肽的富集和纯化,同时IMAC也可以用于纯化各种和金属离子结合的多肽,应用非常广泛。 Ni柱中的氯化镍可以与有HIs(组蛋白)标签的蛋白结合,也可以与咪唑结合。 步骤是:过柱子前可以选择Ni柱重生,也就是往柱子里倒氯化镍,一个柱长体积就行了,然后平衡柱子,拿你自己的buffer,给蛋白提供最适的环境,我一般平衡4个柱长,然后蛋白上样,你可以让他自己挂,这样挂柱子的效果好一些,如果流速太慢,可以加个恒流泵,但是一定不能太快,太快挂柱效果差,当然你也可以选择循环挂柱,就是恒流泵的一头接你装蛋白的烧杯,从柱子中留下来的液体还用同一个烧杯接回去。挂完之后,按理想来讲,你的蛋白在Ni柱中与Ni就结合了,杂蛋白多数在烧杯里,留下来了,当然肯定有少量杂蛋白也挂上了,这时候你要,拿咪唑和你的buffer配,一般从0 20mM 40mM。。。。100mM 这样洗脱(当你不知道你的蛋白大概在什么时候出来的时候)我指的是咪唑的终浓度。咪唑加入之后,会和蛋白争夺与Ni的结合位点,杂蛋白、你的目的蛋白,会在不同的浓度被洗脱下来,洗完之后,你可以用400mM咪唑洗柱子,清理一切蛋白,然后平衡几次,是否选择重生你自己定咯~然后放上20%乙醇保存柱子就可以咯~过的蛋白用不同的管子收下,然后SDS-page检测在哪个管子里。 市面常见的商品化IMAC用于带六个组氨酸标签蛋白的配基有以下几种: 一、组氨酸(His)标签蛋白的纯化步骤: 大肠杆菌的破碎方法: 1)收集培养发酵液,4度7000-8000g离心10分钟,收集沉淀的菌体(如果不是马上破碎可以放-70度冷冻,但是最好能保存成小块或者薄片,这样好用。) 2)取1-2克菌体加10ml破碎缓冲液(的50mM磷酸缓冲液含NaCl,ml溶菌酶,1mM PMSF,1mM MgCl2,ml Benzonase,其中的菌酶,1mM PMSF,ml Benzonase现加)在冰上混合45分钟,如果pH不在7-8,需要用NaOH一边搅拌一边滴加.如果溶菌酶10mg/ml混合时间可以缩短到10分钟.
蛋白质纯化与结晶的原理 获得蛋白质的晶体结构的第一个瓶颈,就是制备大量纯化的蛋白质(>10 mg),其浓度通常在10 mg/ml 以上,并以此为基础进行结晶条件的筛选。运用重组基因的技术,将特定基因以选殖(clone)的方式嵌入表现载体(expression vector)内,此一载体通常具有易于调控的特性。之后再将带有特定基因的载体送入可快速生长的菌体中,如大肠杆菌(Escherichia coli),在菌体快速生长的同时,也大量生产表现载体上的基因所解译出之蛋白质。一般而言纯度越高的蛋白质比较有机会形成晶体,因此纯化蛋白质的步骤就成为一个重要的决定因素。 在取得高纯度的蛋白质溶液后,接下来就是晶体的培养。蛋白质晶体与其他化合物晶体的形成类似,是在饱和溶液中慢慢产生的,每一种蛋白质养晶的条件皆有所差异,影响晶体形成的变量很多,包含化学上的变量,如酸碱度、沈淀剂种类、离子浓度、蛋白质浓度等;物理上的变数,如溶液达成过饱和状态的速率、温度等;及生化上的变数,如蛋白质所需的金属离子或抑制剂、蛋白质的聚合状态、等电点等,皆是养晶时的测试条件。截至目前为止,并无一套理论可以预测结晶的条件,所以必须不断测试各种养晶溶液的组合后,才可能得到一颗完美的单一晶体(图一) 。 蛋白质晶体的培养,通常是利用气相扩散法(Vapor Diffusion Method) 的原理来达成;也就是将含有高浓度的蛋白质(10-50 mg/ml)溶液加入适当的溶剂,慢慢降低蛋白质的溶解度,使其接近自发性的沈淀状态时,蛋白质分子将在整齐的堆栈下形成晶体。举例来说,我们将蛋白质溶于低浓度(~1.0 M) 的硫酸铵溶液中,将它放置于一密闭含有高浓度(~2.0 M)硫酸铵溶液的容器中,由气相平衡,可以缓慢提高蛋白质溶液中硫酸铵的浓度,进而达成结晶的目的(图二)。 蛋白质晶体在外观上与其他晶体并无明显不同之处,但在晶体的内部,却有很大的差异。一般而言,蛋白质晶体除了蛋白质分子外,其他的空间则充满约40 %至60 %之间的水溶液,其液态的成分不仅使晶体易碎,也容易使蛋白质分子在晶格排列上有不规则的情形出现,造成晶体处理时的困难及绕射数据上的搜集不易等缺点。但也由于高含水量的特性,让蛋白质分子在晶体内与水溶液中的状态,极为相似。所以由晶体所解出的蛋白质结构,基本上可视为自然状态下的结构。 蛋白质结构解析的方法简介 到目前为止,蛋白质结构解析的方法主要是两种,x射线衍射和NMR。近年来还出现了一种新的方法,叫做Electron Microscopy。其中X射线的方法产生的更早,也更加的成熟,解析的数量也更多,我们知道,第一个解析的蛋白的结构,就是用x晶体衍射的方法解析的。而NMR方法则是在90年代才成熟并发展起来的。这两种方法各有优点和缺点。 首先来说一下,这两种方法的一般的步骤和各自的优点和缺点。电子显微镜(electron microsco py)作为一种新型的技术,目前的应用还是非常少,并且比较狭窄,我可能等到最后在给它作些
蛋白质的提取和纯化-- 选择分离材料及预处理蛋白质的提取和纯化-- 选择分离材料及预处理 以蛋白质和结构与功能为基础,从分子水平上认识生命现象,已经成为现代生物学发展的主要方向,研究蛋白质,首先要得到高度纯化并具有生物活性的目的物质。 蛋白质的制备工作涉及物理、化学和生物等各方面知识,但基本原理不外乎两方面。一是得用混合物中几个组分分配率的差别,把它们分配到可用机械方法分离的两个或几个物相中,如盐析,有机溶剂提取,层析和结晶等;二是将混合物置于单一物相中,通过物理力场的作用使各组分分配于来同区域而达到分离目的,如电泳,超速离心,超滤等。在所有这些方法的应用中必须注意保存生物大分子的完整性,防止酸、硷、高温,剧烈机械作用而导致所提物质生物活性的丧失。 蛋白质的制备一般分为以下四个阶段:选择材料和预处理,细胞的破碎及细胞器的分离,提取和纯化,浓细、干燥和保存。 微生物、植物和动物都可做为制备蛋白质的原材料,所选用的材料主要依据实验目的来确定。对于微生物,应注意它的生长期,在微生物的对数生长期,酶和核酸的含量较高,可以获得高产量,以微生物为材料时有两种情况:( 1 )得用微生物菌体分泌到培养基中的代谢产物和胞外酶等;(2)利用菌体 含有的生化物质,如蛋白质、核酸和胞内酶等。植物材料必须经过去壳,脱脂并注意植物品种和生长发育状况不同,其中所含生物大分子的量变化很大,另外与季节性关系密切。对动物组织,必须选择有效成份含量丰富的脏器组织为原材料,先进行绞碎、脱脂等处理。另外,对预处理好的材料,若不立即进行实验,应冷冻保存,对于易分解的生物大分子应选用新鲜材料制备。 细胞的破碎 1、高速组织捣碎:将材料配成稀糊状液,放置于筒内约1/3 体积,盖紧筒盖,将调速器先拨至最慢处, 开动开关后,逐步加速至所需速度。此法适用于动物内脏组织、植物肉质种子等。 2、玻璃匀浆器匀浆:先将剪碎的组织置于管中,再套入研杆来回研磨,上下移动,即可将细胞研碎,此法细胞破碎程度比高速组织捣碎机为高,适用于量少和动物脏器组织。 3、超声波处理法:用一定功率的超声波处理细胞悬液,使细胞急剧震荡破裂,此法多适用于微生物材料, 用大肠杆菌制备各种酶,常选用50-100 毫克菌体/毫升浓度,在1KG 至10KG 频率下处理10-15 分钟,此法的缺点是在处理过程会产生大量的热,应采取相应降温措施。对超声波敏感和核酸应慎用。 4、反复冻融法:将细胞在-20 度以下冰冻,室温融解,反复几次,由于细胞内冰粒形成和剩余细胞液的盐浓度增高引起溶胀,使细胞结构破碎。 5、化学处理法:有些动物细胞,例如肿瘤细胞可采用十二烷基磺酸钠(SDS)、去氧胆酸钠等细胞膜破 坏,细菌细胞壁较厚,可采用溶菌酶处理效果更好。 无论用哪一种方法破碎组织细胞,都会使细胞内蛋白质或核酸水解酶释放到溶液中,使大分子生物 降解,导致天然物质量的减少,加入二异丙基氟磷酸(DFP)可以抑制或减慢自溶作用;加入碘乙酸可以
中国试剂网 3.15.2.19 蛋白质纯化与结晶的原理 获得蛋白质的晶体结构的第一个瓶颈,就是制备大量纯化的蛋白质(>10mg),其浓度通常在10mg/ml以上,并以此为基础进行结晶条件的筛选。运用重组基因的技术,将特定基因以选殖(clone)的方式嵌入表现载体(expression vector)内,此一载体通常具有易于调控的特性。之后再将带有特定基因的载体送入可快速生长的菌体中,如大肠杆菌(Escherichia coli),在菌体快速生长的同时,也大量生产表现载体上的基因所解译出之蛋白质。一般而言纯度越高的蛋白质比较有机会形成晶体,因此纯化蛋白质的步骤就成为一个重要的决定因素。 在取得高纯度的蛋白质溶液后,接下来就是晶体的培养。蛋白质晶体与其他化合物晶体的形成类似,是在饱和溶液中慢慢产生的,每一种蛋白质养晶的条件皆有所差异,影响晶体形成的变量很多,包含化学上的变量,如酸碱度、沉淀剂种类、离子浓度、蛋白质浓度等;物理上的变数,如溶液达成过饱和状态的速率、温度等;及生化上的变数,如蛋白质所需的金属离子或抑制剂、蛋白质的聚合状态、等电点等,皆是养晶时的测试条件。截至目前为止,并无一套理论可以预测结晶的条件,所以必须不断测试各种养晶溶液的组合后,才可能得到一颗完美的单一晶体。 蛋白质晶体的培养,通常是利用气相扩散法(Vapor Diffusion Method)的原理来达成;也就是将含有高浓度的蛋白质(10~50mg/ml)溶液加入适当的溶剂,慢慢降低蛋白质的溶解度,使其接近自发性的沈淀状态时,蛋白质分子将在整齐的堆栈下形成晶体。举例来说,我们将蛋白质溶于低浓度(~1.0M)的硫酸铵溶液中,将它放置于一密闭含有高浓度(~2.0M)硫酸铵溶液的容器中,由气相平衡,可以缓慢提高蛋白质溶液中硫酸铵的浓度,进而达成结晶的目的。 蛋白质晶体在外观上与其他晶体并无明显不同之处,但在晶体的内部,却有很大的差异。一般而言,蛋白质晶体除了蛋白质分子外,其他的空间则充满约40%至60%之间的水溶液,其液态的成分不仅使晶体易碎,也容易使蛋白质分子在晶格排列上有不规则的情形出现,造成晶体处理时的困难及绕射数据上的搜集不易等缺点。但也由于高含水量的特性,让蛋白质分子在晶体内与水溶液中的状态,极为相似。所以由晶体所解出的蛋白质结构,基本上可视为自然状态下的结构。
(二)利用溶解度差别 影响蛋白质溶解度的外部因素有:1、溶液的pH;2、离子强度;3、介电常数;4、温度。但在同一的特定外部条件下,不同蛋白质具有不同的溶解度。 1、等电点沉淀:原理:蛋白质处于等电点时,其净电荷为零,由于相邻蛋白质分子之间没有静电斥力而趋于聚集沉淀。因此在其他条件相同时,他的溶解度达到最低点。在等电点之上或者之下时,蛋白质分子携带同种符号的净电荷而互相排斥,阻止了单个分子聚集成沉淀,因此溶解度较大。不同蛋白质具有不同的等电点,利用蛋白质在等电点时的溶解度最低的原理,可以把蛋白质混合物分开。当pH被调到蛋白质混合物中其中一种蛋白质的等电点时,这种蛋白质大部分和全部被沉淀下来,那些等电点高于或低于该pH的蛋白质则仍留在溶液中。这样沉淀出来的蛋白质保持着天然的构象,能重新溶解于适当的pH和一定浓度的盐溶液中。 5、盐析与盐溶:原理:低浓度时,中性盐可以增加蛋白质溶解度这种现象称为盐溶.盐溶作用主要是由于蛋白质分子吸附某种盐类离子后,带电层使蛋白质分子彼此排斥,而蛋白质与水分子之间的相互作用却加强,因而溶解度增高。球蛋白溶液在透析过程中往往沉淀析出,这就是因为透析除去了盐类离子,使蛋白质分子之间的相互吸引增加,引起蛋白质分子的凝集并沉淀。当溶液的离子强度增加到一定程度时,蛋白质溶解程度开始下降。当离子强度增加到足够高时,例如饱和或半饱和程度,很多蛋白质可以从水中沉淀出来,这种现象称为盐析。盐析作用主要是由于大量中性盐的加入使水的活度降低,原来溶液中的大部分甚至全部的自由水转变为盐离子的水化水。此时那些被迫与蛋白质表面的疏水集团接触并掩盖他们的水分子成为下一步最自由的可利用的水分子,因此被移去以溶剂化盐离子,留下暴露出来的疏水基团。蛋白质疏水表面进一步暴露,由于疏水作用蛋白质聚集而沉淀。 盐析沉淀的蛋白质保持着他的天然构象,能再溶解。盐析的中性盐以硫酸铵为最佳,在水中的溶解度很高,而溶解度的温度系数较低。 3、有机溶剂分级分离法:与水互溶的有机溶剂(甲醇、乙醇和丙酮等)能使蛋白质在水中的溶解度显著降低。在室温下有机溶剂会引起蛋白质变性,如果预先将有机溶剂冷却到-40°C以下,然后在不断搅拌下逐滴加入有机溶剂,以防局部浓度过高,那么变性可以得到很大程度缓解。蛋白质在有机溶剂中的溶解度也随温度、pH和离子强度而变化。在一定温度、pH和离子强度条件下,引起蛋白质沉淀的有机溶剂的浓度不同,因此控制有机溶剂浓度也可以分
G S T标签蛋白纯化原理 SANY GROUP system office room 【SANYUA16H-
在实验中有过被高等教育的教授骂过:别人让你吃屎,你就去吃屎么?或者稍微高级点的问候声:你有脑子么? 请稍微忽视当事人的失落、自尊心碎一地的感受,苦难不是一种财富,对苦难的反思才是一种财富。在实验时,或许着急按照步骤一步一步的做下来,而没有肯定的问自己:为什么要这样做?为什么要加这样的试剂?所以忽视了一些看似无关紧要的细节,坏结果便发生了。 回归原理部分,回归解决若干个为什么的问题,让我们的实验更加规范化。所以在接下来若干个专题阐述GST 标签蛋白纯化的实验时,生物日记实验室部落首先搞清楚GST 蛋白纯化的原理。 ——当别人问为什么时,你知道为什么么 GST 标签蛋白纯化原理 重组标签蛋白纯化利用谷胱甘肽的结构能够和GST (谷胱甘肽S-转移酶)的结合位点互补,谷胱甘肽通过SH 基团与琼脂糖介质上的环氧乙烷基团通过环氧激活特异性偶联在琼脂糖介质上,带有GST 标签蛋白纯化原理 Justwhywhywhy
GST的标签蛋白与琼脂糖介质上交联的谷胱甘肽配体互补性结合,这种可逆性的结合在温和、非变性的条件下通过加入还原型谷胱甘肽洗脱下来,杂质通过结合缓冲液被洗脱去除,从而分离目的蛋白。如果需要将GST标签从目的蛋白上除去,则可将蛋白酶结合到柱子上,在标签蛋白与谷胱甘肽结合时使用蛋白酶位点特异性切割,也可在洗脱之后再酶切。另外,蛋白质的性质、载体的选择、宿主菌、表达和纯化的条件不同都能使最终标签蛋白的产量有很大的差别。 目的基因的表达在选择合适的表达载体时,应该注意载体的克隆位点、蛋白酶切位点、标签的位置、编码框等多种因素。 载体的选择 pGEX载体可用于构建可诱导、高水平表达目的基因片段,带有GST标签蛋白通过特定的目的基因或者基因片段插入到pGEX的多克隆位点,在pGEX载体上带有laclp基因,该基因表达产物作为抑制剂结合在tac启动子的操纵子区,而在乳糖类似物IPTG的诱导下,目的基因能够在tac启动子的调控下表达目的蛋白。 宿主菌的选择 在选择宿主菌时,大部分大肠杆菌都能够克隆和表达pGEX载体,如果要获得全长的标签蛋白,一些特殊的工程菌即蛋白酶Lon、OmpT等缺失的菌种能够保护目的蛋白不被宿主菌降解,获得全长的标签蛋白,并且产量也可能更高。
名词解释 1. 截留分子量(molecular weight cut-off, MWCO):不能通过膜的最小分子量 2. 超滤:选择合适孔径的超滤膜,在离心力或较高压力下,使水分子和其他小分子物质通过超滤膜,而目标蛋白样品分子被截留不能通过超滤膜,从而增加蛋白样品浓度,达到浓缩效果的方法 3、陶南效应:离子交换剂表面pH与溶液pH是不一致的。在阳IE表面的微环境中,H+被吸引而OH-被排斥,交换剂表面pH比周围低1个pH单位;而阴IE表面的微环境中,H+被排斥,交换剂表面pH比周围高1个pH单位 4、离子交换剂的有效(实际)交换容量:指在一定的实验条件下,每克干介质或每毫升湿胶吸附蛋白质的实际容量 5. 聚沉(coagulation)是指在聚沉剂的作用下,溶液中的蛋白质相互聚集为较大在聚沉物(>1mm)的过程 ?常见的聚沉剂:无机盐类(如氯化锌、氯化铁、氯化铝、硫酸锌、硫酸铝),聚合无机盐(聚合铝、聚合铁等) ?聚沉条件:-20℃以下,pH3~6,较高离子强度,高多价金属离子(Fe3+、Al3+) 6. 絮凝(flocculation)是指在絮凝剂的作用下,通过吸附、交联、网捕,把蛋白质聚结为大絮体沉降的过程 ?常见的絮凝剂:淀粉、树脂、单宁、离子交换树脂及纤维素衍生物 ?絮凝作用一般在聚沉作用之后使用;絮凝剂的选择应根据成本、毒性等具体情况考虑;应通过试验筛选获得最适合的絮凝剂类型、用量及作用条件 7、盐溶:蛋白质在稀盐溶液中,溶解度会随着盐浓度的增高而上升 8、盐析:但当盐浓度增高到一定数值时,其溶解度又逐渐下降,直至蛋白质析出 9、透析:利用小分子能通过,而大分子不能透过半透膜的原理,把不同性质的物质彼此分开的一种方法。对于蛋白质样品,透析过程中因蛋白质分子体积很大,不能透过半透膜,而溶液中的无机盐小分子则能透过半透膜进入水中,因此不断更换透析用水即可将蛋白质与无机盐小分子物质完全分开。蛋白的分离纯化过程中常用此法脱去无机盐(如硫酸铵) 10、排阻极限:指不能进入凝胶颗粒内部网孔的最小蛋白的分子量 11、凝胶颗粒的分级范围:指能进入凝胶颗粒内部网孔的最大分子和最小分子的分子量范围 12、过滤:利用多孔介质(滤纸、滤膜等)阻截大的颗粒物质,而使小于孔隙的物质通过的一种的分离方法。主要用于悬浮液的分离,最简单、最常用 13、离子交换剂的电荷密度:指IE介质颗粒单位表面积的功能基团数量,它决定着离子交换剂的总交换容量 14、离子交换剂膨胀度(吸水值):指干态的离子交换剂在水溶液中吸水后造成的体积膨胀程度。用每克干离子交换剂吸水膨胀后的体积表示(ml/g) 15、梯度洗脱:进行梯度洗脱时,洗脱缓冲液的pH或离子强度是连续发生变化的,洗脱剂的洗脱能力也是连续增加的 16、阶段洗脱:指在一个时间段内用一固定pH或I的条件进行洗脱,而在下一个时间段内用另一固定pH或I的条件进行洗脱的分段式洗脱方式 也分为pH阶段洗脱和I阶段洗脱 17、亲和层析:利用蛋白质与其专一性配体之间的特异性生物学亲和力作用,对蛋白质进行分离纯化的层析技术。 18、等电聚焦电泳:利用不同蛋白质的等电点的不同而使其在pH梯度中相互分离的一
分离纯化蛋白质的方法及原理 (一)利用分子大小 1、透析:原理:利用蛋白质分子不能透过半透膜的性质,使蛋白质和其他小分子物质如无机盐、单糖、水等分开。 方法:将待提纯蛋白质放在透析袋中放在蒸馏水中进行 涉及的问题: 如何加快透析过程 (1)加大浓度差,及时更换透析液 (2)利用磁力搅拌器 常用的半透膜:玻璃纸、火棉和其他材料合成 2、超过滤:原理:利用压力和离心力,强行使其他小分子和水通过半透膜,而蛋白质留在膜上 3、凝胶过滤层析:原理:当不同分子大小的蛋白质混合物流进凝胶层析柱时,比凝胶网孔大的分子不能进入珠内网状结构,排阻在凝胶珠以外,在凝胶珠缝隙间隙中向下移动。而比孔小的分子不同程度地进入凝胶珠内,这样由于不同大小分子所经历的路径不同而到分离。 结果:大分子先被洗脱下来,小分子后被洗脱下来 (二)利用溶解度差别 4、等电点沉淀:原理:不同蛋白质具有不同的等电点,当蛋白质混合物调到其中一种蛋白质的等电点时,这种蛋白质大部分和全部被沉淀下来.。 5、盐析与盐溶:原理:低浓度时,中性盐可以增加蛋白质溶解度这种现象称为盐溶.当离子强度增加,足够高时,例如饱和或半饱和程度,很多蛋白质可以从水中沉淀出来,这种现象称为盐析
(三)根据电荷不同 6、SDS-PAGE 全称十二烷基硫酸钠—聚丙烯酰胺凝胶电泳 原理:通过加热和SDS可以使蛋白质变性,多亚基的蛋白质也解离为单亚基,处理后的样品中肽链是处于无二硫键连接的,分离的状态。电泳时SDS-蛋白质复合物在凝胶中的迁移率不再受蛋白质原有电荷和形状的影响,而主要取决于蛋白质分子量。所以SDS-PAGE常用来分析蛋白质的纯度和大致测定蛋白质的分子量。 7、离子交换层析:原理:氨基酸分离常用阳离子交换树脂,树脂被处理成钠型,将混合氨基酸上柱,氨基酸主要以阳离子形式存在,在树脂上与钠离子发生交换,而被挂在树脂上。 氨基酸在树脂上结合的牢固程度取决于氨基酸与树脂之间的亲和力,决定亲和力的因素有:(1)主要是静电吸引力(2)氨基酸侧链同树脂之间的疏水作用氨基酸与阳离子交换树脂间的静电引力大小次序依次是: 碱性氨基酸R2+>中性氨基酸R+>酸性氨基酸R0。 因此洗脱顺序应该是: 酸性氨基酸中性氨基酸碱性氨基酸 为使氨基酸从树脂上洗脱下来采用逐步提高pH和盐浓度的方法
蛋白过镍柱纯化的原理和步骤是什么 ** Ni柱中的氯化镍可以与有HIs(组蛋白)标签的蛋白结合,也可以与咪唑结合。 步骤是:过柱子前可以选择Ni柱重生,也就是往柱子里倒氯化镍,一个柱长体积就行了,然后平衡柱子,拿你自己的buffer,给蛋白提供最适的环境。我一般平衡4个柱长,然后蛋白上样,你可以让他自己挂,这样挂柱子的效果好一些,如果流速太慢,可以加个恒流泵,但是一定不能太快,太快挂柱效果差,当然你也可以选择循环挂柱,就是恒流泵的一头接你装蛋白的烧杯,从柱子中留下来的液体还用同一个烧杯接回去。 挂完之后,按理想来讲,你的蛋白在Ni柱中与Ni就结合了,杂蛋白多数在烧杯里,留下来了,当然肯定有少量杂蛋白也挂上了。这时候你要梯度洗脱,拿咪唑和你的buffer配,一般从0、20mM、40mM......100mM这样洗脱(当你不知道你的蛋白大概在什么时候出来的时候)我指的是咪唑的终浓度。咪唑加入之后,会和蛋白争夺与Ni的结合位点,杂蛋白、你的目的蛋白,会在不同的浓度被洗脱下来,洗完之后,你可以用200mM咪唑洗柱子,清理一切蛋白,然后平衡几次(怎么平衡?),是否选择重生你自己定咯~然后放上20%乙醇保存柱子就可以咯~ 过的蛋白用不同的管子收下,然后SDS-page检测在哪个管子里。
**楼上说的基本对,但基本上里面填料时硫酸镍,因此柱子可以和碱性蛋白结合,组蛋白标签一般是6个组氨酸(碱性氨基酸)。在蛋白上样后,带有组氨酸标签的蛋白特异性结合到柱子里,其他的杂蛋白流出。这时候再用咪唑洗脱,梯度洗脱,咪唑竞争性结合到硫酸镍上,目的蛋白就被洗脱了,这时候收集浸出液,那么里面就是你要的目的蛋白,然后透析掉咪唑就行了。现在的柱子大多是预装柱,也就是不用自己填柱,就是装好镍的傻瓜柱,也不贵,上样,洗脱,基本两个步骤就结束了。剩下的看你怎么处理了。可以参考楼上的。不会再问。
蛋白质的纯化原理 一)根据蛋白质溶解度不同的分离方法 1、蛋白质的盐析 中性盐对蛋白质的溶解度有显著影响,一般在低盐浓度下随着盐浓度升高,蛋白质的溶解度增加,此称盐溶;当盐浓度继续升高时,蛋白质的溶解度不同程度下降并先后析出,这种现象称盐析,将大量盐加到蛋白质溶液中,高浓度的盐离子(如硫酸铵的SO4和NH4)有很强的水化力,可夺取蛋白质分子的水化层,使之“失水”,于是蛋白质胶粒凝结并沉淀析出。盐析时若溶液pH在蛋白质等电点则效果更好。由于各种蛋白质分子颗粒大小、亲水程度不同,故盐析所需的盐浓度也不一样,因此调节混合蛋白质溶液中的中性盐浓度可使各种蛋白质分段沉淀。 影响盐析的因素有:(1)温度:除对温度敏感的蛋白质在低温(4度)操作外,一般可在室温中进行。一般温度低蛋白质溶介度降低。但有的蛋白质(如血红蛋白、肌红蛋白、清蛋白)在较高的温度(25度)比0度时溶解度低,更容易盐析。(2)pH值:大多数蛋白质在等电点时在浓盐溶液中的溶介度最低。(3)蛋白质浓度:蛋白质浓度高时,欲分离的蛋白质常常夹杂着其他蛋白质地一起沉淀出来(共沉现象)。因此在盐析前血清要加等量生理盐水稀释,使蛋白质含量在2.5-3.0%。 蛋白质盐析常用的中性盐,主要有硫酸铵、硫酸镁、硫酸钠、氯化钠、磷酸钠等。其中应用最多的硫酸铵,它的优点是温度系数小而溶解度大(25度时饱和溶液为4.1M,即767克/升;0度时饱和溶解度为3.9M,即676克/升),在这一溶解度范围内,许多蛋白质和酶都可以盐析出来;另外硫酸铵分段盐析效果也比其他盐好,不易引起蛋白质变性。硫酸铵溶液的pH常在4.5-5.5之间,当用其他pH值进行盐析时,需用硫酸或氨水调节。 蛋白质在用盐析沉淀分离后,需要将蛋白质中的盐除去,常用的办法是透析,即把蛋白质溶液装入秀析袋内(常用的是玻璃纸),用缓冲液进行透析,并不断的更换缓冲液,因透析所需时间较长,所以最好在低温中进行。此外也可用葡萄糖凝胶G-25或G-50过柱的办法除盐,所用的时间就比较短。 2、等电点沉淀法 蛋白质在静电状态时颗粒之间的静电斥力最小,因而溶解度也最小,各种蛋白质的等电点有差别,可利用调节溶液的pH达到某一蛋白质的等电点使之沉淀,但此法很少单独使用,可与盐析法结合用。 3、低温有机溶剂沉淀法 用与水可混溶的有机溶剂,甲醇,乙醇或丙酮,可使多数蛋白质溶解度降低并析出,此法分辨力比盐析高,但蛋白质较易变性,应在低温下进行。 (二)根据蛋白质分子大小的差别的分离方法 1、透析与超滤 透析法是利用半透膜将分子大小不同的蛋白质分开。 超滤法是利用高压力或离心力,强使水和其他小的溶质分子通过半透膜,而蛋白质留在膜上,可选择不同孔径的泸膜截留不同分子量的蛋白质。 2、凝胶过滤法 也称分子排阻层析或分子筛层析,这是根据分子大小分离蛋白质混合物最有效的方法之一。柱中最常用的填充材料是葡萄糖凝胶(Sephadex ged)和琼脂糖凝胶(agarose gel)。 (三)根据蛋白质带电性质进行分离 蛋白质在不同pH环境中带电性质和电荷数量不同,可将其分开。 1、电泳法 各种蛋白质在同一pH条件下,因分子量和电荷数量不同而在电场中的迁移率不同而得以分开。值得重视的是等电聚焦电泳,这是利用一种两性电解质作为载体,电泳时两性电解质形成一个由正极到负极逐渐增加的pH梯度,当带一定电荷的蛋白质在其中泳动时,到达各自等电点的pH位置就停止,此法可用于分析和制备各种蛋白质。
一.分子筛(凝胶层析) 原理:用一般的柱层析方法使相对分子质量不同的溶质通过具有分子筛性质的固定相(凝胶),从而使蛋白质分离。 优点:1.洗脱条件简单,往往只需要一种缓冲溶液,可以使用任何缓冲液。 2.实验操作相对简单 3.条件温和,对蛋白活性保持率高 4.既可以对标签蛋白纯化也可以对非标签蛋白纯化。 缺点:1. 工艺放大困难:分子筛层析无法遵循线性放大原则,即使遵循柱床高度不变的原则,工艺流速如何进行调整,也是需要面临的问题。 2. 层析柱装填困难 3.对上样量有要求 4.测定柱效困难 5.反复使用层析柱困难 二.亲和层析 原理:亲和层析是一种吸附层析,亲和层析利用固相介质中的配基与混合生物分子之间亲和能力不同而进行分离,当蛋白混合液通过层析柱时,与配基能够特异性结合的蛋白质就会被吸附固定在层析柱中,其他的蛋白质对配体不具有特异性的结合能力,将通过柱子洗脱下来,这种结合在一定条件下是可逆的,选用适当的洗脱液,改变缓冲液的离子强度和pH 值或者选择更强的配体结合溶液将结合的蛋白质洗脱下来,而无亲和力的蛋白质最先流出层析柱。 优点:1. 亲和层析法是分离蛋白质的一种极为有效的方法,它经常只需经过一步处理即可使某种待提纯的蛋白质从很复杂的蛋白质混合物中分离出来,而且纯度很高。 2. 是最有效的生物活性物质纯化方法,它对生物分子选择性的吸附和分离,可以取得很高的纯化倍数。此外蛋白在纯化过程中得到浓缩,结合到亲和配基后,性质更加稳定,其结果提高了活性回收率。此外它可以减少纯化步骤,缩短纯化时间,对不稳定蛋白的纯化十分有利。 缺点:1.除特异性的吸附外,仍然会因分子的错误认别和分子间非选择性的作用力而吸附一些杂蛋白质,另洗脱过程中的配体不可避免的脱落进入分离体系。 2. 载体较昂贵,机械强度低,配基制备困难,有的配基本身要经过分离纯化,配基与载体耦联条件激烈等。 三.离子交换层析 原理:离子交换层析根据样品表面电荷不同进行分离纯化的技术,根据不同蛋白样品在同一Ph条件下所带电荷正负以及带电荷量不同而将不同蛋白样品分离。 适用范围:适合任何带点生物分子的初步纯化,除去杂质和提纯。 优点:1.领域宽:适合任何带点生物分子的分离(蛋白,多肽,核酸,多糖) 2.高分辨率:只有一个氨基酸区别的样品也可以分离。 3.经过洗脱后可以重复使用层析柱。 4.有亲水性,对大分子吸附性不强,温和条件可洗脱,蛋白质不易变性 缺点:1.定性能力较差 四.高效液相层析 原理:储液器中的流动相被高压泵打入系统,样品溶液经进样器进入流动相,被流动相载入色谱柱(固定相)内,由于样品溶液中的各组分在两相中具有不同的分配系数,在两相中作相对运动时,经过反复多次的吸附-解吸的分配过程,各组分在移动速度上产生较大的差别。适用范围:高效液相色谱更适宜于分离、分析高沸点、热稳定性差、有生理活性及相对分子
分离纯化蛋白质的方法及 原理 This manuscript was revised on November 28, 2020
分离纯化蛋白质的方法及原理 (一)利用分子大小 1、透析:原理:利用蛋白质分子不能透过半透膜的性质,使蛋白质和其他小分子物质如无机盐、单糖、水等分开。 方法:将待提纯蛋白质放在透析袋中放在蒸馏水中进行 涉及的问题: 如何加快透析过程 (1)加大浓度差,及时更换透析液 (2)利用磁力搅拌器 常用的半透膜:玻璃纸、火棉和其他材料合成 2、超过滤:原理:利用压力和离心力,强行使其他小分子和水通过半透膜,而蛋白质留在膜上 3、凝胶过滤层析:原理:当不同分子大小的蛋白质混合物流进凝胶层析柱时,比凝胶网孔大的分子不能进入珠内网状结构,排阻在凝胶珠以外,在凝胶珠缝隙间隙中向下移动。而比孔小的分子不同程度地进入凝胶珠内,这样由于不同大小分子所经历的路径不同而到分离。 结果:大分子先被洗脱下来,小分子后被洗脱下来 (二)利用溶解度差别 4、等电点沉淀:原理:不同蛋白质具有不同的等电点,当蛋白质混合物调到其中一种蛋白质的等电点时,这种蛋白质大部分和全部被沉淀下来.。 5、盐析与盐溶:原理:低浓度时,中性盐可以增加蛋白质溶解度这种现象称为盐溶.当离子强度增加,足够高时,例如饱和或半饱和程度,很多蛋白质可以从水中沉淀出来,这种现象称为盐析
(三)根据电荷不同 6、SDS-PAGE 全称十二烷基硫酸钠—聚丙烯酰胺凝胶电泳 原理:通过加热和SDS可以使蛋白质变性,多亚基的蛋白质也解离为单亚基,处理后的样品中肽链是处于无二硫键连接的,分离的状态。电泳时SDS-蛋白质复合物在凝胶中的迁移率不再受蛋白质原有电荷和形状的影响,而主要取决于蛋白质分子量。所以SDS-PAGE常用来分析蛋白质的纯度和大致测定蛋白质的分子量。 7、离子交换层析:原理:氨基酸分离常用阳离子交换树脂,树脂被处理成钠型,将混合氨基酸上柱,氨基酸主要以阳离子形式存在,在树脂上与钠离子发生交换,而被挂在树脂上。 氨基酸在树脂上结合的牢固程度取决于氨基酸与树脂之间的亲和力,决定亲和力的因素有:(1)主要是静电吸引力(2)氨基酸侧链同树脂之间的疏水作用氨基酸与阳离子交换树脂间的静电引力大小次序依次是: 碱性氨基酸R2+>中性氨基酸R+>酸性氨基酸R0。 因此洗脱顺序应该是: 酸性氨基酸中性氨基酸碱性氨基酸 为使氨基酸从树脂上洗脱下来采用逐步提高pH和盐浓度的方法
蛋白质下游纯化工艺简介 Downstream(下游)与上游相对的概念,一般而言,上游指基因克隆、细胞培养等目的产物表达工序,下游指从表达有目的产物的复杂的混合液中分离纯化得到符合要求的目的产物的一系列步骤。美、日、欧等发达国家生物技术产品工业化的实践证明:生物技术产品的产业化高度依赖于基因工程下游工序技术及设备的进步。 一、纯化工艺常用衔接技术: 1、盐析。常采用硫酸铵、硫酸钠盐析,获得的盐析沉淀物在低温冰箱(<-20℃)中可以长期保存。 2、等电点沉淀。根据蛋白质、多肽在等电点时溶解度最小的特点进行。 3、有机溶剂沉淀。采用冷的丙酮、乙醇沉淀蛋白质。例如人血浆蛋白的乙醇分级沉淀。 4、透析。根据目的蛋白质、多肽的分子量,选取适宜的透析袋进行透析,可以起到更换缓冲液以及去除一些低分子杂质的目的。 5、超滤。根据目的蛋白质、多肽的分子量,选取适宜截留分子量(MWcutoff)的超滤膜进行超滤。 6、真空冷冻干燥。利用在低温和高真空条件下,冰不经融化为液态水而直接升华为水蒸气的原理。可以很好地浓缩样品和保存蛋白质、多肽的生物活性。 7、变性沉淀。根据目的蛋白质、多肽对于温度或者酸碱性的耐受性,在较高的温度或者较极端的pH条件下处理样品,使杂质去除的方法。例如胸腺肽制备中使用的热变性(~80℃)去除杂蛋白。如果目的蛋白、多肽本身热稳定性、酸碱稳定性不高则不宜采用。 8、离心沉淀。利用离心沉降力将不溶性颗粒物与溶液分离的技术,常采用高速冷冻离心机进行。 9、脱盐。透析、超滤可用于进行脱盐,SephadexG25、G50常用于蛋白质脱盐。
10、过滤 澄清过滤、除菌过滤(0.22微米膜过滤)、超滤(1000~300000道尔顿) 二、蛋白质、多肽液相色谱纯化方法简介 chromatography,色谱,层析 表1常用的液相色谱纯化方法及其原理 1、纯化的一般目标和方法 首先,自然来源或者重组表达的蛋白质经过一些粗提的步骤(例如:匀浆、离心、硫酸铵沉淀等)成为稳定的可以用于色谱分离的样品。 然后进行捕获色谱(capturechromatography),主要目标是浓缩和去除大量的容易去除的杂质,此步最关心的是流速和载量,常采用高载量、快流速凝胶。
GST标签蛋白纯化原理 在实验中有过被高等教育的教授骂过:别人让你吃屎,你就去吃屎么?或者稍微高级点的问候声:你有脑子么? 请稍微忽视当事人的失落、自尊心碎一地的感受,苦难不是一种财富,对苦难的反思才是一种财富。在实验时,或许着急按照步骤一步一步的做下来,而没有肯定的问自己:为什么要这样做?为什么要加这样的试剂?所以忽视了一些看似无关紧要的细节,坏结果便发生了。 回归原理部分,回归解决若干个为什么的问题,让我们的实验更加规范化。所以在接下来若干个专题阐述GST标签蛋白纯化的实验时,生物日记实验室部落首先搞清楚GST蛋白纯化的原理。 ——当别人问为什么时,你知道为什么么 Just why? why? why? GST标签蛋白纯化原理 重组标签蛋白纯化利用谷胱甘肽的结构能够和GST(谷胱甘肽S-转移酶)的结合位点互补,谷胱甘肽通过SH基团与琼脂糖介质上
的环氧乙烷基团通过环氧激活特异性偶联在琼脂糖介质上,带有GST的标签蛋白与琼脂糖介质上交联的谷胱甘肽配体互补性结合,这种可逆性的结合在温和、非变性的条件下通过加入还原型谷胱甘肽洗脱下来,杂质通过结合缓冲液被洗脱去除,从而分离目的蛋白。如果需要将GST标签从目的蛋白上除去,则可将蛋白酶结合到柱子上,在标签蛋白与谷胱甘肽结合时使用蛋白酶位点特异性切割,也可在洗脱之后再酶切。另外,蛋白质的性质、载体的选择、宿主菌、表达和纯化的条件不同都能使最终标签蛋白的产量有很大的差别。 目的基因的表达在选择合适的表达载体时,应该注意载体的克隆位点、蛋白酶切位点、标签的位置、编码框等多种因素。 载体的选择 pGEX载体可用于构建可诱导、高水平表达目的基因片段,带有GST标签蛋白通过特定的目的基因或者基因片段插入到pGEX的多克隆位点,在pGEX载体上带有laclp基因,该基因表达产物作为抑制剂结合在tac启动子的操纵子区,而在乳糖类似物IPTG的诱导下,目的基因能够在tac启动子的调控下表达目的蛋白。 宿主菌的选择 在选择宿主菌时,大部分大肠杆菌都能够克隆和表达pGEX载体,如果要获得全长的标签蛋白,一些特殊的工程菌即蛋白酶Lon、OmpT等缺失的菌种能够保护目的蛋白不被宿主菌降解,获得全长的标签蛋白,并且产量也可能更高。 大肠杆菌BL21是获得GST标签蛋白比较合适的宿主菌种,该
硫酸铵分级纯化蛋白质原理操作 硫酸铵分级沉淀蛋白质原理与操作 一,基本原理 硫酸铵沉淀法可用于从大量粗制剂中浓缩和部分纯化蛋白质。用此方法可以将主要的免疫球从样品中分离,是免疫球蛋白分离的常用方法。高浓度的盐离子在蛋白质溶液中可与蛋白质竞争水分子,从而破坏蛋白质表面的水化膜,降低其溶解度,使之从溶液中沉淀出来。各种蛋白质的溶解度不同,因而可利用不同浓度的盐溶液来沉淀不同的蛋白质。这种方法称之为盐析。盐浓度通常用饱和度来表示。硫酸铵因其溶解度大,温度系数小和不易使蛋白质变性而应用最广。 二,试剂及仪器 1 . 组织培养上清液、血清样品或腹水等 2. 硫酸铵(NH 4 )SO 4
3. 饱和硫酸铵溶液(SAS ) 4. 蒸馏水 5. PBS( 含0.2g /L 叠氮钠) 6. 透析袋 7. 超速离心机 8. pH 计 9. 磁力搅拌器 三,操作步骤 以腹水或组织培养上清液为例来介绍抗体的硫酸铵沉淀。各种不同的免疫球蛋白盐析所需硫酸铵的饱和度也不完全相同。通常用来分离抗体的硫酸铵饱和度为33% —50% 。 (一)配制饱和硫酸铵溶液(SAS ) 1.将767g (NH 4 )2 SO 4 边搅拌边慢慢加到1 升蒸馏水中。用氨水或硫酸调到硫酸pH7.0 。此即饱和度为100% 的硫酸铵溶液(4.1 mol/L, 25 °C ).
2.其它不同饱和度铵溶液的配制 (二)沉淀 1.样品(如腹水)20 000 ′g 离心30 min ,除去细胞碎片; 2.保留上清液并测量体积; 3.边搅拌边慢慢加入等体积的SAS 到上清液中,终浓度为1 :1 ( 4.将溶液放在磁力搅拌器上搅拌6 小时或搅拌过夜(4 °C ),使蛋白质充分沉淀。 (三)透析 1.蛋白质溶液10 000 ′g 离心30 min (4 °C )。弃上清保留沉淀; 2.将沉淀溶于少量(10-20ml )PBS -0.2g /L 叠氮钠中。沉淀溶解后放入透析袋对 PBS -0.2g /L 叠氮钠透析24-48 小时(4 °C ),每隔3-6 小时换透析缓冲液一次,以彻底除去硫酸氨; 3.透析液离心,测定上清液中蛋白质含量。 四,应用提示 (一)先用较低浓度的硫酸氨预沉淀,除去样品中的杂蛋白。 1.边搅拌边慢慢加SAS 到样品溶液中,使浓度为0.5:1 (v/v) ; 2.将溶液放在磁力搅拌器上搅拌 6 小时或过夜( 4 ° C ); 3.3000 ′g 离心30 min (4 °C ),保留上清液;上清液再加SAS
蛋白质分离纯化步骤
一、蛋白质分离纯化的一般原则 大多数蛋白质在组织细胞中都是和核酸等生物分子结合在一起,而且每种类型的细胞都含有成千上万种不同的蛋白质。许多蛋白质在结构、性质上有许多相似之处,所以蛋白质的分离提纯是一项复杂的工作。到目前为止,还没有一套现成的方法能把任何一种蛋白质从复杂的混合物中提取出来。但是对于任何一种蛋白质都有可能选择一种较合适的分离纯化程序以获得高纯度的制品。且分离的关键步骤、基本手段还是共同的。 蛋白质提纯的目的是增加产品的纯度和产量,同时又要保持和提高产品的生物活性。因此,要分离纯化某一种蛋白质,首先应选择一种含目的蛋白质较丰富的材料。其次,应设法避免蛋白质变性,以制备有活性的蛋白质。对于大多数蛋白质来说,纯化操作都是在0~4℃的低温下进行的。同时也应避免过酸、过碱的条件以及剧烈的搅拌和振荡。另外,还要设法除去变性的蛋白质和其它杂蛋白,从而达到增加纯度和提高产量的目的。
力不均而使细胞破碎。 5. 酶法 如用溶菌酶破坏微生物细胞等。(二) 蛋白质的抽提 通常选择适当的缓冲液溶剂把蛋白质提取出来。抽提所用缓冲液的pH、离子强度、组成成分等条件的选择应根据欲制备的蛋白质的性质而定。如膜蛋白的抽提,抽提缓冲液中一般要加入表面活性剂(十二烷基磺酸钠、tritonX-100等),使膜结构破坏,利于蛋白质与膜分离。在抽提过程中,应注意温度,避免剧烈搅拌等,以防止蛋白质的变性。 (三)蛋白质粗制品的获得 选用适当的方法将所要的蛋白质与其它杂蛋白分离开来。比较方便的有效方法是根据蛋白质溶解度的差异进行的分离。常用的有下列几种方法: 1. 等电点沉淀法 不同蛋白质的等电点不同,可用等电点沉淀法使它们相互分离。 2. 盐析法 不同蛋白质盐析所需要的盐饱和度