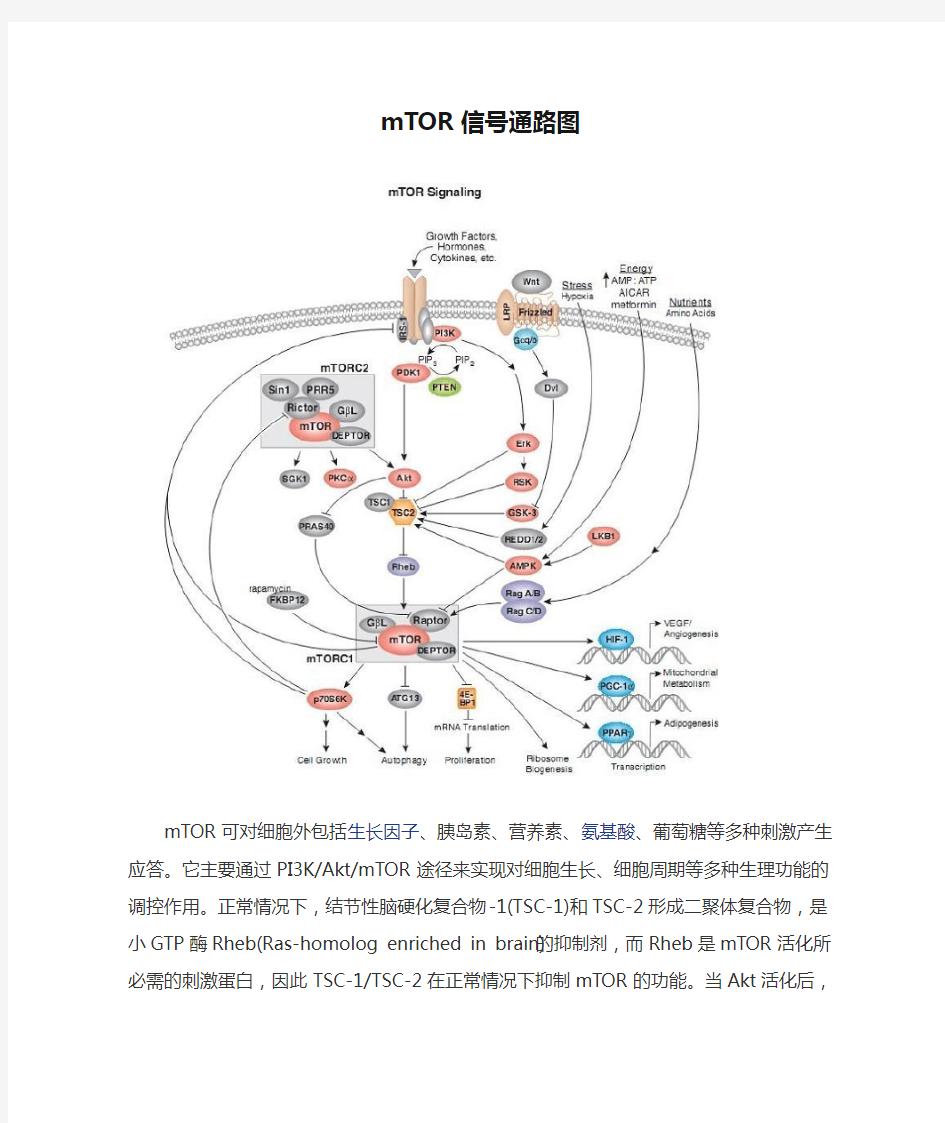

mTOR信号通路图
mTOR可对细胞外包括生长因子、胰岛素、营养素、氨基酸、葡萄糖等多种刺激产生应答。它主要通过PI3K/Akt/mTOR途径来实现对细胞生长、细胞周期等多种生理功能的调控作用。正常情况下,结节性脑硬化复合物-1(TSC-1)和TSC-2形成二聚体复合物,是小GTP 酶Rheb(Ras-homolog enriched in brain)的抑制剂,而Rheb是mTOR活化所必需的刺激蛋白,因此TSC-1/TSC-2在正常情况下抑制mTOR的功能。当Akt活化后,它可磷酸化TSC-2的Ser939和Thr1462,抑制了TSC-1/TSC-2复合物的形成,从而解除了对Rheb 的抑制作用,使得mTOR被激活。活化的mTOR通过磷酸化蛋白翻译过程中的某些因子来参与多项细胞功能,其中最主要的是4EBP1和P70S6K。
在整个PI3K/Akt/mTOR信号通路中,有一条十分重要的负反馈调节剂就是10号染色体上缺失与张力蛋白同源的磷酸酶基因(phosphatase and tensin homology deleted on chromosome 10, PTEN)。PTEN是一个肿瘤抑制基因,位于人染色体10q23。它有一个蛋白酪氨酸磷酸酶结构域,在这条通路中可以将PI-3,4-P2与PI-3,4,5-P3去磷酸化,从而负调节PI3K下游AKt/mTOR信号通路的活性。
本信号转导涉及的信号分子主要包括
IRS-1,PI3K,PIP2,PIP3,PDK1,PTEN,Akt,TSC1,TSC2,Rheb,mTOR,Raptor,DEPTOR,GβL,p70S6K,ATG13,4E-BP1,HIF-1,PGC-1α,PPARγ,Sin1,PRR5,Rictor,PKCα,SGK1,PRAS40,FKBP12,Wnt,LRP,Frizzled,Gαq/o,Dvl,Erk,RSK,GSK-3,REDD1,REDD2,AMPK,LKB1,RagA/B,RagC/D等。
1 PPAR信号通路:过氧化物酶体增殖物激活受体( PPARs) 是与维甲酸、类固醇 和甲状腺激素受体相关的配体激活转录因子超家族核激素受体成员。它们作为脂 肪传感器调节脂肪代谢酶的转录。PPARs由PPARα、PPARβ和PPARγ 3种亚型组成。PPARα主要在脂肪酸代谢水平高的组织,如:肝、棕色脂肪、心、肾和骨骼肌表达。他通过调控靶基因的表达而调节机体许多生理功能包括能量代谢、生 长发育等。另外,他还通过调节脂质代谢的生物感受器而调节细胞生长、分化与 凋亡。PPARa同时也是一种磷酸化蛋白,他受多种磷酸化酶的调节包括丝裂原激活蛋白激酶( ERK-和p38.M APK) ,蛋白激酶A和C( PKA,PKC) ,AM PK和糖原合成酶一3( G SK3) 等调控。调控PPARa生长信号的酶报道有M APK、PKA和G SK3。PPARβ广泛表达于各种组织,而PPAR γ主要局限表达在血和棕色脂肪,其他组织如骨骼肌和心肌有少量表达。PPAR-γ在诸如炎症、动脉粥样硬化、胰岛素抵抗和糖代谢调节,以及肿瘤和肥胖等方面均有着举足轻重的作用, 而其众多生物学效应则是通过启动或参与的复杂信号通路予以实现。鉴于目前人 们对PPAR—γ信号通路尚不甚清,PPARs通常是通过与9-cis维甲酸受体( RXR)结合实现其转录活性的。 2 MAPK信号通路:mapk简介:丝裂原激活蛋白激酶(mitogen—activated protein kinase,MAPK)是广泛存在于动植物细胞中的一类丝氨酸/苏氨酸蛋白激酶。作用主要是将细胞外刺激信号转导至细胞及其核内,并引起细胞的生物化学反应(增殖、分化、凋亡、应激等)。 MAPKs家族的亚族 :ERKs(extracellular signal regulated kinase):包括ERK1、ERK2。生长因子、细胞因子或激素激活此通路,介导细胞增殖、分化。 JNKs(c-Jun N-terminal kinase)包括JNK1、JNK2、JNK3。此亚族成员能使 Jun转录因子N末端的两个氨基酸磷酸化而失活,因此称为Jun N末端激酶(JNKs)。物理、化学的因素引起的细胞外环境变化以及致炎细胞因子调节此通路。P38 MAPKs:丝氨酸/络氨酸激酶,包括p38 α、p38β、p38γ、p38δ。p38 MAP K参与多种细胞内信息传递过程 ,能对多种细胞外刺激发生反应,可磷酸化其它细胞质蛋白,并能从胞浆移位至细胞核而调节转录因子的活性来改变基因的表达水平 ,从而介导细胞生长、发育、分化及死亡的全过程。 ERK5:是一种非典型的MAPK通路,也叫大MAPK通路,只有一个成员。它可被各种刺激因素激活。不仅可以通过磷酸化作用使底物活化,并且通过C端的物理性结合作用激活底物。 3 ERBB信号途径:ErbB 蛋白属于跨膜酪氨酸激酶的 EGF 受体家族成员。ErbB 的命名来源于在禽红白血病 B( v-Erb-B) 发现的 EGF 受体的突变体,因而 EGF 受体 亦称为“ ErbB1”。人源 ErbB2 称为HER2, 特指人的 EGF 受体。ErbB 家族的
1JAK-STAT信号通路 1)JAK与STAT蛋白 JAK-STAT信号通路是近年来发现的一条由细胞因子刺激的信号转导通路,参与细胞的增殖、分化、凋亡以及免疫调节等许多重要的生物学过程。与其它信号通路相比,这条信号通路的传递过程相对简单,它主要由三个成分组成,即酪氨酸激酶相关受体、酪氨酸激酶JAK和转录因子STAT。(1)酪氨酸激酶相关受体(tyrosinekinaseassociatedreceptor) 许多细胞因子和生长因子通过JAK-STAT信号通路来传导信号,这包括白介素2?7(IL-2?7)、GM-CSF(粒细胞/巨噬细胞集落刺激因子)、GH(生 长激素)、EGF(表皮生长因子)、PDGF(血小板衍生因子)以及IFN(干扰素)等等。这些细胞因子和生长因子在细胞膜上有相应的受体。这些受体的共同特点是受体本身不具有激酶活性,但胞内段具有酪氨酸激酶JAK 的结合位点。受体与配体结合后,通过与之相结合的JAK的活化,来磷酸化各种靶蛋白的酪氨酸残基以实现信号从胞外到胞内的转递。 (2)酪氨酸激酶JAK(Januskinase) 很多酪氨酸激酶都是细胞膜受体,它们统称为酪氨酸激酶受体(receptor tyrosinekinase,RTK),而JAK却是一类非跨膜型的酪氨酸激酶。JAK是英文Januskinase的缩写,Janus在罗马神话中是掌管开始和终结的两面神。之所以称为两面神激酶,是因为JAK既能磷酸化与其相结合的细胞因子受体,又能磷酸、JAK1个成员:4蛋白家族共包括JAK结构域的信号分子。SH2化多个含特定
JAK2、JAK3以及Tyk2,它们在结构上有7个JAK同源结构域(JAKhomologydomain,JH),其中JH1结构域为激酶区、JH2结构域是“假”激酶区、JH6和JH7是受体结合区域。 (3)转录因子STAT(signaltransducerandactivatoroftranscription)STAT被称为“信号转导子和转录激活子”。顾名思义,STAT在信号转导和转录激活上发挥了关键性的作用。目前已发现STAT家族的六个成员,即STAT1-STAT6。STAT蛋白在结构上可分为以下几个功能区段:N-端保守序列、DNA结合区、SH3结构域、SH2结构域及C-端的转录激活区。其中,序列上最保守和功能上最重要的区段是SH2结构域,它具有与酪氨酸激酶Src的SH2结构域完全相同的核心序列“GTFLLRFSS”。 2)JAK-STAT信号通路 与其它信号通路相比,JAK-STAT信号通路的传递过程相对简单。信号传 递过程如下:细胞因子与相应的受体结合后引起受体分子的二聚化,这使得与受体偶联的JAK激酶相互接近并通过交互的酪氨酸磷酸化作用而活化。JAK激活后催化受体上的酪氨酸残基发生磷酸化修饰,继而这些磷酸化的酪氨酸位点与周围的氨基酸序列形成“停泊位点”(dockingsite),同时含有SH2结构域的STAT蛋白被招募到这个“停泊位点”。最后,激酶JAK 催化结合在受体上的STAT蛋白发生磷酸化修饰,活化的STAT蛋白以二 聚体的形式进入细胞核内与靶基因结合,调控基因的转录。值得一提的是,一种JAK激酶可以参与多种细胞因子的信号转导过程,一种细胞因子的信号通路也可以激活多个JAK激酶,但细胞因子对激活的STAT分子却具有一定的选择性。例如IL-4激活STAT6,而IL-12 。STAT4却特异性激活
干货细胞信号通路图解之MAPK通路【值得珍藏】 科研小助手原创,转载请注明来源。公众号内回复“Cell Signaling Pathway”获取全套信号通路图本文由百度贴吧nosce吧吧主黄杰投稿一、MAPK信号通路: (1)有丝分裂原激活的蛋白激酶(MAPK)是一族在真核生物中非常保守的丝/苏氨酸蛋白激酶,在许多细胞活动中起作用,如生长增殖,细胞分化,细胞运动或死亡。MAPK级联信号传导由3 个不同层次的分子所组成。MAPK被MAPK的激 酶( MAPKK)磷酸化后激活,MAPKK被MAPKK的激酶(MAPKKK )磷酸化而激活。而MAPKKK通过与小GTPase 和/或其他蛋白酶相互作用而被激活,从而将MAPK和细胞 表面的受体以及胞外的信号联系在一起。 (2)许多参与生长和分化的受体都能够激活MAPK/ERK信号通路,比如说受体酪氨酸激酶(RTK),整合素,和离子通道。响应特定信号所涉及到的具体分子会相差很大,但通路的结构是一致的,那就是接头分子(adaptor,如Shc, GRB2, Crk等)将鸟苷酸交换因子(SOS, C3G 等)和受体连接在一起,然后把信号向小GTP 结合蛋白(Ras, Rap1)传递,后者又激活核心的级联反应,这是由一个MAPKKK( Raf) ,一个MAPKK( MEK1/2)和MAPK( Erk)所构成的。活化的ERK 二聚体能调节胞浆中的目标分子,也可以转移到细胞核中,然
后对一系列转录因子进行磷酸化以调节基因表达。SciRes(3)很多外部的刺激都能够激活G蛋白偶联受体(GPCR)。在受体活化以后,G 蛋白将GDP 转换成GTP ,然后结合了GTP的α和β/γ亚基从受体脱离开,启动信号向胞内的传导。与不同亚型的异质三聚体G 蛋白结合的受体可以采取不同 的手段激活小G 蛋白/MAPK级联反应,至少有三个不同家族的酪氨酸激酶参与其中。Src家族激酶响应活化的PI3Kγ,而后者被β/γ亚基激活。它们还能够响应受体的内化,受体酪氨酸激酶的交叉活化,以及有Pyk2 和/或FAK参与的整 合素途径信号。GPCRs同样可以通过PLCβ去激活PKC 和CaMKII ,对下游的MAPK通路可以有激活或抑制的影响。SciRes(4)压力激活的蛋白激酶(Stress-activated protein kinase, SAPK)或称Jun氨基端激酶(Jun amino-terminal kinase, JNK) 是MAPK的家族成员,能被一系列的环境压力,炎症细胞因子,生长因子和GPCR激动剂所激活。压力信号通过Rho家族的小GTP 酶(small GTPase)向这条级联通路传导,这些小GTP酶包括(Rac, Rho, cdc42) 。和其他的MAPK情况一样,靠近膜的激酶是一个MAPKKK,一般 是MEKK1-4 ,或者是一个混合激酶去磷酸化并激活 MKK4(SEK)或MKK7,它们是SAPK/JNK的激酶。另外,MKK4/7也可以被生发中心激酶(germinal center kinase, GCK)以一种GTPase 依赖的方式激活。活化后的
目录 actin肌丝 (5) Wnt/LRP6 信号 (7) WNT信号转导 (7) West Nile 西尼罗河病毒 (8) Vitamin C 维生素C在大脑中的作用 (10) 视觉信号转导 (11) VEGF,低氧 (13) TSP-1诱导细胞凋亡 (15) Trka信号转导 (16) dbpb调节mRNA (17) CARM1甲基化 (19) CREB转录因子 (20) TPO信号通路 (21) Toll-Like 受体 (22) TNFR2 信号通路 (24) TNFR1信号通路 (25) IGF-1受体 (26) TNF/Stress相关信号 (27) 共刺激信号 (29) Th1/Th2 细胞分化 (30) TGF beta 信号转导 (32) 端粒、端粒酶与衰老 (33) TACI和BCMA调节B细胞免疫 (35) T辅助细胞的表面受体 (36) T细胞受体信号通路 (37) T细胞受体和CD3复合物 (38) Cardiolipin的合成 (40) Synaptic突触连接中的蛋白 (42) HSP在应激中的调节的作用 (43) Stat3 信号通路 (45) SREBP控制脂质合成 (46) 酪氨酸激酶的调节 (48) Sonic Hedgehog (SHH)受体ptc1调节细胞周期 (51) Sonic Hedgehog (Shh) 信号 (53) SODD/TNFR1信号 (56) AKT/mTOR在骨骼肌肥大中的作用 (58) G蛋白信号转导 (59) IL1受体信号转导 (60) acetyl从线粒体到胞浆过程 (62) 趋化因子chemokine在T细胞极化中的选择性表达 (63) SARS冠状病毒蛋白酶 (65) SARS冠状病毒蛋白酶 (67) Parkin在泛素-蛋白酶体中的作用 (69)
mTOR信号通路图 mTOR可对细胞外包括生长因子、胰岛素、营养素、氨基酸、葡萄糖等多种刺激产生应答。它主要通过PI3K/Akt/mTOR途径来实现对细胞生长、细胞周期等多种生理功能的调控作用。正常情况下,结节性脑硬化复合物-1(TSC-1)和TSC-2形成二聚体复合物,是小GTP 酶Rheb(Ras-homolog enriched in brain)的抑制剂,而Rheb是mTOR活化所必需的刺激蛋白,因此TSC-1/TSC-2在正常情况下抑制mTOR的功能。当Akt活化后,它可磷酸化TSC-2的Ser939和Thr1462,抑制了TSC-1/TSC-2复合物的形成,从而解除了对Rheb 的抑制作用,使得mTOR被激活。活化的mTOR通过磷酸化蛋白翻译过程中的某些因子来参与多项细胞功能,其中最主要的是4EBP1和P70S6K。
在整个PI3K/Akt/mTOR信号通路中,有一条十分重要的负反馈调节剂就是10号染色体上缺失与张力蛋白同源的磷酸酶基因(phosphatase and tensin homology deleted on chromosome 10, PTEN)。PTEN是一个肿瘤抑制基因,位于人染色体10q23。它有一个蛋白酪氨酸磷酸酶结构域,在这条通路中可以将PI-3,4-P2与PI-3,4,5-P3去磷酸化,从而负调节PI3K下游AKt/mTOR信号通路的活性。 本信号转导涉及的信号分子主要包括 IRS-1,PI3K,PIP2,PIP3,PDK1,PTEN,Akt,TSC1,TSC2,Rheb,mTOR,Raptor,DEPTOR,GβL,p70S6K,ATG13,4E-BP1,HIF-1,PGC-1α,PPARγ,Sin1,PRR5,Rictor,PKCα,SGK1,PRAS40,FKBP12,Wnt,LRP,Frizzled,Gαq/o,Dvl,Erk,RSK,GSK-3,REDD1,REDD2,AMPK,LKB1,RagA/B,RagC/D等。
KEGG上的信号通路图怎么看? 提示:请点击标题下方蓝色“实验万事屋”,添加关注后,发“嗯”可以查看我们之前的文章。未经允许,其他公众号不得转载哦! 想要把自己研究的分子扯上明星分子或者明星通路?那是不难,难的是具体到底要怎么去扯,芯片结果啊,生信结果啊,都会给你提示,但真的要具体扯上去,还得看懂那些七七八八的信号通路图。 KEGG Pathway上有着大量的信号通路图,画得一个复杂啊!巨坑爹有没有?曾经有师弟说我之前曾经把Wnt通路描述错了,他师兄告诉他,应该是GSK-3β磷酸化抑制β-Catenin降解,并促进它入核的。在这里,我们只能默默地祝福这位师兄了…… 那我们就用Wnt通路来做例子吧。先上KEGG下载一个Wnt的信号通路图,如下: 绝壁是很高大上的不是么?这要咋看呢?其实这张图上把三个Wnt通路都画上去了,也就是Wnt/β-Catenin(经典Wnt通路),Wnt/PCP(平面的细胞极性途径)和Wnt/Ca2+(Wnt/钙离子)三条信号通路组成,我们就删减一下,就光看经典的Wnt通路,就变成了下面这个模样:
感觉还是很高大上有木有?那就再删减一下,把它变成经典Wnt信号通路的骨架会是什么样呢?就是这样: 简洁明快了吧,但要怎么来看懂这样的图呢?我们来看一下KEGG Pathway的具体图例:
把这些图例用来解释经典Wnt信号通路骨架图,就变成了: 看懂了么?那给你从左到右解释一下: 1)Wnt激活膜上受体,将信号传递到第二信使Dvl,活化的Dvl抑制由Axin、APC 和GSK-3β组成的复合物的活性,使β-catenin不能被GSK-3β磷酸化。 2)磷酸化的β-catenin才可通过泛素化(ubiquitination)而被胞浆内的蛋白酶体所降解,由于非磷酸化的β-catenin不能被蛋白酶体降解,从而导致β-catenin在胞浆内积聚,并移向核内。
Toll样受体信号通路图 TLR家族成员(TLR3除外)诱导的炎症反应都经过一条经典的信号通路(图1),该通路起始于TLRs的一段胞内保守序列—Toll/IL-1受体同源区(Toll/IL-1receptorhomologousregion,TIR).TIR可激活胞内的信号介质—白介素1受体相关蛋白激酶(IL-1Rassociatedkinase,IRAK)IRAK-1和IRAK-4、肿瘤坏死因子受体相关因子6(TNFR-associatedfactor6,TRAF-6)、促分裂原活化蛋白激酶(mitogenactivatedproteinkinase,MAPK)和IκB激酶(IκBkinase,IκK),进而激活核因子κB(nuclearfactorκB,NF-κB),诱导炎症因子的表达。 Toll-liker Receptor Signaling 本信号转导涉及的信号分子主要包括: CD14,MD-2,TRAM,TRIF,TIRAP,MyD88,TLR1,TLR2,TLR3,TLR4,TLR5,TLR6,TLR7,TLR8,TLR9,IRAK-1,IRAK-2,IRAK-4,IRAK-M,TRAF6,TRIAD3A,ST2L,SOCS1,RIG-I,FADD,TOLLIP,RIP1,A20,UEV1A,Ubc13,ECSIT,MEKK-1,TAK1,
TBK1,MKK3/6,p38,TAB1/2,MKK4/7,JNK,IKKα,IKKβ,IKKγ,IKKε,NEMO,IκBα,NF-κB,p65/RelA,Casp-8,IRF-3,IRF-7,MA VS等
最新细胞各种信号通路《Cell》 SnapShots are handy reference guides, carefully designed to highlight the key information on a particular topic on one page. SnapShots present up-to-date tables of nomenclature and glossaries, full signaling pathways, and schematic diagrams of cellular processes.Snapshots in red are FREE[/B]. Actin Regulators I[/url] Actin Regulators II[/url]
Antibiotic Inhibition of Protein Synthesis I[/url] Antibiotic Inhibition of Protein Synthesis II[/url] ENHANCED[/url]
Auxin Signaling and Transport Bacterial Appendages I Bacterial Appendages II B7/CD28 Costimulation
BCL-2 Proteins Ca2+-Calcineurin-NF A T Signaling
Ca2+-Dependent Transcription in Neurons Cell-Cycle Regulators I
1 PPAR 信号通路:过氧化物酶体增殖物激活受体( PPARs) 是与维甲酸、类固醇和甲状腺激素受体相关的配体激活转录因子超家族核激素受体成员。它们作为脂肪传感器调节脂肪代谢酶的转录。PPARs由PPAR a、PPAR B和PPAR 丫3种亚型组成。PPAR a主要在脂肪酸代谢水平高的组织,如:肝、棕色脂肪、心、肾和骨骼肌表达。他通过调控靶基因的表达而调节机体许多生理功能包括能量代谢、生长发育等。另外,他还通过调节脂质代谢的生物感受器而调节细胞生长、分化与凋亡。PPARa 同时也是一种磷酸化蛋白,他受多种磷酸化酶的调节包括丝裂原激活蛋白激酶(ERK-和p38 . M APK),蛋白激酶A和C( PKA , PKC) , AM PK和糖原合成酶一3( G SK3) 等调控。调控PPARa 生长信号的酶报道有M APK 、PKA和G SK3。PPAR B广泛表达于各种组织,而PPAR 丫主要局限表达在血和棕色脂肪,其他组织如骨骼肌和心肌有少量表达。PPAR- 丫在诸如炎症、动脉粥样硬化、胰岛素抵抗和糖代谢调节,以及肿瘤和肥胖等方面均有着举足轻重的作用,而其众多生物学效应则是通过启动或参与的复杂信号通路予以实现。鉴于目前人们对PPAR―丫信号通路尚不甚清,PPARs通常是通过与9-cis维甲酸受体(RXR) 结合实现其转录活性的。 2 MAPK信号通路:mapk简介丝裂原激活蛋白激酶(mitogen —activated protein kinase ,MAPK )是广泛存在于动植物细胞中的一类丝氨酸/苏氨酸蛋白激酶。作用主要是将细胞外刺激信号转导至细胞及其核内,并引起细胞的生物化学反应(增殖、分化、凋亡、应激等)。 MAPKs 家族的亚族:ERKs (extracellular signal regulated kinase) :包括ERK1、 ERK2。生长因子、细胞因子或激素激活此通路,介导细胞增殖、分化。 JNKs(c-Jun N-terminal kinase) 包括JNK1、JNK2、JNK3。此亚族成员能使 Jun 转录因子N 末端的两个氨基酸磷酸化而失活,因此称为Jun N 末端激酶(JNKs)。物理、化学的因素引起的细胞外环境变化以及致炎细胞因子调节此通路。 P38 MAPKs :丝氨酸/ 络氨酸激酶,包括p38 a、p38 B、p38 丫、p38 3。p38 MAP K 参与多种细胞内信息传递过程,能对多种细胞外刺激发生反应,可磷酸化其它细胞质蛋白,并能从胞浆移位至细胞核而调节转录因子的活性来改变基因的表达水平,从而介导细胞生长、发育、分化及死亡的全过程。 ERK5:是一种非典型的MAPK通路,也叫大MAPK通路,只有一个成员。它可被各种刺激因素激活。不仅可以通过磷酸化作用使底物活化,并且通过 C 端的物理性结合作用激活底物。 3 ERBB信号途径:ErbB蛋白属于跨膜酪氨酸激酶的EGF受体家族成员。ErbB的命名来源于在禽红白血病B( v-Erb-B) 发现的EGF 受体的突变体,因而EGF 受体亦称为“ ErbB1 ” 。人源ErbB2 称为HER2, 特指人的EGF 受体。ErbB 家族的另外两个
p i k a k t信号通路图谱 WTD standardization office【WTD 5AB- WTDK 08- WTD 2C】
P I3K/A K T信号通路 磷脂酰肌醇3-激酶(PI3Ks)信号参与增殖、分化、凋亡和葡萄糖转运等多种细胞功能的调节. 近年来发现, IA型PI3K和其下游分子蛋白激酶B(PKB或Akt)所组成的信号通路与人类肿瘤的发生发展密切相关. 该通路调节肿瘤细胞的增殖和存活, 其活性异常不仅能导致细胞恶性转化, 而且与肿瘤细胞的迁移、黏附、肿瘤血管生成以及细胞外基质的降解等相关, 目前以PI3K-Akt信号通路关键分子为靶点的肿瘤治疗策略正在发展中. 在PI3K家族中, 研究最广泛的是能被细胞表面受体所激活的I型PI3K. 哺乳动物细胞中Ι型PI3K又分为IA和IB两个亚型, 他们分别从酪氨酸激酶连接受体和G蛋白连接受体传递信号.IA 型PI3K是由催化亚单位p110和调节亚单位p85所组成的二聚体蛋白, 具有类脂激酶和蛋白激酶的双重活性.PI3K通过两种方式激活, 一种是与具有磷酸化酪氨酸残基的生长因子受体或连接蛋白相互作用, 引起二聚体构象改变而被激活; 另一种是通过Ras和p110直接结合导致PI3K的活化. PI3K激活的结果是在质膜上产生第二信使PIP3, PIP3与细胞内含有PH结构域的信号蛋白Akt和 PDK1(phosphoinositidedependentkinase-1)结合, 促使PDK1磷酸化Akt蛋白的 Ser308导致Akt的活化. Akt还能通过PDK2(如整合素连接激酶ILK)对其Thr473的磷酸化而被激活.活化的Akt通过磷酸化作用激活或抑制其下游靶蛋白Bad 、 Caspase9、NF-κB、GSK-3、FKHR、 p21Cip1和p27 Kip1等, 进而调节细胞的增殖、分化、凋亡以及迁移等. PI3K-Akt信号通路的活性被类脂磷酸酶PTEN(phosphatase and tensin homolog deleted on chromosome ten)和SHIP(SH2-containing inositol 5-phosphatase)负调节, 他们分别从PIP3的3′和5′去除磷酸而将其转变成PI(4,5)P2和PI(3,4)P2而 降解. 迄今为止, 尚未发现下调Akt活性的特异磷酸酶, 但用磷酸酶抑制剂处理细胞后, 发现Akt的磷酸化和活性均有所增加. 最近发现Akt能被一种C末端调节蛋白(CTMP)所失活, CTMP能结合Akt并通过抑制Akt的磷酸化而阻断下游信号的传递, CTMP的过表达能够逆转v-Akt转化细胞的表型. 热休克蛋白90(HSP90)亦能结合Akt, 阻止Akt被PP2A磷酸酶的去磷酸化而失活, 因此具有保护Akt的作用. 本信号转导涉及的信号分子主要包括 Integrin,FAK,Paxillin,ILK,PIP3,S6,p70S6K,RTK,Gab1,Gab2,IRS-1, PI3K,PTEN,AKT,PDK1,Cytokine Receptor,Jak1,CD19,BCR,Ag,BCAP,Syk,Lyn,GPCR,TSC1,TSC2,Gβγ,GαGTP,PP2A,PHLPP,CTMP,PDCD4,4E-BP1,ATG13,mTORC1,TSC1,TSC2,PRAS40,XIAP,FoxO1,Bim,Bcl-2,Bax,MDM2, p53,Bax,Bad,14-3-3,Wee1,Myt1,p27Kip1,p21Waf1/Cip1,CyclinD1,GSK-3,GS,Bcl-2,mTORC2,LaminA,Tpl2,IKKα,eNOS,GABAAR,Huntingtin,Ataxin-1,PFKFB2,PIP5K,AS160等。
MAPK/ERK信号通路图及简介 MAPK信号通路专题 MAPK,丝裂原活化蛋白激酶(mitogen-activated protein kinases,MAPKs)是细胞内的一类丝氨酸/苏氨酸蛋白激酶。研究证实,MAPKs信号转导通路存在于大多数细胞内,在将细胞外刺激信号转导至细胞及其核内,并引起细胞生物学反应(如细胞增殖、分化、转化及凋亡等)的过程中具有至关重要的作用。研究表明,MAPKs信号转导通路在细胞内具有生物进化的
高度保守性,在低等原核细胞和高等哺乳类细胞内,目前均已发现存在着多条并行的MAPKs 信号通路,不同的细胞外刺激可使用不同的MAPKs信号通路,通过其相互调控而介导不同的细胞生物学反应。 ERK(extracellular signal-regulated kinase)信号通路1986年由Sturgill等人首先报告的MAPK。最初其名称十分混乱,曾根据底物蛋白称之为MAP2K、ERK、MBPK、RSKK、ERTK等。此后,由于发现其具有共同的结构和生化特征,而被命名为MAPK。近年来,随着不同MAPK家族成员的发现,又重新改称为ERK。 哺乳类动物细胞中,与ERK相关的细胞内信号转导途径被认为是经典MAPK信号转导途径,目前对其激活过程及生物学意义已有了较深入的认识。研究证实,受体酪氨酸激酶、G蛋白偶联的受体和部分细胞因子受体均可激活ERK信号转导途径。如:生长因子与细胞膜上的特异受体结合,可使受体形成二聚体,二聚化的受体使其自身酪氨酸激酶被激活;受体上磷酸化的酪 氨酸又与位于胞膜上的生长因子受体结合蛋白2(Grb2)的SH2结构域相结合,而Grb2的SH3结构域则同时与鸟苷酸交换因子SOS(Son of Sevenless)结合,后者使小分子鸟苷酸结合蛋白Ras的GDP解离而结合GTP,从而激活Ras;激活的Ras进一步与丝/苏氨酸蛋白激酶Raf-1的氨基端结合,通过未知机制激活Raf-1;Raf-1可磷酸化MEK1/MEK2(MAP kinase/ERK kinase)上的二个调节性丝氨酸,从而激活MEKs;MEKs为双特异性激酶,可以使丝/苏氨酸和酪氨酸发生磷酸化,最终高度选择性地激活ERK1和ERK2(即p44MAPK 和p42MAPK)。ERKs为脯氨酸导向的丝/苏氨酸激酶,可以磷酸化与脯氨酸相邻的丝/苏氨酸。在丝裂原刺激后,ERKs接受上游的级联反应信号,可以转位进入细胞核。因此,ERKs 不仅可以磷酸化胞浆蛋白,而且可以磷酸化一些核内的转录因子如c-fos、c-Jun、Elk-1、c-myc和ATF2等,从而参与细胞增殖与分化的调控。另外,ERK还可以磷酸化ERKs通路的上游蛋白如NGF受体、SOS、Raf-1、MEK等,进而对该通路进行自身的负反馈调节。还有研究发现,ERKs可磷酸化胞浆内的细胞骨架成份,如微管相关蛋白MAP-1、MAP-2和MAP-4,参与细胞形态的调节及细胞骨架的重分布。 最近,国外学者又新克隆出ERK5及其上游激酶MEK5,这条MAPKs信号通路可被H2O2 及高渗刺激活[2],其底物为c-Myc。通过分子生物学技术发现还有ERK3 Kinase/ERK3及ERK4两条通路存在,但目前对其激活信号、底物及生物学意义还不清楚。
Cell Signalling Pathways --Michael J. Berridge--module 2 胞内信号通路可分为两类,大多数的信号通路受细胞表面的胞外信号刺激,通常以化学信号的形式,如神经递质、激素及生长因子等;其他类的信号通路是由细胞内产生的信号激活的。胞内信号主要来自内质网或代谢物。 一、环腺苷酸信号通路(Cyclic AMP signalling pathway) 环腺苷酸是广泛存在的一种第二信使,其形成依赖于GPCR的活化,GPCR通过异质三聚体激活放大器AC(腺苷酸环化酶)。cAMP的信号效应器有PKA、EPACs等可激活小GTP连接蛋白Rap1及环核苷酸门控通道(CNGCs),这些效应器负责进行cAMP信号功能。cAMP 的许多功能取决于PKA的准确定位,而A激酶锚定蛋白(AKAPs)家族约定了PKA及其他许多信号组分的细胞定位。 Cyclic AMP formation 环腺苷酸的形成可被许多细胞刺激活化,主要是神经递质和激素,这些刺激可被G蛋白偶联受体通过异质三聚体G蛋白检测到。在腺苷酸环化酶刺激下,外部刺激结合到G蛋白偶
联受体上,作为鸟苷酸交换因子(GEF)用GTP替代GDP,从而使得异质三聚体G蛋白分裂成Gβ?和Gα亚基。Gα亚基和GTP的复合体激活腺苷酸环化酶,然而抑制性GαGTP 抑制AC。Gα亚基具有GTP酶活性,可水解GTP成GDP,因而停止其对AC的作用。Adenylyl cyclase (AC) AC家族由十个亚型组成,前九个为膜结合的,另外一个是水溶性的。AC1-9的域结构具有两个含六个转膜区的区域。大的细胞浆域C1和C2含有催化区,形成异质二聚体使得ATP 转化成AMP。 Cyclic AMP signalling effectors EPACs、CNGCs等,cAMP的大多数作用都是通过PKA发挥作用的。 Protein kinase A (PKA) PKA由两个调节亚基(R)和两个催化亚基(C)组成。环腺苷酸激活PKA是通过结合到R 亚基上,然后使C亚基磷酸化大量的不同底物。在两种PKA中,PKA1主要游离在细胞浆中,对环腺苷酸具有高亲和力,然而PKA2可更准确定位,与A激酶锚定蛋白偶联。AKAPs 是一种支架蛋白的例子,在信号通路的空间组织中,其可把PKA带到适当位置与其底物接触。 Protein kinase A (PKA) I PKA1与RI亚型结合。对所有亚型来说,R亚基通过其N末端二聚化/对接域形成二聚体。除了使两个R亚基在一起,N末端也负责与AKAPs对接(针对PKA2)。RI对AKAPs低亲和,所以主要是水溶的。cAMP连接到串联环腺苷酸结合域释放活性C亚基,然后磷酸化特定底物。RI对环腺苷酸具有高的结合亲和力,所以PKA1可对更低浓度环腺苷酸反应。Protein kinase A (PKA) II PKA2的特点是其调节二聚体是有RII亚基组成的。PKA2更容易与AKAPs对接,从而能更准确的定位到特定的细胞靶位。 环腺苷酸的磷酸化底物可分为主要两组,调节特点细胞过程的底物和其他信号系统组分的底
?·NGF信号通路(2004-8-16) ?·TGF beta 信号转导(2004-8-16) ?·细胞凋亡信号(2004-8-16) ?·线粒体输入信号(2004-8-16) ?·ROS信号(2004-8-16) ?·Toll-Like 受体家族(2004-8-16) ?·Toll-Like 受体(2004-8-16) ?·actin肌丝(2004-8-16) ?·Wnt/LRP6 信号(2004-8-16) ?·WNT信号转导(2004-8-16) ?·West Nile 西尼罗河病毒(2004-8-16) ?·Vitamin C 维生素C在大脑中的作用(2004-8-16) ?·视觉信号转导(2004-8-16) ?·VEGF,低氧(2004-8-16) ?·TSP-1诱导细胞凋亡(2004-8-16) ?·Trka信号转导(2004-8-16) ?·dbpb调节mRNA (2004-8-16) ?·CARM1甲基化(2004-8-16) ?·CREB转录因子(2004-8-16) ?·TPO信号通路(2004-8-16) ?·Toll-Like 受体(2004-8-16) ?·TNFR2 信号通路(2004-8-16) ?·TNFR1信号通路(2004-8-16) ?·TNF/Stress相关信号(2004-8-16) ?·IGF-1受体(2004-8-16) ?·共刺激信号(2004-8-16) ?·Th1/Th2 细胞分化(2004-8-16) ?·TGF beta 信号转导(2004-8-16) ?·端粒、端粒酶与衰老(2004-8-16) ?·TACI和BCMA调节B细胞免疫(2004-8-16) ?·T辅助细胞的表面受体(2004-8-16) ?·T细胞受体信号通路(2004-8-16) ?·T细胞受体和CD3复合物(2004-8-16) ?·Cardiolipin的合成(2004-8-16) ?·Synaptic突触连接中的蛋白(2004-8-16) ?·HSP在应激中的调节的作用(2004-8-16) ?·Stat3 信号通路(2004-8-16) ?·SREBP控制脂质合成(2004-8-16) ?·酪氨酸激酶的调节(2004-8-16) ?·Sonic Hedgehog (SHH)受体ptc1调节细胞周期(2004-8-16)?·Sonic Hedgehog (Shh) 信号(2004-8-16) ?·SODD/TNFR1信号(2004-8-16) ?·AKT/mTOR在骨骼肌肥大中的作用(2004-8-16) ?·G蛋白信号转导(2004-8-16)
细胞信号转导途径研究方法 一、蛋白质表达水平和细胞内定位研究 1、信号蛋白分子表达水平及分子量检测: Western blot analysis. 蛋白质印迹法是将蛋白质混合样品经SDS-PAGE后,分离为不同条带,其中含有能与特异性抗体(或McAb)相应的待检测的蛋白质(抗原蛋白),将PAGE胶上的蛋白条带转移到NC膜上此过程称为blotting,以利于随后的检测能够的进行,随后,将NC膜与抗血清一起孵育,使第一抗体与待检的抗原决定簇结合(特异大蛋白条带),再与酶标的第二抗体反应,即检测样品的待测抗原并可对其定量。 基本流程: 检测示意图:
2、免疫荧光技术Immunofluorescence (IF) 免疫荧光技术是根据抗原抗体反应的原理,先将已知的抗原或抗体标记上荧光素制成荧光标记物,再用这种荧光抗体(或抗原)作为分子探针检查细胞或组织内的相应抗原(或抗体)。在细胞或组织中形成的抗原抗体复合物上含有荧光素,利用荧光显微镜观察标本,荧光素受激发光的照射而发出明亮的荧光(黄绿色或桔红色),可以看见荧光所在的细胞或组织,从而确定抗原或抗体的性质、定位,以及利用定量技术测定含量。 采用流式细胞免疫荧光技术(FCM)可从单细胞水平检测不同细胞亚群中的蛋白质分子,用两种不同的荧光素分别标记抗不同蛋白质分子的抗体,可在同一细胞内同时检测两种不同的分子(Double IF),也可用多参数流式细胞术对胞内多种分子进行检测。 二、蛋白质与蛋白质相互作用的研究技术 1、免疫共沉淀(Co- Immunoprecipitation, Co-IP)
Co-IP是利用抗原蛋白质和抗体的特异性结合以及细菌蛋白质的“protein A”能特异性地结合到免疫球蛋白的FC片段的现象而开发出来的方法。目前多用精制的protein A预先结合固化在agarose的beads上,使之与含有抗原的溶液及抗体反应后,beads上的prorein A就能吸附抗原抗体达到沉淀抗原的目的。 当细胞在非变性条件下被裂解时,完整细胞内存在的许多蛋白质-蛋白质间的相互作用被保留了下来。如果用蛋白质X的抗体免疫沉淀X,那么与X在体内结合的蛋白质Y也能沉淀下来。进一步进行Western Blot 和质谱分析。这种方法常用于测定两种目标蛋白质是否在体内结合,也可用于确定一种特定蛋白质的新的作用搭档。缺点:可能检测不到低亲和力和瞬间的蛋白质-蛋白质相互作用。 2、GST pull-down assay GST pull-down assay是将谷胱甘肽巯基转移酶(GST)融合蛋白(标记蛋白或者饵蛋白,GST, His6, Flag, biotin …)作为探针,与溶液中的特异性搭档蛋白(test protein或者prey被扑获蛋白)结合,然后根据谷胱甘肽琼脂糖球珠能够沉淀GST融合蛋白的能力来确定相互作用的蛋白。一般在发现抗体干扰蛋白质-蛋白质之间的相互作用时,可以启用GST沉降技术。该方法只是用于确定体外的相互作用。
常见的几种信号通路 1 JAK-STAT信号通路 1) JAK与STAT蛋白 JAK-STAT信号通路是近年来发现的一条由细胞因子刺激的信号转导通路,参与细胞的增殖、分化、凋亡以及免疫调节等许多重要的生物学过程。与其它信号通路相比,这条信号通路的传递过程相对简单,它主要由三个成分组成,即酪氨酸激酶相关受体、酪氨酸激酶JAK 和转录因子STAT。 (1) 酪氨酸激酶相关受体(tyrosine kinase associated receptor) 许多细胞因子和生长因子通过JAK-STAT信号通路来传导信号,这包括白介素2?7(IL-2?7)、GM-CSF(粒细胞/巨噬细胞集落刺激因子)、GH(生长激素)、EGF(表皮生长因子)、PDGF (血小板衍生因子)以及IFN(干扰素)等等。这些细胞因子和生长因子在细胞膜上有相应的受体。这些受体的共同特点是受体本身不具有激酶活性,但胞内段具有酪氨酸激酶JAK的结合位点。受体与配体结合后,通过与之相结合的JAK的活化,来磷酸化各种靶蛋白的酪氨酸残基以实现信号从胞外到胞内的转递。 (2) 酪氨酸激酶JAK(Janus kinase) 很多酪氨酸激酶都是细胞膜受体,它们统称为酪氨酸激酶受体(receptor tyrosine kinase, RTK),而JAK却是一类非跨膜型的酪氨酸激酶。JAK是英文Janus kinase 的缩写,Janus在罗马神话中是掌管开始和终结的两面神。之所以称为两面神激酶,是因为JAK既能磷酸化与其相结合的细胞因子受体,又能磷酸化多个含特定SH2结构域的信号分子。JAK蛋白家族共包括4个成员:JAK1、JAK2、JAK3以及Tyk2,它们在结构上有7个JAK同源结构域(JAK homology domain, JH),其中JH1结构域为激酶区、JH2结构域是“假”激酶区、JH6和JH7是受体结合区域。 (3) 转录因子STAT(signal transducer and activator of transcription)STAT被称为“信号转导子和转录激活子”。顾名思义,STAT在信号转导和转录激活上发挥了关键性的作用。目前已发现STAT家族的六个成员,即STAT1-STAT6。STAT蛋白在结构上可分为以下几个功能区段:N-端保守序列、DNA结合区、SH3结构域、SH2结构域及C-端的转录激活区。其中,序列上最保守和功能上最重要的区段是SH2结构域,它具有与酪氨酸激酶Src 的SH2结构域完全相同的核心序列“GTFLLRFSS”。 2) JAK-STAT信号通路
NF-kappaB是一个大家族,包括:RelA(p65)、c-Rel、RelB、NF-kappaB1 (p50/p105)、NF-kappaB2 (p52/p100)。其中以RelA(p65)研究最为深入。 通常所说的是左边的经典途径,大致意思是这样:非激活状态下,RelA(p65)与一种名为IkappaB的蛋白结合,停留在胞浆中,不发挥转录活性。当炎症因子等刺激时,IkappaB的上游激酶IkappaB kinase(IKK)磷酸化而激活。IKK使IkappaB降解。p50有核定位信号,没有了IkappaB 的束缚,p50会拽着RelA(p65)往核里面跑。RelA(p65)识别特定的DNA序列,并结合上去,从而调控靶基因的转录(这些基因的启动子上含有RelA(p65)的结合序列:5’-GGG(A/G)(C/A/T)T(C/T)-3’)。 一般来说,NF-kappaB的激活促进细胞生长,许多抗肿瘤药物以NF-kappaB为靶点,有兴趣的战友可以找些相关文献来看看。然而,某些情况下NF-kappaB的激活却有诱导凋亡的作用。
本信号转导涉及的信号分子主要包括: BCR,TCR,TLRs,IL-1R,TNFR,GF-Rs,LTβR,CD40,BR3,MyD88,IRAK1/4,TRAF2,TRAF3,TRAF5,TRAF6,Ubc13,UEV1A,TRADD,RIP,CYLD,Pellino,TAB1,TAB2,TAB3,TAK1,A20,ITCH,TAX1BP1,PI3K,PDK1,NIK,NEMO,ELKS,IKKα,IKKβ,IKKγ,ELKS,Tax,β-TrCP,P50,P52,P65,P100,P300,RelA,RelB,cRel,NF-κB,NF-κBα,NF-κBβ,NF-κBε, IκBα,IκBε,IκBζ,NF-κB1,NF-κB2,NAP1,NAK,
P I3K/A K T信号通路磷脂酰肌醇3-激酶(PI3Ks)信号参与增殖、分化、凋亡和葡萄糖转运等多种细胞功能的调节.近年来发现,IA型PI3K和其下游分子蛋白激酶B(PKB或Akt)所组成的信号通路与人类肿瘤的发生发展密切相关.该通路调节肿瘤细胞的增殖和存活,其活性异常不仅能导致细胞恶性转化,而且与肿瘤细胞的迁移、黏附、肿瘤血管生成以及细胞外基质的降解等相关,目前以PI3K-Akt信号通路关键分子为靶点的肿瘤治疗策略正在发展中. 在PI3K家族中,研究最广泛的是能被细胞表面受体所激活的I型PI3K.哺乳动物细胞中Ι型PI3K又分为IA和IB两个亚型,他们分别从酪氨酸激酶连接受体和G蛋白连接受体传递信号.IA型PI3K是由催化亚单位p110和调节亚单位p85所组成的二聚体蛋白,具有类脂激酶和蛋白激酶的双重活性.PI3K通过两种方式激活,一种是与具有磷酸化酪氨酸残基的生长因子受体或连接蛋白相互作用,引起二聚体构象改变而被激活;另一种是通过Ras和p110直接结合导致PI3K的活化.PI3K激活的结果是在质膜上产生第二信使PIP3,PIP3与细胞内含有PH结构域的信号蛋白Akt和 PDK1(phosphoinositidedependentkinase-1)结合,促使PDK1磷酸化Akt蛋白的Ser308导致Akt的活化.Akt还能通过PDK2(如整合素连接激酶ILK)对其Thr473的磷酸化而被激活.活化的Akt通过磷酸化作用激活或抑制其下游靶蛋白Bad、Caspase9、NF-κB、GSK-3、FKHR、p21Cip1和p27Kip1等,进而调节细胞的增殖、分化、凋亡以及迁移等. PI3K-Akt信号通路的活性被类脂磷酸酶 PTEN(phosphataseandtensinhomologdeletedonchromosometen)和 SHIP(SH2-containinginositol5-phosphatase)负调节,他们分别从PIP3的3′和5′去除磷酸而将其转变成PI(4,5)P2和PI(3,4)P2而降解.迄今为止,尚未发现下调Akt活性的特异磷酸酶,但用磷酸酶抑制剂处理细胞后,发现Akt的磷酸化和活性均有所增加.最近发现Akt能被一种C末端调节蛋白(CTMP)所失活,CTMP能结合Akt并通过抑制Akt 的磷酸化而阻断下游信号的传递,CTMP的过表达能够逆转v-Akt转化细胞的表型.热休克蛋白90(HSP90)亦能结合Akt,阻止Akt被PP2A磷酸酶的去磷酸化而失活,因此具有保护Akt的作用. 本信号转导涉及的信号分子主要包括 Integrin,FAK,Paxillin,ILK,PIP3,S6,p70S6K,RTK,Gab1,Gab2,IRS-1,PI3K,PTEN,AKT,PDK1,CytokineReceptor,Jak1,CD19,BCR,Ag,BCAP,Syk,Lyn,GPCR,TSC1,TSC2,Gβγ,GαGTP,PP2A,PHLPP,CTMP,PDCD4,4E-BP1,ATG13,mTORC1,TSC1,TSC2,PRAS40,XIAP,FoxO1,Bim,Bcl-2,Bax,MDM2,p53,Bax,Bad,14-3-3,Wee1,Myt1,p27Kip1,p21Waf1/Cip1,CyclinD1,GSK-3,GS,Bcl-2,mTORC2,LaminA,Tpl2,IKKα,eNOS,GABAAR,Huntingtin,Ataxin-1,PFKFB2,PIP5K,AS160等。