染色体互换规律
- 格式:docx
- 大小:25.45 KB
- 文档页数:11


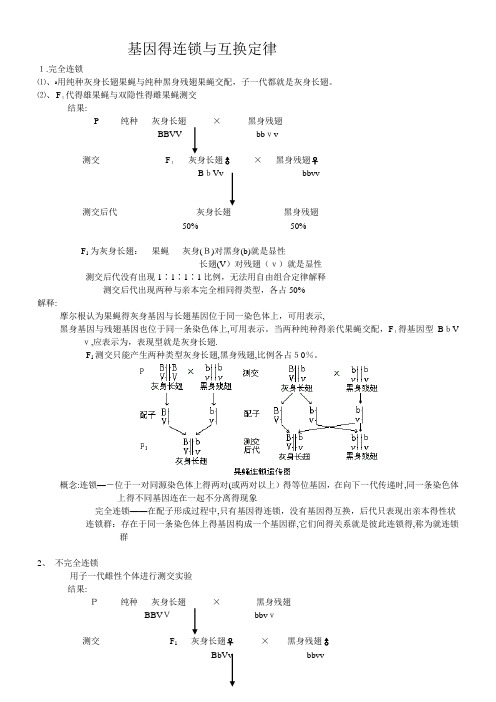
基因得连锁与互换定律1.完全连锁⑴、ﻩ用纯种灰身长翅果蝇与纯种黑身残翅果蝇交配,子一代都就是灰身长翅。
⑵、 F1代得雄果蝇与双隐性得雌果蝇测交结果:P 纯种灰身长翅×黑身残翅BBVV bbvv测交F1灰身长翅♂×黑身残翅♀BbVv bbvv测交后代灰身长翅黑身残翅50%50%F1为灰身长翅:果蝇灰身(B)对黑身(b)就是显性长翅(V)对残翅(v)就是显性测交后代没有出现1∶1∶1∶1比例,无法用自由组合定律解释测交后代出现两种与亲本完全相同得类型,各占50%解释:摩尔根认为果蝇得灰身基因与长翅基因位于同一染色体上,可用表示,黑身基因与残翅基因也位于同一条染色体上,可用表示。
当两种纯种得亲代果蝇交配,F1得基因型BbV v,应表示为,表现型就是灰身长翅.F1测交只能产生两种类型灰身长翅,黑身残翅,比例各占50%。
概念:连锁—-位于一对同源染色体上得两对(或两对以上)得等位基因,在向下一代传递时,同一条染色体上得不同基因连在一起不分离得现象完全连锁——在配子形成过程中,只有基因得连锁,没有基因得互换,后代只表现出亲本得性状连锁群:存在于同一条染色体上得基因构成一个基因群,它们间得关系就是彼此连锁得,称为就连锁群2、不完全连锁用子一代雌性个体进行测交实验结果:P纯种灰身长翅×黑身残翅BBVVbbvv测交F1灰身长翅♀×黑身残翅♂BbVv bbvv测交后代灰身长翅黑身残翅42%42%灰身残翅黑身长翅8% 8%后代出现四种性状,其中亲本类型占多数,新组合类型占少数.解释:细胞在进行减数分裂形成配子得过程中,减数分裂第一期前期,同源染色体联会,形成四分体。
联会复合体中同源染色体间得非姐妹染色单体间会发生染色单体得交叉互换,在交换区段上得基因随染色体发生交换,这种交换产生新得基因组合。
交叉互换后形成四种配子,其数量相同;其中有两种配子就是亲本类型,两种配子就是重组合类型,各占一半。

基因的连锁互换定律1. 引言基因连锁互换定律是遗传学中的一个重要原理,它描述了基因在染色体上的相对位置以及它们之间的遗传关系。
该定律的提出和发展对于理解基因的遗传方式以及基因的遗传变异具有重要意义。
本文将详细介绍基因连锁互换定律的背景、原理、实验方法以及应用领域。
2. 背景基因连锁互换定律最早由托马斯·亨特·摩尔根于1911年提出。
当时,他研究了果蝇的眼色突变,并发现某些突变基因总是以固定的方式出现,即它们似乎被”连锁”在一起,遗传方式与经典的孟德尔遗传规律不同。
这一发现引发了他对基因连锁互换的研究,并最终推导出了基因连锁互换定律。
3. 原理基因连锁互换定律基于遗传连锁现象,即位于同一染色体上的基因倾向于一起遗传。
这是因为染色体在有丝分裂过程中的交叉互换现象导致的。
交叉互换是指染色体上的两条同源染色单体(同一对同源染色体)发生物理上的交换,从而导致基因重组。
交叉互换的发生是在减数分裂过程中。
在减数分裂的第一次分裂中,同源染色单体之间发生交叉互换,将某些片段的基因从一个染色体转移到另一个染色体上。
这种交叉互换的结果是,原本连锁在一起的基因在染色体上被打破,重新组合成新的连锁方式。
基因连锁互换定律的原理可以用以下公式表示:距离 = 互换事件数 / 总体数 * 100%。
其中,距离表示两个基因之间的距离,互换事件数表示两个基因在交叉互换中发生重组的次数,总体数表示总的遗传体数。
4. 实验方法为了验证基因连锁互换定律,科学家们进行了一系列经典的果蝇实验。
这些实验通常包括交配、观察后代的表型以及计算连锁距离等步骤。
首先,选择具有不同表型的果蝇进行交配。
例如,选择一个拥有红眼睛和长翅膀的果蝇和一个拥有白眼睛和短翅膀的果蝇进行交配。
然后观察后代果蝇的表型,记录各种表型的果蝇数量。
接下来,根据后代果蝇的表型比例计算连锁距离。
连锁距离可以通过以下公式计算:连锁距离 = 重组类型数 / 总体数 * 100%。

1913年谁根据经营的连锁互换规律绘制了世界上第一张染色体连孟德尔定律的“例外”——连锁和互换遗传现象的阐释摩·尔根证明了基因位于染色体上。
但一种生物的基因数目远远多于染色体的数目,因而一条染色体上存在着多个基因,就成为一个必然的推论。
早在1906年,遗传学的早期倡导者、英国遗传学家贝特森和他的学生庞尼特在用香豌豆进行的杂交实验中,就发现了生物性状的连锁遗传现象,但当时他们无法对此做出正确的解释。
1912年在进行两对相对性状的杂交试验时,摩·尔根和他的学生们用纯种灰身长翅果蝇与纯种黑身残翅果蝇交配,他们看到子一代(F1)都是灰身长翅的,由此可以推出,果蝇的灰身(B)对黑身(b)是显性;长翅(V)对残翅(v)是显性。
所以,纯种灰身长翅果蝇的基因型与纯种黑身残翅果蝇的基因型应该分别是(BBVV)和(bbvv),F1的基因型应该是(BbVv)。
摩·尔根又让F1的雄果蝇(BbVv)与双隐性类型的雌果蝇(bbvv)测交,按照自由组合定律,测交后代中应该出现4种不同的类型,即灰身长翅、灰身残翅、黑身长翅、黑身残翅,并且它们之间的数量比应该为1:1:1:1。
但是,测交的结果与原来预测的完全不同,只出现两种和亲本完全相同的类型:灰身长翅(BbVv)和黑身残翅(bbvv),并且两者的数量各占50%。
很明显,这个测交的结果是无法用基因的自由组合定律来解释的。
为什么会出现上述试验结果呢?摩·尔根认为果蝇的灰身基因和长翅基因位于同条染色体上;黑身基因和残翅基因也位于同一条染色体上。
所以,当两种纯种的亲代果蝇交配后,F1的基因型BbVv,应该表示为,表现型是灰身长翅。
这样,在F1雄果蝇产生配子时,原来位于同一条染色体上的两个基因(B和V、b和v)就不能分离,而是连在一起向后代传递。
因此,当F1雄果蝇与黑身残翅的雌果蝇交配后,只能产生灰身长翅(BbVv)和黑身残翅(bbvv)两种类型,并且这两者的数量各占50%。

第五章连锁与互换规律1910年摩尔根将果蝇白眼基因定位于X上后,又发现一些伴性遗传基因,证明X上确有许多基因(人有23对染色体和十万个基因)。
提出问题:(1)同一染色体上众多基因如何排列(2)遗传传递有何规律经研究得到结论:(1)基因论(2)连锁交换规律第一节基因的连锁与交换一、连锁遗传现象(一)连锁遗传现象发现1906年英国学者贝特森(Bateson)和潘耐特(Pannett)研究香豌豆两对性状遗传时,首先发现的。
分离比不符花颜色紫色P对红色p显性,花粉粒形状长形L对圆形l显性。
F2合9:3:3:1,亲组合类型较多,重组合类型偏少。
原为同一亲本的两个性状,在F中常常有联系在一起的倾向,这说明来自同2一亲本的基因,有较多的在一起传递的可能。
但贝特森和潘耐特未能提出科学的产生配子数找原因,4种配子数目不等,亲组合类型多,解释。
摩尔根认为应从F1重组类型少。
(二)摩尔根的实验果蝇翅的长短,复眼的颜色长翅Vg,残翅vg,红色复眼Pr,紫色复眼pr解释相引组杂交红眼雄果蝇产生两种数目相同的配子,测交后代1:1,红眼雌果蝇产生四种配子,但亲组型配子Vg Pr和vg pr多,而重组型配子Vg pr和vgPr少。
同理可解释相斥组杂交结果。
果蝇翅的长短和眼的颜色是两对相对性状,这两对相对性状在杂交后代中具有某种程度的相关性或相连性,把这种现象叫连锁。
把这两对相对性状叫连锁性状。
控制连锁性状的基因叫连锁基因,连锁基因位于同一对同源染色体上。
连锁基因之间能够发生交换的连锁叫不完全连锁(雌果蝇),不能发生交换的连锁叫完全连锁(雄果蝇、雌家蚕)。
目前人们经过研究发现仅有两种动物雄果蝇和雌蚕属于完全连锁,不发生交换。
相引组:两个显性在一起,两个隐性在一起配成的杂交组合。
相斥组:一个显性一个隐性和一个隐性一个显性在一起配成的杂交组合。
(三)连锁的细胞学基础同一染色体上非等位基因随染色体一起遗传——完全连锁,又能按一定比例发生交换而重组——不完全连锁。
专题二十六基因的连锁和交换定律【学习内容】1、基因的连锁和交换【课堂笔记】一、完全连锁现象:F1灰身长翅♂(BbVv)×黑身残翅♂(bbvv) →1灰身长翅(BbVv):1黑身残翅(bbvv)原因:(只有亲本类型,没有重组类型)(♂)−−−→−减数分裂1:1F1雄果蝇产配子时,同一条染色体上的不同基因常连在一起不分离,所以只有亲本类型的配子没有重组类型的配子。
二、不完全连锁现象:F1灰身长翅雌(BbVv)×黑身残翅雄(bbvv)↓灰长(BbVv):黑残(bbvv):灰残(Bbvv):黑长(bbVv)42 :42 :8 :8(亲本类型特别多,重组类型特别少)原因:B V B V b v B v b V(♀) ::F1雌果蝇产配子时,部分初级卵母细胞的同一条染色体上不同基因因同源染色体的非姐妹染色单体之间的交叉互换,即产生亲本类型的配子又产生了重组类型的配子。
三、三者比较图B vb vB v b v42 42 8 8四、交换值的计算1、发生互换的初级性母细胞的比例:如果一个生物体中有20%的初级性母细胞发生了互换,那么发生互换的初级性母细胞比例就是20%。
发生互换的初级性母细胞比例= 发生互换的初级性母细胞所有初级性母细胞2、交换值:重组型配子占总配子的比例(1)交换值= 重组型配子总配子(2)交换值=发生互换的初级性母细胞比例/2(3)交换值=某种重组型配子所占的比例*2【运用练习一】1、基因的连锁和交换定律是由哪一位科学家发现的()A.孟德尔B.艾弗里C.摩尔根D.达尔文2、100个精母细胞在减数分裂中,有40个细胞的染色体发生了一次交换,在所形成的配子中,重组配子占()A.5%B.10%C.20%D.40%3、有一种红花大叶的植株与白花小叶的植株杂交,其测交后代得到红花大叶370株,白花小叶342株。
那么该植株上述性状的遗传遵循的规律是()A.分离规律B.自由组合规律C.完全连锁遗传D.不完全连锁遗传4、杂合体AaBb若完全连锁遗传经减数分裂产生的配子可能有几种()A.一种B.两种C.三种D.四种5、生物体通过减数分裂形成配子时,基因的交换发生在()A.一条染色体的姐妹染色单体之间B.两条非同源染色体之间C.一对同源染色体的非姐妹染色单体之间D.两对同源染色体之间6、杂合体AaBb经过减数分裂产生了4种类型的配子AB、Ab、aB、ab,其中AB和ab两种配子各占42%。
[键入文字]高一生物知识点:基因的连锁和交换定律高一生物为我们展示了一个丰富多彩的生物界,是一门十分有意思的学科。
高一生物的学习需要将所有知识点进行总结,方便大家集中记忆。
但是如何进行总结是摆在同学们面前的一个难题,下面小编为大家提供高一生物知识点:基因的连锁和交换定律,供大家参考,希望对大家学习有帮助。
基因的连锁和交换定律 名词:1、基因的连锁:位于同一条染色体上的不同基因在减数分裂过程形成配子时,常常连在一起不相分离,进入配子,这种现象,我们把它叫做基因的连锁。
2、基因的互换:位于同一条染色体上的不同基因,在减数分裂的四分体时期,由于同源染色体上的等位基因随着非姐妹染色体单体的交换而发生互换的现象,我们称之为基因的互换。
3、不完全连锁遗传:像这种雌果蝇的遗传,基因既有连锁,又有互换的现象。
4、完全连锁:雄果蝇的遗传只有连锁,没有互换。
语句:1、完全连锁的实例:用果蝇做杂交实验:纯种的灰身长翅与黑身残翅杂交,F1代为灰身长翅,所以,灰身长翅为显性,黑身残翅为隐性,对F1代中的雄性个体测交,测交后代的表现型是1灰身长翅:1黑身残翅,与F1代完全相同。
2、不完全连锁杂交实例:选择F1中的雌性BbVv测交:BbVvXbbvv42%BbVv:42%bbvv:8%Bbvv:8%bbVv。
2、比较完全连锁与不完全连锁的异同。
(1)相同点:二组杂交的亲代与F1代情况相同。
(2)不同点:完全连锁的测交后代只有两种基因型,与亲本相同,数量比1:1。
不完全连锁的测交后代有四种基因型,其中亲本基因型(与其亲本相同的基因型)各占42%,重组基因型(与其亲本不同的基因型)各占8%。
3、(1)自由组合是分1。
基因的连锁互换定律摘要:一、基因连锁互换定律的概念1.基因的连锁与互换定律的定义2.连锁与互换定律的实质二、基因连锁互换定律的发现与发展1.摩尔根及学生的实验2.连锁与互换定律的证明三、基因连锁互换定律的应用1.育种工作中的应用2.基因治疗中的连锁互换四、基因连锁互换定律与人类遗传病1.遗传病的分类2.基因连锁互换定律与遗传病的关系正文:基因的连锁互换定律是遗传学中的一个重要定律,它揭示了在进行减数分裂形成配子时,位于同一条染色体上的不同基因,常常连在一起进入配子;在减数分裂形成四分体时,位于同源染色体上的等位基因有时会随着非姐妹染色单体的交换而发生交换,因而产生了基因的重组。
基因的连锁互换定律的发现和发展离不开摩尔根及其学生的实验。
他们利用果蝇进行的杂交实验,揭示了位于同源染色体上不同座位的两对以上等位基因的遗传规律,即著名的连锁与互换规律。
基因的连锁互换定律在育种工作中有着广泛的应用。
例如,大麦抗秆锈病与抗散黑穗病的基因就是紧密连锁的,只要选择大麦抗秆锈病的植株,也就等于同时选择了抗散黑穗病的植株,达到一举两得、提高选择效率的目的。
在基因治疗中,基因的连锁互换定律也发挥着重要作用。
通过基因交换,可以让人所需要的基因重组在一起,从而培育出优良品种来。
例如,有两个大麦品种:一个是矮秆抗倒伏但不抗锈病的品种,另一个是高秆易倒伏但抗锈病的品种。
每一个品种中控制这两个性状的基因都位于同一条染色体上。
经过杂交,F2 会出现四种类型的后代,其中由于基因交换而出现的矮秆抗倒伏同时又抗锈病的类型就是符合需要的类型,经过进一步培育和大量繁殖就可以成为新的优良品种。
当然,基因的连锁互换定律与人类遗传病也息息相关。
遗传病可分为单基因遗传病、多基因遗传病和染色体异常遗传病。
第1篇一、引言基因是生物体内决定个体性状的基本单位,它们通过遗传的方式传递给后代。
在遗传学的研究过程中,科学家们发现了一种特殊的遗传现象——基因的连锁互换。
这一现象揭示了基因之间的相互作用和调控机制,为理解生物体的遗传规律提供了重要线索。
本文将围绕基因的连锁互换展开论述,探讨其基本原理、研究方法及其在生物学和医学领域的应用。
二、基因连锁互换的基本原理1. 基因连锁基因连锁是指位于同一染色体上的基因在遗传过程中倾向于一起传递给后代。
这是由于染色体在减数分裂过程中发生交叉互换,导致连锁基因的重组。
基因连锁现象最早由孟德尔发现,并提出了基因分离定律和自由组合定律。
2. 基因互换基因互换是指在连锁的基因之间发生交换,导致基因重组。
基因互换可以通过以下两种方式实现:(1)同源重组:在减数分裂过程中,同源染色体上的同源片段发生交换,导致连锁基因重组。
(2)非同源重组:在减数分裂过程中,非同源染色体上的非同源片段发生交换,导致连锁基因重组。
三、基因连锁互换的研究方法1. 育种实验育种实验是研究基因连锁互换的重要方法。
通过杂交、自交、回交等手段,观察和分析基因连锁互换现象,从而揭示基因之间的相互作用。
2. 基因图谱构建基因图谱构建是研究基因连锁互换的基础。
通过基因测序、分子标记等技术,确定基因在染色体上的位置,构建基因图谱。
3. 生物信息学分析生物信息学分析是研究基因连锁互换的重要手段。
通过比较基因组、转录组、蛋白质组等数据,揭示基因之间的相互作用和调控机制。
四、基因连锁互换在生物学和医学领域的应用1. 生物学领域(1)揭示基因调控机制:基因连锁互换研究有助于揭示基因之间的相互作用和调控机制,为理解生物体的遗传规律提供重要线索。
(2)基因定位:通过基因连锁互换研究,可以确定基因在染色体上的位置,为基因克隆和功能研究提供依据。
2. 医学领域(1)遗传病诊断:基因连锁互换研究有助于发现遗传病相关基因,为遗传病诊断和治疗提供依据。
染色体互换规律 首先要明确的是有丝分裂过程同源染色体一般不会交叉换。只有减数分裂才会。 减数分裂,交换以后,先同源染色体分离,减数第一次分裂结束,减数第二次分裂着丝点一分为二。 有丝分裂偶然也会交叉互换,其实你可以这样理解:交叉互换跟交叉互换,其他的规律没有变化。 !需要注意的是: 有丝分裂后期姐妹染色单体分开,向两极移动,同源染色体没有分开,你仔细观察有丝分裂后期的图,在移向细胞两极的每侧染色体中你都可找到形态大小两两一致的染色体,它们就是同源染色体(注意不是两极彼此之间的,那是由姐妹染色单体分开的子染色体)。 我们继续了解一下有丝分裂: 比如:1、有丝分裂后期,姐妹染色体以分离,为什么还会有同源染色体? 因为同源染色体是形态大小相同,它分别来自父方和母方的一对染色体.有丝分裂后期姐妹染色体分离,但是分离后的染色体依然符合同源染色体的定义.所以同源染色体依然存在. 2、有丝分裂.减数分裂 同源染色体的行为分别是什么? 在有丝分裂过程中有同源染色体,但是同源染色体没有什么具体的变化,就是在有丝分裂后期姐妹染色单体分开,形成新的染色体,末期分裂成两个细胞染色体数恢复到正常值。 减数分裂过程中只有减数第一次分裂有同源染色体的变化,在减数第二次分裂没有同源染色体。在减数第一次分裂中,同源染色体是先联会形成四分体(此时可以发生同源染色体上非姐妹染色单体的交叉互换)接着同源染色体排列在赤道板两侧,后期的时候同源染色体向两极移动,末期形成同源染色体分离分别进入两个子细胞中。 同源染色体的相关知识 定义1:形态相同、在减数分裂中能配对的两条染色体 定义2:二倍体细胞中染色体以成对的方式存在, 一条来自父本,一条来自母本,且形态、大小相同,并在减数分裂前期相互配对的染色体。含相似的遗传信息。 定义3:一条来自父本,一条来自母本,且形态、大小相同,在减数分裂前期相互配对的染色体。是在二倍体生物细胞中,形态、结构基本相同的染色体,并在减数第一次分裂(参考减数分裂)的四分体时期中彼此联会(若是三倍体及其他奇数倍体生物细胞,联会时会发生紊乱),最后分开到不同的生殖细胞(即精子、卵细胞)的一对染色体,在这一对染色体中一个来自母方,另一个来自父方。 同源染色体(homologous chromosomes)是有丝分裂中期看到的长度和着丝点位置相同的两个染色体,或减数分裂时看到的两两配对的染色体。同源染色体一个来自父本,一个来自母本;它们的形态、大小和结构相同。由于每种生物染色体的数目是一定的,所以它们的同源染色体的对数也一定。例如豌豆有14条染色体,7对同源染色体。同源染色体上常含有不同的等位基因,减数分裂时又进行了交换并随机地分配到不同的性细胞中去,这对于遗传重组有重要意义。 正常细胞中,一对同源染色体若表示为由两条染色体A、B组成,在减数分裂中变成“AA`”组和“BB`”组,两组联会形成四分体;有丝分裂中变成“AA`”组和“BB`”组,但两组不联会,在有丝分裂中期染色体的着丝点整齐排列在赤道板上,着丝点分开后染色体平均分配到两个子细胞。 在生物体的有性生殖过程中,有性生殖细胞是通过细胞分裂的一种——减数分裂形成的。在减数分裂的分裂间期,精原细胞的体积略微增大,染色体进行复制,成为初级精母细胞。复制后的每条染色体都含有两条姐妹染色体,这两条姐妹染色单体并列在一起,由同一个着丝点连接着。分裂期开始后不久,初级精母细胞中原来分散存在的染色体进行配对。而在减数第二次分裂过程中不存在同源染色体。 区分同源染色体与姐妹染色单体:姐妹染色单体是由一个着丝点连着的并行的两条染色单体,是在细胞分裂的间期由同一条染色体经复制后形成的——由一条染色体复制形成的两条子染色体不是同源染色体,因为它们尽管形状大小相同,但它们并非一条来自父方、一条来自母方。 不仅只有减数分裂有同源染色体的存在,有丝分裂也有。所有的只要有一对染色体存在的都会存在。 举例:同源染色体分离,某细胞复制后染色体为以下AAA*A*BBB*B*,因为某些原因把它分解成这样,AA与A*A*为相同基因.我们看看他的分离过程变化。 在减数第一次分裂前期,同源染色体联会,形成四分体(因其含有四条染色单体而得名)。到了中期,四分体排列在赤道板上(着丝点位于同一平面内)。进入后期,中心体发出的纺锤丝牵引着四分体中的一对同源染色体,使它们分别移向细胞两极,这样就完成了一对同源染色体的分离。 例如上述细胞复制,AAaaBBbb分离,AA和aa必须分离,BB和bb也必须分离,但是至于是AA与BB进入一个细胞还是AA与bb或aa与BB以及aa与bb进入一个细胞就是随机的了。比如得到的次级性母细胞基因型可以是AABB、AAbb、aaBB 、aabb。 1、同源染色体在减数分裂中分离共几次? 就一次。是在减数第一次分裂的后期。之后就没有同源染色体了,开始减数第二次分裂。减数第二次分裂后期是姐妹染色单体分离。 2、同源染色体交叉互换 正常发生,属于基因重组的一种类型,应该是频繁发生 四分体时期非姐妹染色单体之间的交叉互换,是指等位基因的交换,属于基因重组。如一条为AB的染色体和一条ab的染色体在交换后可得到Ab及aB两条新基因型的染色体,基因发生重新组合。 减数分裂 减数分裂是生物细胞中染色体数目减半的分裂方式。性细胞分裂时,染色体只复制一次,细胞连续分裂两次,这是染色体数目减半的一种特殊分裂方式。减数分裂不仅是保证物种染色体数目稳定的机制,同时也是物种适应环境变化不断进化的机制。在减数分裂过程中,染色体只复制一次,而细胞分裂两次。减数分裂的结果是:成熟生殖细胞中的染色体数目比原始生殖细胞的减少一半。减数分裂(Meiosis) 范围是进行有性生殖的生物;时期是从原始生殖细胞发展到成熟生殖细胞。 分裂过程: 注:减数分裂可以分为两个阶段,间期和分裂期,其中分裂期又分为减数第一次分裂期(减一),减数第二次分裂期(减二)。在高中知识范围内,减一的末期和减二的前期可以看作同一个时期,我们一般将其称为减一的末期。(减一末期与减二前期间有间期但很短可以忽略)
1.细胞分裂前的间期,进行DNA和染色体的复制,染色体数目不变,DNA数目变为原细胞的两倍。 2.减一前期同源染色体联会.形成四分体(或“二联体”),核仁核膜消失。
3.减一中期.出现纺锤体,同源染色体着丝点对称排列在赤道板两端。(与动物细胞的有丝分裂大致相同,动物细胞有丝分裂为着丝点排列在赤道板上)
4.减一后期,同源染色体分离,非同源染色体自由组合,移向细胞两极。
5.减一末期细胞一分为二,形成次级精母细胞或形成次级卵母细胞和第一极体。 6.减二前期次级精母细胞中染色体再次聚集,再次形成纺锤体。
7.减二中期染色体着丝点排在赤道板上。 8.减二后期染色体着丝点分离,染色体移向两极。 9.减二末期,细胞一分为二,精原细胞形成精细胞,卵原细胞形成卵细胞和第二极体。
减数第一次分裂 间期 可以分为三个阶段:G1期、S期、G2期。根据现代细胞生物学的研究,细胞分裂的间期分为三个阶段:第一间隙期,称为G1期;合成期,称为S期;第二间隙期,称为G2期。其中G1和G2期主要是合成有关蛋白质和RNA,S期则完成DNA 的复制。 G1期 G1期的特点:G1期是从上次细胞增殖周期完成以后开始的。G1期是一个生长期。在这一时期主要进行RNA和蛋白质的生物合成,并且为下阶段S期的DNA合成做准备。如合成各种与DNA复制有关的酶,线粒体、核糖体等都增多了,内质网在更新扩大,来自内质网的高尔基体、溶酶体等也增加了。动物细胞的2个中心粒也彼此分离并开始复制。也就是说为S期储备物质和能量。 S期 S期的特点:从G1期进入S期是细胞增殖的关键。S期最主要的特征是DNA的合成,DNA分子的复制就是在这个时期进行的。通常只要DNA的合成一开始,细胞增殖活动就会进行下去,直到分裂成两个子细胞。 G2期 G2期的特点:G2期又叫做“有丝分裂的准备期”因为它主要为后面的分裂期(M期)做准备。在G2期中,DNA的合成终止,但是还有RNA和蛋白质的合成,不过合成量逐渐减少。特别是微管蛋白的合成,为分裂期(M期)纺锤体微管的组装提供原料。在G2期中心粒完成复制而成2对中心粒。 前期 根据染色体的形态,可分为5个阶段: 〖细线期〗 细胞核内出现细长、线状染色体,细胞核和核仁体积增大。每条染色体含有两条姐妹染色单体。 〖偶线期〗 又称配对期。细胞内的同源染色体两两侧面紧密相进行配对,这一现象称作联会。由于配对的一对同源染色体中有4条染色单体,称为四分体(或“二联体”) 〖粗线期〗 染色体连续缩短变粗,同时,四分体中的非姐妹染色单体之间发生了DNA的片断交换,从而导致了父母基因的互换,产生了基因重组,但每个染色单体上仍都具有完全相同的基因。 〖双线期〗发生交叉的染色单体开始分开。由于交叉常常不止发生在一个位点,因此,染色体呈现V、X、8、O等各种形状。 〖终变期〗(又叫浓缩期) 染色体变成紧密凝集状态并向核的周围靠近。以后,核膜、核仁消失,最后形成纺锤体。 中期 各成对的同源染色体双双移向细胞中央的赤道板,着丝点成对排列在赤道板两侧,细胞质中形成纺锤体。