信号通路9—MAPK Signaling
APExBIO
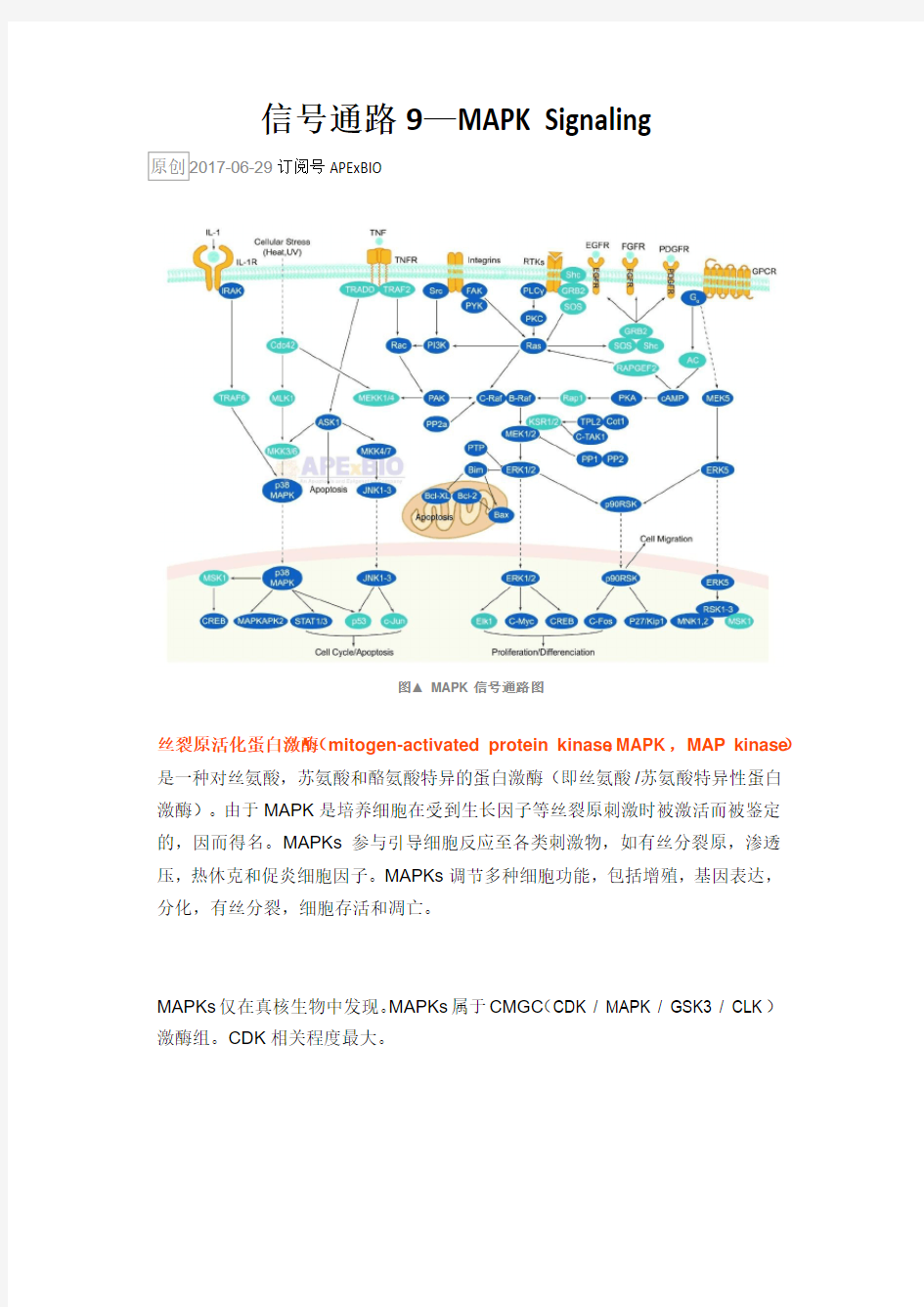
图▲ MAPK信号通路图
丝裂原活化蛋白激酶(mitogen-activated protein kinase,MAPK, MAP kinase)是一种对丝氨酸,苏氨酸和酪氨酸特异的蛋白激酶(即丝氨酸/苏氨酸特异性蛋白激酶)。由于MAPK是培养细胞在受到生长因子等丝裂原刺激时被激活而被鉴定的,因而得名。MAPKs参与引导细胞反应至各类刺激物,如有丝分裂原,渗透压,热休克和促炎细胞因子。MAPKs调节多种细胞功能,包括增殖,基因表达,分化,有丝分裂,细胞存活和凋亡。
MAPKs仅在真核生物中发现。MAPKs属于CMGC(CDK / MAPK / GSK3 / CLK)激酶组。CDK相关程度最大。
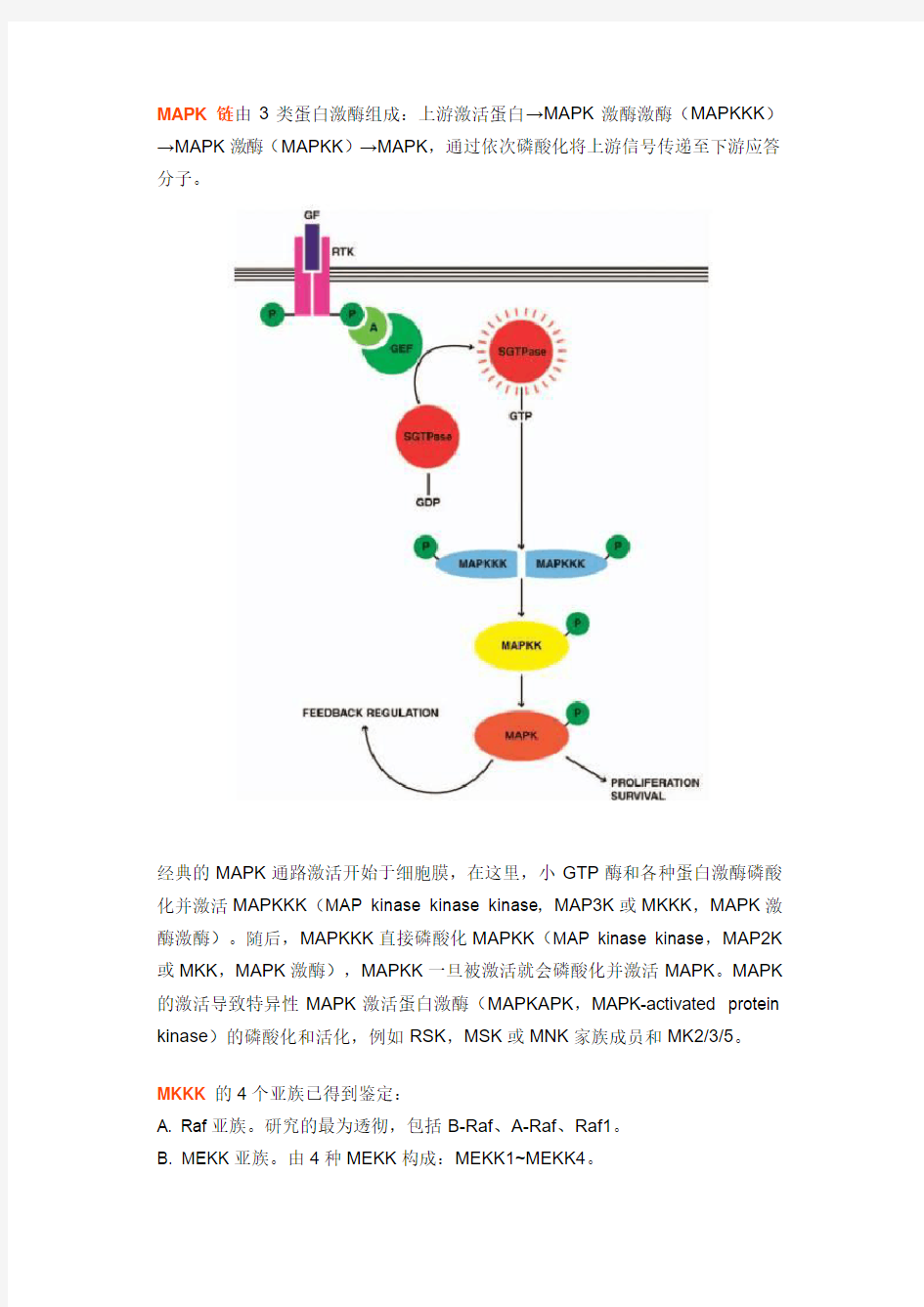
MAPK链由3类蛋白激酶组成:上游激活蛋白→MAPK激酶激酶(MAPKKK)→MAPK激酶(MAPKK)→MAPK,通过依次磷酸化将上游信号传递至下游应答分子。
经典的MAPK通路激活开始于细胞膜,在这里,小GTP酶和各种蛋白激酶磷酸化并激活MAPKKK(MAP kinase kinase kinase,MAP3K或MKKK,MAPK激酶激酶)。随后,MAPKKK直接磷酸化MAPKK(MAP kinase kinase,MAP2K 或MKK,MAPK激酶),MAPKK一旦被激活就会磷酸化并激活MAPK。MAPK 的激活导致特异性MAPK激活蛋白激酶(MAPKAPK,MAPK-activated protein kinase)的磷酸化和活化,例如RSK,MSK或MNK家族成员和MK2/3/5。
MKKK的4个亚族已得到鉴定:
A. Raf亚族。研究的最为透彻,包括B-Raf、A-Raf、Raf1。
B. MEKK亚族。由4种MEKK构成:MEKK1~MEKK4。
C. 第三个亚族:ASK1和Tpl2。
D. 第四个亚族与上述三个有较大不同,它包括MST(mammalian sterile 20-like)、SPRK、MUK(MAPK upstream kinase)、TAK1,以及相关程度最小的MOS (molony sarcoma oncoprotein)。
MKK包括:
MAP2K1 (MEK1),MAP2K2 (MEK2),
MAP2K3 (MKK3),MAP2K4 (MKK4),
MAP2K5 (MKK5),MAP2K6 (MKK6),
MAP2K7 (MKK7)。
MEK1与MEK2密切相关,而MKK3则与MKK6密切相关。
MAPK包括 ERK,JNK和p38激酶。这些MAPK通过在三肽基序Thr-Xaa-Tyr 处的双磷酸化而被激活。这三肽基序的序列在每组MAPKs中是不同的:ERK (Thr-Glu-Tyr);p38(Thr-Gly-Tyr);JNK(Thr-Pro-Tyr)。
目前分为6个亚组:
A. Extracellular signal-regulated kinases (ERK1, ERK2);
B. c-Jun N-terminal kinases (JNKs);
C. p38 isoforms (MAPK11/p38-β, MAPK12/p38-γ/ERK6, MAPK13/p38-δ/SAPK4, MAPK14/p38-α);
D. ERK5 (MAPK7);
E. ERK3 (MAPK6) and ERK4 (MAPK4);
F. ERK7/8 (MAPK15)。
在未受刺激的细胞内,MAPK处于静止状态。细胞受到生长因子或其他因素刺激后,MAPK接收MKK和MKKK的活化信号而被激活,表现为逐级磷酸化。哺乳动物中,ERK广泛存在于各种组织,参与细胞的增殖分化的调控。多种生长因子受体、营养相关因子受体等都需要ERK的活化来完成信号转导过程。JNK家族
是细胞对各种应激原诱导的信号转导的关键分子,参与细胞对辐射、渗透压、温度变化等应激反应。p38介导炎症、凋亡等,因而成为开发抗炎药物的靶位。
ERK1和ERK2异构体的经典活化是通过配体与受体酪氨酸激酶(receptor tyrosine kinase,RTK)在质膜(PM)结合,然后激活小G蛋白Ras引发的。反过来,Ras招募并激活丝氨酸/苏氨酸蛋白激酶Raf(一种MAP3K),其激活MAP2K,MEK,这反过来又在TEY基序中的苏氨酸和酪氨酸残基处磷酸化MAPK,ERK1 / 2。Ras / Raf / MEK / ERK1 / 2途径可以通过双特异性MAPK磷酸酶(MKP)失活。例如,MKP2 / 4去磷酸化ERK1 / 2,也可使其它MAPKs失活,MKP3和MKP-X对ERK特异。酪氨酸磷酸酶SHP2也通过激活G蛋白Ras而作用于该信号通路。ERK1 / 2途径调节IL-10的产生,其诱导Th细胞分化为Th2型。
应激和细胞因子激活p38 MAPK同工型,其在炎症反应中起重要作用,尽管各种同工型由不同基因编码并显示不同的组织表达模式。与ERK同工型一样,p38 MAPK也被依次激活。当响应于应激或细胞因子时,发生典型的激活,一种MAP3K 如MEKK1,ASK1或TAK1被TRAF (TNF受体相关因子) 2/3/6或Rho蛋白激活。反过来,MAP3K磷酸化MAP2K(MKK3或MKK6),然后磷酸化p38同工型的TGY基序。p38 MAPKs α,β,δ和γ被几种双特异性蛋白磷酸酶(如MPK2 / 4)去磷酸化,也可以使ERK失活。磷酸酶MPK5 / 7也可以去磷酸化JNK 和p38,而MPK1对p38表现出更高的特异性。IL-12的产生由p38MAPK调节,参与Th1应答的诱导。
在哺乳动物中编码JNK的三个基因:Jnk1,Jnk2和Jnk3,其中Jnk1和Jnk2在体内广泛表达,而Jnk3在脑,心脏和睾丸中表达。JNKs被许多应激物激活,一旦激活,JNK从细胞质转移到细胞核。JNK的下游靶点包括转录因子c-Jun,其在JNK介导的磷酸化后转位到细胞核。JNK在c-Jun N-末端磷酸化丝氨酸残基63和73,从而激活c-Jun并增强其转录活性。c-Jun调节促凋亡或抗凋亡基因Bax和Bcl-2的表达。