脂筏的结构与功能
- 格式:pdf
- 大小:151.47 KB
- 文档页数:6

名词解释Cell Biology:广泛采用现代生物学的实验技术和手段,应用分析和综合的方法,将细胞的整体活动水平,亚细胞水平和分子水平三方面的研究有机地结合起来,以动态的观点观察细胞和细胞器的结构和功能,以期最终阐明生命的基本规律。
脂筏(lipid raft)是质膜上富含胆固醇和鞘磷脂的微结构域(microdomain)。
大小约70nm 左右,是一种动态结构,位于质膜的外小叶。
质膜主要由膜脂和膜蛋白组成,另外还有少量糖,主要以糖脂和糖蛋白的形式存在。
膜骨架membrane associated skeleton细胞膜下与膜蛋白相连的由纤维蛋白组成的网架结构,它参与维持细胞膜的形状并协助质膜完成多种生理功能。
被动运输(passive transport):通过简单扩散或协助扩散实现物质由高浓度向低浓度方向的跨膜转运。
动力来自物质的浓度梯度,不需要细胞提供代谢能量。
简单扩散(simple diffusion)疏水的小分子或小的不带电荷的极性分子的热运动可以使分子从膜的一侧通过细胞膜到另一侧,其结果是分子沿着浓度梯度降低的方向转运。
因无需细胞提供能量,也没有膜蛋白的协助,故名。
协助扩散(facilitated diffusion)小分子物质沿其浓度梯度(或电化学梯度)减小方向的跨膜运动,是由膜转运蛋白“协助”完成的。
主动运输active transport由载体蛋白所介导的物质逆着浓度梯度或电化学梯度由低浓度侧到高浓度侧转运,需要供给能量。
ATP直接供能、间接供能、光能。
协同运输(cotransport):由离子泵与载体蛋白协同作用,利用跨膜的离子浓度梯度或电化学梯度,使特定离子的顺梯度运动与被转运分子或离子的逆梯度运输相偶联。
直接动力是膜两侧的离子浓度梯度。
胞吞作用:质膜内陷形成囊泡将外界大分子裹进并输入细胞的过程。
胞吐作用:与胞吞作用的顺序相反,将细胞内的分泌泡或其它某些膜泡中的物质通过细胞膜运出细胞的过程。

脂筏的结构,功能及在阿尔茨海默症,朊病毒病中的研究进展北京大学力学与工程科学系生物医学工程专业02级博士钟建学号: 10203829摘要:脂筏是膜脂双层内含有特殊脂质和蛋白质的微区。
主要由鞘脂,胆固醇及蛋白质组成,脂筏的组分和结构特点有利于蛋白质之间相互作用和构向转化,可以参与信号转导和蛋白质转运。
病原体如病毒,细菌及其毒素等利用脂筏进入宿主细胞,利用细胞表面的GPI-锚固蛋白和筏脂作为主要的或补充的受体,因此,很多的疾病如阿尔茨海默症,朊病毒病等均有可能与脂筏功能紊乱有着密切的关系。
关键词:脂筏,信号转导,蛋白质转运,阿尔茨海默症,朊病毒病自从1972年S. Single和G. Nicolson提出膜的流动镶嵌模型[1]以来,生物膜的研究有了飞速的发展,大量的科学家进入这一领域,推动了膜生物学的发展。
近几年来,发现大多数哺乳动物细胞质膜中具有富含胆固醇和鞘脂(鞘磷脂和鞘糖脂)的微区(microdomain),称为“脂筏(lipid raft)”[2,3]。
脂筏的组分和结构特点有利于蛋白质之间相互作用和构象转化,脂筏区主要参与信号转导和细胞蛋白转运。
脂筏曾经有过许多的名称,如糖基磷脂酰肌醇脂微区(GPI lipid microdomain),鞘糖脂富含微区(glycosphingolipid-enriched microdomain),去垢剂不溶的糖脂富含复合物(detergent-insoluble glycolipid-enriched complexes, DIGs),低浓度Triton不溶复合物(low-density triton-insoluble complex, LDTI)等等。
脂筏的组分和结构特点有利于蛋白质之间相互作用和构象转化,可以参与信号转导(Cellular signaling)和蛋白质转运(Protein trafficking)。
一些感染性疾病,心血管疾病,肿瘤,肌营养不良症,阿尔茨海默症,HIV,朊病毒病等可能与脂筏功能紊乱有着密切的关系[4]。

检测案5细胞膜与细胞核[基础巩固练]考点一细胞膜的结构与功能1.[2024·九省联考·安徽]细胞各部分结构既分工又合作,共同执行细胞的各项生命活动。
下列相关叙述正确的是()A.内质网是一个内腔不相通的膜性管道系统,与蛋白质等大分子物质的合成有关B.细胞骨架是由蛋白质组成的纤维状网架结构,与细胞运动等生命活动密切相关C.生物膜系统由细胞器膜、细胞膜和核膜等构成,具有相同的组成成分、结构和功能D.细胞核具有核膜、核仁等结构,核膜上的核孔是大分子物质被动运输进出细胞核的通道2.[2024·山西高三校联考]细胞膜流动镶嵌模型由辛格和尼科尔森提出,其结构模型如图所示。
下列有关细胞膜结构与功能的叙述,错误的是()A.物质①为糖被,精子与卵细胞进行受精时离不开物质①的识别作用B.物质③为载体蛋白,细胞膜功能的复杂程度取决于物质③的种类和数量C.细胞骨架由蛋白质纤维构成,能够支撑细胞膜,具有维持细胞形态等生理功能D.神经细胞释放氨基酸类神经递质时,需依赖于物质②和部分物质③的流动性3.[2024·沧州月考]下列关于真核细胞生物膜结构和功能的叙述,正确的是()A.构成细胞膜的脂质主要是磷脂、脂肪和胆固醇B.生物膜是细胞内所有化学反应进行的场所C.细胞膜上的受体是细胞间信息交流所必需的结构D.肌细胞的细胞膜上有协助葡萄糖跨膜运输的载体4.[2024·重庆高三统考]脂筏模型是对流动镶嵌模型的重要补充。
下图为脂筏的结构示意图,下列叙述错误的是()A.细胞膜主要是由脂质和蛋白质分子构成的B.脂筏的形成可能有助于高效地进行某些细胞代谢活动C.脂筏模型否定了“细胞膜具有流动性”D.脂筏模型中蛋白质以不同的方式分布在磷脂双分子层中5.[2024·衡水模拟]下列不属于细胞间信息交流方式的是()A.高等植物细胞之间通过胞间连丝相互连接B.胰岛B细胞分泌的胰岛素通过血液运输并作用于组织细胞C.成熟的高等植物细胞放入浓蔗糖溶液中发生质壁分离D.同种高等动物的精子和卵细胞相互接触完成受精作用考点二细胞核的结构与功能6.[2024·江西鹰潭一中校联考]下列有关真核生物细胞核的叙述,正确的是()A.核膜由4层磷脂分子组成,代谢旺盛的细胞核孔数量较多B.人体上皮细胞经碱性染料染色后,可观察到细胞核中染色体呈棒状C.细胞核是遗传信息库,是细胞的代谢中心和遗传的控制中心D.黑白两种美西螈胚胎细胞核移植实验表明,美西螈的性状取决于细胞质7.[2024·南京市高三调研]核孔结构复杂,至少由50种蛋白质构成,称为核孔复合体,该结构是核内外物质转运的通道(如下图所示)。
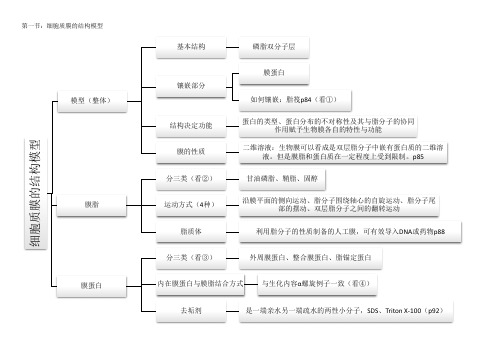
第一节:细胞质膜的结构模型细胞质膜的结构模型模型(整体)基本结构磷脂双分子层镶嵌部分膜蛋白如何镶嵌:脂筏p84(看①)结构决定功能蛋白的类型、蛋白分布的不对称性及其与脂分子的协同作用赋予生物膜各自的特性与功能膜的性质二维溶液:生物膜可以看成是双层脂分子中嵌有蛋白质的二维溶液。
但是膜脂和蛋白质在一定程度上受到限制。
p85膜脂分三类(看②)甘油磷脂、鞘脂、固醇运动方式(4种)沿膜平面的侧向运动、脂分子围绕轴心的自旋运动、脂分子尾部的摆动、双层脂分子之间的翻转运动脂质体利用脂分子的性质制备的人工膜,可有效导入DNA 或药物p88膜蛋白分三类(看③)外周膜蛋白、整合膜蛋白、脂锚定蛋白内在膜蛋白与膜脂结合方式与生化内容α螺旋例子一致(看④)去垢剂是一端亲水另一端疏水的两性小分子,SDS 、Triton X-100(p92)①脂筏:以甘油磷脂为主体的生物膜上,胆固醇、鞘磷脂等形成相对有序的脂相,如同漂浮在脂双层上的“脂筏”一样,载着执行某些生物学功能的各种膜蛋白。
脂筏最初可能在内质网或高尔基体上形成,最终转移到细胞膜上。
有些脂筏与膜下细胞支架蛋白交联。
推测一个直径100nm的脂筏可载有600个蛋白分子。
②膜脂分类:④内在膜蛋白的结合方式1、内在膜蛋白的跨膜结构域与脂双层分子的疏水核心的相互作用;(最主要的)2、内在膜蛋白的跨膜结构域两端携带正电荷的氨基酸或者负电荷的氨基酸,正电荷则与极性头(带负电荷)形成离子键,带负电荷则通过Ca2+、Mg2+等阳离子与极性头(带负电荷)相互作用;3、通过细胞质基质一侧的半胱氨酸残基上共价结合的脂肪酸分子插入到膜双层之间。
P91(α螺旋外侧非极性链,内测极性链,形成跨膜通道(这里的内侧和外侧值得是α螺旋自身,并非伸向细胞质和细胞外液的部分);孔蛋白还具有β折叠片→生化内容)。

脂筏的结构功能及与某些疾病的研究进展自从Nicolson在1972年提出膜流动镶嵌结构模型以来,人们对生物膜的研究不断深入,在1988年Simons提出了‘脂筏’的概念,Brown等[1]在1992年通过实验提出了脂筏的假说,直到1997年的Simons [2]等才正式用脂筏这个名称来描述这种富含胆固醇和鞘脂的动态微区。
至今有许多关于膜结构的报道,发现膜中有富含胆固醇及鞘脂的微区(microdomain),这个区曾有过许多名称:不溶于去垢剂的糖脂富含区(detergent-insoluble glycolipid-rich domain,DIFs);低浓度Triton不溶复合物(low-density triton-insolublecomplex,LDTI);富含糖脂的膜(glycolipid-enriched membrane,GEMs).2001年在西班牙召开的欧洲研究讨论会(European Research Conference,EURESCO)上对“微区、脂筏(lipid raft)及小窝(caveolae)”作了讨论,Meer于次年综述了此专题。
1.脂筏脂筏是脂质双层内含有特殊脂质的蛋白质的微区,具有低流动性,呈现有序液相,富含胆固醇和鞘磷脂[4],形成相对稳定的具有一定功能的筹结构漂浮于二维流动的细胞质膜中,被人们形象地称之为“功能筏”。
具有参与胞吞胞饮、信号转导、运输胆固醇等重要功能,随着人们在分子水平上认识的不断深化,脂筏结构和功能的物理、化学基础研究方面也取得了初步的进展。
在脂双层的不同区域有不同的脂筏,而且这些脂筏是运动的。
脂筏可能有 3 类:小窝、富含糖鞘脂膜区、富含多磷酸肌醇膜区。
不同的脂筏有其各自特异的蛋白质,所含脂质也不完全相同,并且有不同的功能。
脂筏的主要成分是鞘磷脂、神经节苷脂及胆固醇。
由于鞘磷脂含有长链饱和脂肪酸,与胆固醇相互作用成一种有序脂质相,其相变温度(Tm)较高,使脂膜的流动性降低而稳定性增加。

高中生物第3章细胞的基本结构知识点总结归纳单选题1、下列有关细胞的组成、结构和功能的叙述,错误的是()A.活细胞内水和无机盐的主要存在形式分别是自由水和离子B.植物细胞的最外层结构具有将细胞与外界环境分隔开的功能C.若某真核细胞内不能合成蛋白质,则有可能是细胞核出现了问题D.吞噬细胞内溶酶体较多,有利于消化其吞噬的细菌或病毒答案:B分析:1 .植物细胞壁的主要成分是纤维素和果胶,对植物细胞具有支持和保护的作用;2 .核仁与某种RNA的合成和核糖体形成有关。
3 .溶酶体内含有水解酶,能够消化其吞噬的细菌或病毒,处理衰老、受损的细胞器。
4 .细胞内的水包括自由水和结合水。
A、活细胞内的水主要以自由水的形式存在,无机盐主要以离子的形式存在,A正确;B、植物细胞的最外层结构是细胞壁,而细胞壁是全透性的,不具有将细胞与外界环境分隔开的功能,B错误;C、蛋白质的合成场所是核糖体,而真核细胞核糖体的形成离不开细胞核(核仁),C正确;D、吞噬细胞能吞噬侵入机体的细菌和病毒,而溶酶体能将侵入细胞的细菌或病毒消化分解,D正确。
故选B。
2、如图是细胞局部结构示意图,下列叙述正确的是A.①是糖脂,具有保护和润滑的作用B.②具有运输、信息传递、免疫和构成膜基本支架的作用C.③可以是细胞器膜、核膜的主要成分D.④由纤维素组成,能保持细胞内部结构有序性答案:CA、图示为细胞膜的流动镶嵌模型,①是糖蛋白,具有识别和信息交流的作用,A错误;B、②是膜蛋白,具有运输、信息传递、免疫等作用,磷脂双分子层是构成膜的基本支架,B错误;C、③是磷脂分子,是细胞器膜、核膜等各种生物膜的主要成分,C正确;D、④由蛋白质组成,能保持细胞内部结构有序性,D错误。
故选C。
小提示:生物膜的基本骨架是磷脂双分子层,蛋白质分子以不同程度地镶嵌在磷脂双分子层中,生物膜的结构特点是具有一定的流动性,功能特点是具有选择透过性;生物膜功能的复杂程度取决于膜蛋白的种类及数量,尤其需要注意常考的四种“膜蛋白”:(1)信号分子(如激素、淋巴因子、神经递质)的受体膜蛋白——糖蛋白。
1、脂筏模型lipid rafts model:该模型认为在甘油磷脂维生物膜的主体上,胆固醇、鞘磷脂等富集区域形成相对有序的脂相,如同漂浮在脂双层上的“脂筏”一样载着某些特定生物学功能的各种膜蛋白。
P55 在生物膜上胆固醇富集而形成有序脂相,如同脂筏一样载着各种蛋白.脂筏是质膜上富含胆固醇和鞘磷脂的微结构域。
大小约70nm左右,是一种动态结构,位于质膜的外小页。
2、蛋白protein,:蛋白能调节细胞周期和避免细胞癌变发生。
p533、界限Hayflick limitation Hayflick:细胞停止分裂是由细胞自身因素决定的,与环境条件无关,正常细胞具有有限分裂次数,而癌细胞能够在体外无限增殖。
P356细胞,至少是培养的二倍体细胞,不是不死的,而是有一定的寿命;它们的增殖能力不是无限的,而是有一定的界限,这就是 Hayflick 界线。
4、细胞系cell line:原代培养的细胞一般传至10代左右就不易传下去,细胞生长出现停滞,大部分细胞衰老死亡,但有极少数细胞可能渡过“危机” 而传下去。
这些存活的细胞一般又可顺利地传 40-50 代次,并且仍保持原来染色体的二倍数量及接触抑制的行为。
P435、核定位信号Nuclear localization signal (NLS):亲核蛋白一般都含有特殊的氨基酸序列,这些内含的特殊短肽保证了整个蛋白质能够通过核孔复合体被转运到细胞核内,这段具有“定向” “定位”作用的序列被命名为核定位信号。
P2326、细胞程序性死亡programmed cell death (PCD):无论是单细胞生物还是多细胞生物,细胞死亡往往受细胞内由遗传机制决定的“死亡程序”控制,要求特定基因表达,是“主动”而非“被动”的过程。
P3417、生物膜biomembrane:真核生物内部存在由膜围绕构建的各种细胞器。
细胞内的膜系统和细胞质膜统称为生物膜。
P548、第二信使Second messenger;:是指在胞内产生的非蛋白类小分子,通过其浓度变化(增加或减少)应答胞外信号与细胞表面受体的结合,调节细胞内酶和非酶蛋白的活性,从而在细胞信号转到途径中行使携带和放大信号的功能。
脂筏的结构与功能3陈 岚 许彩民33 袁建刚 潘华珍(中国医学科学院基础医学研究所中国协和医科大学基础医学院医学分子生物学国家重点实验室,北京100005)摘要 脂筏是膜脂双层内含有特殊脂质及蛋白质的微区.小窝是脂筏的一种类型,由胆固醇、鞘脂及蛋白质组成,以小窝蛋白为标记蛋白.脂筏的组分和结构特点有利于蛋白质之间相互作用和构象转化,可以参与信号转导和细胞蛋白质运转.一些感染性疾病、心血管疾病、肿瘤、肌营养不良症及朊病毒病等可能与脂筏功能紊乱有着密切的关系.关键词 脂筏,小窝,小窝蛋白,信号转导,糖肌醇磷脂2锚固蛋白学科分类号 Q51 自从1972年Nicolson提出膜流动镶嵌结构模型以来,至今有许多关于膜结构的报道,发现膜中有富含胆固醇及鞘脂的微区(microdomain),这个区曾有过许多名称:不溶于去垢剂的糖脂富含区(detergent2ins oluble glycolipid2rich domain,DIFs);富含糖脂的膜(glycolipid2enriched membrane,GEMs);低浓度T riton不溶复合物(low2density triton2ins oluble complex,LDTI).2001年在西班牙召开了欧洲研究讨论会(European Research C onference,EURESCO),会上对“微区、脂筏(lipid raft)及小窝(caveolae)”作了专题的讨论,Meer于次年综述了此专题[1].下面简要介绍小窝及脂筏的结构与功能.1 小窝与小窝蛋白(caveolin)1953年Palade用非离子去污剂提取内皮细胞及平滑肌细胞膜时,发现在低温下用非离子去污剂提取不溶的部分,经浮力密度分离,仍可再分成两部分,较沉的是骨架蛋白,上浮的是膜的另一些组分,从形态观察似在质膜上的囊泡,他设想这种囊泡可游走穿梭在细胞之间,所以起名为质膜囊泡(plasmalemmal vesicles).1955年Y amado报道了与Palade相似的结果,将这种囊泡称小窝(caveolae),是拉丁字小洞(cave)的意思.Y amado所称的小窝即是现在所说的“脂筏”之一,而且是其中最主要的一种.小窝由胆固醇、鞘脂(糖化神经鞘脂、神经鞘磷脂)及蛋白质组成.大约有50~100nm,形态有多种多样,根据其功能不同有瓶型、囊泡型及管型,多数是瓶型.在细胞表面有开放型,如形成胞吐囊泡;也有封闭型,如形成胞吞囊泡.从冷冻刻蚀图观察,可见小窝表面有曲纹覆盖.经分析,曲纹主要由小窝蛋白结合胆固醇而成.小窝的组装分两个步骤:首先在高尔基体由鞘脂及胆固醇形成去污剂不溶的脂质核,内质网合成的糖肌醇磷脂2锚固蛋白(glycosylphosphatidylinosital2anchored protein,GPI2 anchored protein)和小窝蛋白嵌入脂核体,形成初步的组装体,然后运往细胞表面.在此过程中小窝蛋白及胆固醇起关键作用,如果敲除小窝蛋白的基因,即不能形成小窝.胆固醇是在内质网合成,如合成受抑制,也不能形成小窝,胆固醇与小窝蛋白的比例大约在415∶1.所以小窝蛋白可作为小窝的标记蛋白[2]. 小窝蛋白大约至少有4种:小窝蛋白1α、1β、2、3.在膜内,小窝蛋白21易形成同型二聚体;在脂肪细胞中,小窝蛋白21、22共表达,形成异二聚体;小窝蛋白23主要分布在肌肉,其他细胞较少.小窝蛋白的结构中都含有3个半胱氨酸(位于134,144,157)并在此部位进行酰化.小窝蛋白是一种绞架蛋白(scaffolding protein),绞架区基本已清楚,它可直接与胆固醇及鞘脂结合,又可与信息分子(Src2kinase,H2ras,eNOS,G2protein等)结合[3].小窝能募集多种蛋白质可能与此结构有关.2 脂 筏脂筏是指膜脂双层内含有特殊脂质及蛋白质的 3国家自然科学基金重点资助项目(30130070). 33通讯联系人. Tel:010*********,E2mail:caiminxu@ 收稿日期:2002209203,接受日期:2002211201微区,微区内陷可形成囊泡,近年发现脂筏不仅存于质膜,亦可在高尔基体膜上.脂筏的概念早在发现小窝时即有了,经长期的争论,直至1988年Simon 才正式提出“脂筏”之称.脂质的双层有不同的脂筏:外层的微区主要含有鞘脂、胆固醇及GPI 2锚固蛋白,因为鞘脂含有长链饱和脂肪酸,T m 温度较高,流动性差,而且粘稠,邻近的磷脂区其脂肪酸多不饱和,T m 温度较低,所以出现分相;膜内侧也有相似的微区,与外侧的脂质不完全相同,主要是在此区有许多酰化的蛋白质,特别是信号转导蛋白(图1).虽然两层分别有脂筏,但它们是偶联的,因为用非离子去污剂提取时,不仅有外层的GPI 2锚固蛋白,还有许多信息分子共同被提出.用一种GPI 2锚固蛋白的抗体介导锚固蛋白聚集,与此同时Src 家族的酪氨酸激酶也被激活.如换成糖鞘脂的抗体,也有同样的现象.GPI 蛋白及糖鞘脂都存于膜外侧,Src 酪氨酸激酶在膜内侧,这表明脂筏内外层之间是有联系的[4].Fig 11 Model of lipid rafts图1 脂筏模式图 用不同的去污剂或改变溶解时的温度,所得到的脂质及蛋白质都有差异,说明细胞膜上的脂筏并不都等同.脂筏可能有三类:小窝、富含糖鞘脂膜区(glycosphingolipid enrich membrane )、富含多磷酸肌醇(PIP2)膜区.不同的脂筏有其各自的特异蛋白,并有不同的功能.脂筏的部分脂质及蛋白质的组分见表1.T able 1 partial lipid and protein components of lipid raft [5]表1 脂筏的部分脂质及蛋白质的组成[5]脂质蛋白质受体蛋白胆固醇、鞘脂、糖鞘脂(如GM1)、PIP2、花生四烯酸小窝蛋白、flotillins 、stomatins 、MAL/BENE 、LAT/PAG 、VIP36、GPI 2锚固蛋白类(如叶酸受体,疯牛病蛋白,碱性磷酸酶等)、非受体酪氨酸激酶(Src 2家族)、G蛋白、一氧化氮合酶(eNOS )、Ras 、Fyn 、PKCα受体酪氨酸激酶(RTKs ,如EGF 2R 、PD GF 2RInsulin 2R 等) 脂筏内的蛋白质,有的是经跨膜直接插入膜,但更多的蛋白质需酰化,由酰化后的脂肪酸插入膜.Zacharias [6]的实验表明,用水母蛋白的两个变异体,分别融合蓝荧光蛋白(CFP )及绿荧光(YEP )蛋白,然后在多肽链上分别联接脂化(棕榈酸、豆蔻酸、异戊二烯)的特异共同序列,用荧光能量共振转移(fluorescence resonance energy transfer )测定邻近效应,发现棕榈酰化及豆蔻酰化的蛋白质在脂筏的小窝区,而异戊二烯化的蛋白质形成二聚体,不在小窝区,对胆固醇不敏感,可能是另一类的脂筏.说明不同的酰化蛋白插入不同的脂筏. Stuermer 在神经元细胞膜内,发现两个新蛋白质———flotillin 21及flotillin 22,其不溶于去污剂,与脂质形成脂筏.用其抗体进行免疫组化染色,在光镜及电镜下可见,flotillin 21,2排列在细胞膜的胞浆面,在延伸的轴突及丝足上都存在.它不仅存于神经细胞,而且在人Jurkat T 2淋巴细胞株的细胞上也可被发现,并且与Thy21,Fyn蛋白交联,参与淋巴细胞活化,如加入Thy21抗体,可见与flotillin21,2共聚,但在这个脂筏内没有小窝蛋白,这说明,含有flotillin的结构可组成另一类的脂筏.Salzer用钙诱导红细胞时,从形态及生化的分析发现有两种囊泡:一种称微囊泡(microvesicle),直径约150nm;另一种称纳诺囊泡(nanovesicle),直径约60nm.两种囊泡除都共有GPI2锚固蛋白外,其他的组分不完全相同.微囊泡含有两种蛋白质:stomatin及flotillin21,2.Stomatin是棕榈酰化蛋白质,以棕榈酸插入脂筏区,作用很像小窝蛋白.Flotillin21,2也在脂筏区,但它没有被酰化,在囊泡及质膜都存在.纳诺囊泡还有另两种蛋白质———会联蛋白(synexin)及抗药蛋白(s orcin).会联蛋白是膜联蛋白(annexin)家族的成员annexinⅦ(可介导分泌小泡的聚集),负责细胞内运转及膜的组装.它与钙结合后,转到膜上形成膜融合蛋白.抗药蛋白也可与钙结合,结合后,暴露出疏水基团,直接插入膜,有观点认为抗药蛋白与会联蛋白都在脂筏上,但分别在不同的区域或两者结合后进入脂筏.这说明在红细胞的囊泡中也有脂筏[7].3 脂筏的功能从结构及组分分析,脂筏有两个特点:a1许多蛋白质聚集在脂筏内,便于相互作用.b1脂筏的环境有利于蛋白质的变构,形成有效的构象.所以它具有许多功能.311 参与信号转导由于脂筏内有多种信号分子,它可以参与许多的信号转导通路.以T细胞为例:T细胞膜上有T 细胞抗原抗体(TCR),当它活化后可引起许多信息分子聚集形成复合体.最近Zhang[8]报道这种复合体形成的关键是脂筏提供了平台.在静止的T 细胞脂筏内富含Src家族激酶Lck、Fyn及跨膜的衔接子(linker activated T cell,LA T),还有少量的CD4和CD3,其中最重要的是LA T.它是一个跨膜蛋白,是伸向胞浆的一段肽链,富含酪氨酸,另外LA T的半胱氨酸还可酰化棕榈酸,以此插入脂筏.当TCR被激活,Src酪氨酸激酶活化引起LA T的多个酪氨酸被磷酸化,然后,LA T募集多种底物进行磷酸化,包括ZAP70、PLCγ1、PI3K、Vav,形成活化的大复合体.这些反应都必须在脂筏内完成,如果LA T不能进入脂筏,T2细胞便不能执行其重要的增殖及分化功能.其他免疫识别受体,如B细胞抗原受体(BCR)、肥大细胞高亲和IgE受体(FcζRI)等也都以脂筏作为传递信息的场所.Roy[9]报道调节小窝蛋白的表达,可直接影响信息蛋白分子(包括癌基因).他发现小窝蛋白21的启动子有胆固醇的调节位点,小窝蛋白在小窝内有运转及调节胆固醇水平的作用,所以调节小窝蛋白的表达,可以间接影响信息的传递.Parton制备了一个小窝蛋白的变异体(gavD GV),可抑制H2Ras介导Raf的活化,如加入胆固醇可改善抑制作用.他设想H2Ras是Ras的异构体,有棕榈酸化,需要富有胆固醇的脂筏才能活化Raf,这个筏区被小窝蛋白变异体破坏.HIa[10]发现在细胞质膜有溶血卵磷脂类(L Ps)的受体,这种受体是一个大家族,大约有12个左右,它与G蛋白偶联,称G蛋白偶联受体(G protein2coupled receptors,GPCRs).L Ps受体位于膜富含鞘磷脂的小窝区.L Ps包括溶脂酸(L PA)、溶血卵磷脂(L PC)、鞘磷酸胆碱(SPC)及鞘磷脂212磷酸(S2I2P).L Ps受体的作用包括:参与调节细胞迁移、增殖及存活;调节血管系统的成熟;激活eNOS及活化磷脂酶C(P KC)等.Nanjundan[11]从鼠肺细胞内提取出一种脂质磷酸水解酶(LL P),活化的LL P存于小窝内,它可水解L PA、PA和鞘氨醇212磷酸,水解后的产物皆为第二信使.312 参与细胞蛋白运转31211 参与跨细胞运转:文献[2]报道,分布在内皮细胞的小窝囊泡,开口于细胞的表面,细胞与细胞之间的囊泡可形成一个通道,小分子物质可相互交换,小窝成为一些小分子化合物运转的通路. 31212 参与细胞胞饮作用:小窝内包含许多受体蛋白,小窝将它们运送到细胞内可利用不同的途径:a1受体与配基结合后,被小窝囊泡运送到胞浆,配基与受体分离,受体随囊泡回到质膜;b1小窝囊泡载着受体与配基,通过胞浆,在细胞的反方向开口将配基释放到细胞外基质.有些毒素(如霍乱毒素)与细胞上的受体结合,也以这种方式进入宿主细胞内[2].31213 参与细胞分选:Martin[12]介绍了一个膜内在蛋白MAL.MAL属酰化蛋白质家族,存于Madin2 Darby犬肾细胞(Madin2Darby canine kidney, MDCK),有多个疏水基团插入极化的上皮细胞(polarized epithilia)的脂筏.MAL直接参与极化分选(apical s orting).在MDCK细胞内如加入MAL反义寡核苷酸,就无分选功能.MAL即像一个货车,可在质膜、内质网、高尔基体之间循环,运送蛋白.另外BENE蛋白,和MAL一样,也属酰化蛋白质家族.它与小窝蛋白21结合,位于小窝内,可能这两个蛋白质是脂筏的新成员.Millan[13]报道当T2淋巴细胞执行其功能时,发生极化形态.细胞的一端延伸,形成特殊的突起———“尾足”(uropod),随后选择性地将一些粘附分子(ICAM21,ICAM22,ICAM23,CD43,CD44)浓缩在尾足的脂筏区.血粘素(hemaglutinine,HA)在T淋巴细胞的高尔基体合成后,进入脂筏囊泡,随囊泡也被送到尾足的表面,说明尾足富含脂筏.细胞的另一端称前缘,对抗原及细胞因子很敏感,其复杂的免疫反应都在前缘脂筏区进行.4 脂筏与疾病脂筏的生理功能具有多向性,如果它的正常生理过程发生改变,则会引发多种疾病.近来由于一些小窝蛋白基因缺陷的动物模型的建立,为人们理解和发现与脂筏相关的疾病创造了条件.411 脂筏与感染性疾病细胞内吞作用可以发生在小窝结构上.许多病原体如病毒、寄生虫、细菌及其毒素都能经由小窝进入宿主细胞.这些病原体或毒素通过小窝的内化作用进入宿主细胞后,并不与溶酶体融合(与溶酶体融合导致内吞物的降解是经典细胞内吞作用的途径),这样,外源物质就可以避开机体的免疫反应和一系列的清除行为,得以在体内生存并发挥其致病作用.Abraham等[14]向人们展示了细菌在小鼠骨髓源性肥大细胞(mouse bone marrow2derived mast cells,BMMCs)上经由小窝包裹形成囊后进入噬菌细胞的机制.Oppenheim等的实验说明了SV40经由小窝内吞后,未与溶酶体结合,而转运至内质网的特殊感染途径.用药物阻断这一通路中某些蛋白质的组装,就可以抑制感染的发生[15,16].目前还有待在体内实验和分子水平上阐述小窝在感染性疾病中发挥的作用,借此,人们将利用更为合理的传送途径来设计药物以达到更为有效的阻断病原体感染的治疗目标.412 脂筏与心血管疾病内皮来源的一氧化氮生物利用率低下,在动脉粥样硬化症发生过程中起着关键作用.内皮型一氧化氮合成酶(eNOS)直接受到小窝蛋白的调节,与小窝关系密切.实验表明,利用氧化型的低密度脂蛋白作为胆固醇的接收体,可以导致小窝的耗竭,也可以抑制由乙酰胆碱诱导的eNOS的活性,但不影响eNOS的数量和环前列腺素的产量,也不影响eNOS的肉豆蔻酰化、棕榈酰化和磷酸化.去除氧化型低密度脂蛋白,小窝的结构可以得到恢复,并且eNOS亦可再与小窝结合[17].Bucci等在体内实验中证实了小窝蛋白21对eNOS信号通路起着首要的作用,一旦eNOS通路出现抑制,则会消除由NO介导的血管渗透和收缩反应.随后, Razani[18]和Drab两个研究小组利用敲除小窝蛋白2 1基因的小鼠,在离体大动脉血管收缩紧张度测量的实验中发现,小窝蛋白21的缺乏导致eNOS活性水平低并导致血管弹性降低,在动脉血管中也发现了这一现象.另外,糖化终末产物(advanced glycation end products,A GEs)通过其受体介导,在糖尿病并发症中发挥着重要的作用.Stitt等[19]发现,在视网膜微循环内皮细胞中,A GEs与其受体的相互作用是在小窝中进行的.由此表明,小窝的内化及信号转导功能或许可以调控A GE受体介导的血管功能障碍.413 脂筏与肿瘤许多原癌基因和抑癌基因是信号转导通路中处于不同阶段的蛋白质,当它们的生理功能发生改变或缺失时就会促进肿瘤形成.脂筏可以参与信号转导.由此自然让人联想到,脂筏结构和功能的变化与肿瘤发生有密切的关系.小窝蛋白21基因被作为一种候选的抑癌基因.小窝蛋白21的mRNA和蛋白质表达水平在肿瘤转化的N IH23T3培养细胞中出现下调,在乳腺癌转基因小鼠中以及其他起源于人肿瘤的细胞系中这种基因的表达也出现下调[20].一般来说,确定小鼠的一种原癌基因或抑癌基因需要分析原始鼠胚胎纤维原细胞的生长特性,因此,在对小窝蛋白21进行了细胞增殖和细胞周期评估后,发现小窝蛋白21敲除的小鼠胚胎纤维原细胞出现细胞周期整体活性增加,主要出现合成期的增加[18].Pramudji等[21]利用腺病毒载体介导小窝蛋白21启动子对原位前列腺癌动物模型进行基因治疗,结果出现新生物的坏死.肿瘤抗药性是导致对癌症患者化疗失败的原因.研究发现多药耐药(multidrug resistant, MDR)的癌细胞可表达高水平和高密度的小窝[22].这给人们了解多药效耐药细胞的产生机制和设计有针对性的药物起到了指导作用.414 脂筏与肌营养不良症小窝蛋白21和小窝蛋白22广泛分布在组织细胞上,而小窝蛋白23仅仅局限地分布于肌肉组织,这一点显现出小窝蛋白23可能在肌肉组织功能障碍中起到某些作用.杜兴肌营养不良(Duchenne muscular dystrophy, DMD)是一种严重的肌肉功能紊乱疾病,其特征是缺乏了肌营养不良蛋白.肌营养不良蛋白是一种细胞内的蛋白质,是DMD基因的表达产物,可以联系细胞内的细胞骨架和细胞外基质,这一作用对于保持正常肌肉功能和肌肉纤维的完整性是极为必要的.免疫电镜观察到肌营养不良蛋白局限在肌肉细胞的小窝上[23].还有一些形态学和生物化学的观察发现,在DMD病理过程中有小窝和小窝蛋白23的参与.很早的时候,人们利用电镜和冰冻蚀刻技术在DMD患者的骨骼肌中发现小窝数目的增多.与此结果相一致的是,最近,在mdx小鼠(一种肌营养不良蛋白缺陷的DMD动物模型)和患者的骨骼肌中发现,小窝蛋白23的表达增高了2~3倍.在超表达小窝蛋白23的转基因小鼠中,表现出杜氏肌营养不良样症状[24].这种转基因模型的建立,有助于人们更好地研究和理解人类DMD的病理发生机制.肌带型肌营养不良(limb2girdle muscular dystrophy,L GMD21C)是另一种肌营养不良症,是由于小窝蛋白23基因突变造成的,这种突变基因编码的蛋白质很不稳定,易被降解,所以存在甚少.G albiati等[25]模拟人的基因突变点,构建缺失小窝蛋白23基因的小鼠,证实小窝蛋白23是形成肌细胞内小窝的主体.正常动物肌细胞内小窝蛋白23与肌营养不良蛋白及糖蛋白形成复合体,存于脂筏.基因突变的小鼠,无脂筏,所以不能形成复合体.说明小窝蛋白23的突变是产生L GMD21C的分子基础.由此可见,正常生理条件下,小窝蛋白的表达是一个动态平衡的过程.415 脂筏与朊病毒病朊病毒病包括人克雅氏病(Creutzfeldt2Jakob Disease,C JD)、羊瘙痒症和牛海绵样脑病等,其发病原因是细胞朊蛋白(prion protein,PrP C)的构象转变为异常形式(PrP SC).PrP C是结合在细胞膜外表面,带有GPI锚结构的糖蛋白.研究表明,小鼠的神经瘤母细胞(N2a)的PrP C聚集在小窝样结构域中,经scrapie感染后N2a(ScN2a)的PrP SC也聚集在相应的结构中.细胞胆固醇水平的降低可以抑制PrP C向PrP Sc的转化.这就提示人们,富含胆固醇的小窝样结构域很可能就是朊病毒传播增殖的场所.Kaneko等[26]证实PrP Sc是由GPI 锚固形式的PrP C转变而来的,并不是来自跨膜的PrP C.由于GPI是小窝结构的标志性蛋白质之一,这也证实了上述实验的结论.416 脂筏与肺部疾病肺泡Ⅰ型上皮细胞富含小窝蛋白21.在敲除小窝蛋白21基因的动物中,肺泡空间明显变得狭小,肺泡间隔由于不可控制的上皮细胞增殖和纤维化而变厚,导致严重的肺功能紊乱,这种小窝蛋白21基因敲除的小鼠在游泳实验中很容易疲劳[18].5 结束语人们对脂筏结构与功能的研究由来已久,特别是近几年虽然有许多问题已较清楚,但仍存在不少有待深入研究的问题:a1膜内外层的脂筏组分有何异同,特别是内膜很难提取,至今对其组分还不清楚;b1蛋白质如何识别不同的脂筏,进入一个有效区;c1脂筏内跨膜蛋白与脂筏内的酰化蛋白之间如何相互作用;d1脂筏在膜内如何游走,虽然对脂筏与细胞骨架关系的了解有了良好的开端,但是尚缺乏更深入的研究.期望应用先进的技术,解决在无试剂干扰的条件下,观察天然单细胞膜脂与膜蛋白的活动.从对小窝蛋白的研究,可看到它与癌症、动脉粥样硬化、糖尿病并发症、早老性痴呆、疯牛病及肌营养不良等疾病的关系.同时,对脂筏结构和功能的研究将会给生物学及医学带来更多的信息与启示.参 考 文 献1 Meer G V.The different hues of lipid rafts.Science,2002,296 (5569):855~8572 Anderson R G W.The caveolae membrane system.Ann Rev Biochen,1998,67:199~2253 Massimino M L,Griffoni C,Spisni E,et al.Involvement of caveolae and caveolae2like domain in signaling cell survival and angiogenesis.Cell Signal,2002,14(2):93~984 Brown D A,London E.Structure and function of sphingolipid and cholesterol2rich membrane rafts.J Biol Chem,2000,275(23): 17221~172245 G albiati F,Razani B,Lisanti M P.Emerging themes in lipid rafts and caveolae.Cell,2001,106(4):403~4116 Zacharias D A,Violin J D,Newton A C,et al.Partitioning of lipid2modified monomeric GFPs into membrane microdomains of live cells.Science,2002,296(5569):913~9167 Salzer U,Hinterdorfer P,Hunger U,et al.Ca2+dependent vesicle release from erythrocytes involves stomatin2specific lipid rafts,synexin(annexinⅦ),and sorcin.Blood,2002,97(7): 2569~25778 Zhang W,Sloan2Lancaster J,K itchen J,et T:the ZAP270 tyrosine kinase substrate that links T cell receptor to cellular activation.Cell,1998,92(1):83~929 Roy S,Luetterforst R,Harding A,et al.Dominant2negative caveolin inhibits H2Ras function by disrupting cholesterol2rich plasma membrane domains.Nat Cell Biol,1999,1(2):98~10510Hla T,Lee M J,Ancellin N,et al.Lysophospholipids2receptor revelations.Science,2001,294(5548):1875~187811Nanjundan M,Possmayer F.Pulmonary lipid phosphate phosphohydrolase in plasma membrane signaling platforms.Biochem J,2001,358(Pt3):637~64612Martin2Belmonte F,Puertollano R,Millan J,et al.The MAL proteolipid is necessary for the overall apical delivery of membrane proteins in the polarized epithelial Madin2Darby canine kidney and fischer rat thyroid cell lines.Mol Biol Cell,2000,11(6):2033~204513Millan J,Montoya M C,Sancho D,et al.Lipid rafts mediate biosynthetic transport to the T lymphocyte uropod subdomain and are necessary for uropod integrity and function.Blood,2002,99(3):978~98414Shin J S,Abraham S N.Caveolae2not just craters in the cellular landscape.Science,2001,293(5534):1447~144815Norkin L C,Anderson H A,Wolfrom S A,et al.Caveolar endocytosis of simian virus40is followed by brefeldin A2sensitive transport to the endoplasmic reticulum,where the virus disassembles.J Virol,2002,76(10):5156~516616Richards A A,Stang E,Pepperkok R,et al.Inhibitors of COP2 mediated transport and cholera toxin action inhibit simian virus40 infection.Mol Biol Cell,2002,13(5):1750~176417Alison B,Philip W S,Ivan Y,et al.Smart oxidized low densitylipoprotein displaces endothelial nitric2oxide synthase(eNOS)from plasmalemmal caveolae and impairs eNOS activation.J Biol Chem, 1999,274(45):32512~3251918Razani B,Engelman J A,Wang X B,et al.Caveolin21null mice are viable,but show evidence of hyper2proliferative and vascular abnormalities.J Biol Chem,2001,276(41):38121~38138 19Stitt A W,Burke G A,Chen F,et al.Advanced glycation end2 product receptor interactions on microvascular cells occur within caveolin2rich membrane domains.FASEB J,2000,14(15): 2390~239220Razani B,Schlegel A,Lisanti M P.Caveolin proteins in signaling, oncogenic transformation and muscular dystrophy.J Cell Sci, 2000,113(Pt12):2103~210921Pramudji C,Shimura S,Ebara S,et al.Timme in situ prostate cancer gene therapy using a novel adenoviral vector regulated by the caveolin21promoter.Clin Cancer Res,2001,7(12):4272~427922Lavie Y,Fiucci G,Liscovitch M.Up regulation of caveolae and caveolar constituents in multidrug resistant cancer cells.J Biol Chem,1998,273(49):32380~3238323G albiati F,Razani B,Lisanti M P.Caveolae and caveolin23in muscular dystrophy.Trends Mol Med,2001,7(10):435~44124G albiati F,Volont D,Chu J B,et al.Transgenic overexpression of caveolin23in skeletal muscle fibers induces a Duchenne2like muscular dystrophy phenotype.Proc Natl Acad Sci USA,2000, 97(17):9689~969425G albiati F,Engelman J A,Volonte D,et al.Cavelin23null mice show a loss of caveolar,changes in the microdomain distribution of the dystrophin glycoprotein complex,and T2tubule abnormalities.J Biol Chem,2001,276(24):21425~2143326Kaneko K,Vey M,Scott M,et al.COOH2terminal sequence of the cellular prion protein directs subcellular trafficking and controls conversion into the scrapie isoform.Proc Natl Acad Sci USA, 1997,94(6):2333~2338Structure and Function of Lipid R afts3CHEN Lan,XU Cai2Min33,YUAN Jian2G ang,PAN Hua2Zhen(N ational L aboratory of Medical Molecular Biology,Basic Instit ute of Medical Sciences,The Chi nese Academy of Medical Sciences,Peki ng U nion Medical College,Beiji ng100005,China)Abstract Lipid rafts are liquid2ordered membrane microdomains with a unique protein and lipid composition found on the plasma membrane.Caveolae,a type of lipid rafts,is characterized by high levels of cholesterol, sphingolipids and proteins,and is identified by the presence of the protein caveolin.The structure and component of lipid rafts is used of reciprocity and comformational change between proteins.Lipid rafts are associated with signal transduction and cellar proteins movement.The disfunction of lipid rafts is related to some diseases such as infection,heart disease,cancer,muscular dystrophy and prion protein diseases.K ey w ords lipid raft,caveolae,caveolin,signal transduction,GPI2anchored protein 3This work was supported by a grant from The National Natural Sciences Foundation of China(30130070). 33Corresponding author.Tel:86210265296445,E2mail:caiminxu@ Received:September3,2002 Accepted:November1,2002。
一、名词解释1、O-连接糖链:糖链的N-乙酰半乳糖胺与多肽链的丝氨酸或苏氨酸的羟基连接,形成O-糖苷键,糖链为O-连接糖链,也称O-连接聚糖。
2、MAPK:有丝分裂原激活的蛋白激酶,是一类丝氨酸/苏氨酸蛋白激酶,该激酶家族所有成员的一个共同结构特征是一级结构中第185和187个氨基酸残基分别是可磷酸化的苏氨酸和酪氨酸。
两个氨基茇酸残基必需同时磷酸化才能被激活。
3、双功能蛋白激酶4、糖型5、旁分泌与自分泌:一些胞外化学信号分子如生长因子,细胞因子被分泌后很快被降解,只能对邻近的细胞或自身细胞起作用,这种细胞间通讯方式称旁分泌或自分泌。
6、脂交叉双层7、Grb2:生长因子受体结合蛋-28、Sos:GTP交换因子9、糖结合位点10、GPCR:G蛋白偶联受体,G蛋白偶联受体主要参与各种激素,化学趋化因子的信息跨膜转换过程。
目前已发现有数百种化学信号分子由G蛋白偶联受体来传递信号。
各种胞外信号分子作用于受体后,受体的化学信号由G蛋白进一步向胞内传递。
G蛋白通过激活或抑制质膜内侧的能产生第二信使的酶或离子通道产生第二信使,将胞外信号转变为胞内信号。
11、受体型酪氨酸蛋白激酶:是具有酶活性的催化性受体。
为跨膜蛋白,由胞外区,跨膜区和胞内区三部分组成。
其配体是各种生长因子。
除胰岛素受体外均为单跨膜蛋白。
其胞外部分是与生长因子结合部位,胞内部分具有酪氨酸蛋白激酶活性。
根据结构和配体不同分为十几个亚家族。
12、SH2 结构:SH2 结构专门识别蛋白肽链磷酸化的酪氨酸残基。
SH2 结构存在于多种信息分子中,包括接头蛋白,酪氨酸蛋白激酶,蛋白磷酸酶,小分子G蛋白,GTP交换因子,转录因子等。
13、GPI:糖磷脂酰肌醇(GPI)与多肽链连接,此类蛋白质称为GPI-连接糖蛋白或GPI-锚定糖蛋白。
14、PH结构:血小板-白细胞激酶同源结构,专门识别多磷酸肌醇磷脂,尤其是3位磷酸化的肌醇磷脂如PI-3-P, PI-3,4-P2, PI-3,5-P2和PI-3,4,5-P3等。
某理工大学生物工程学院《细胞生物学》课程试卷(含答案)__________学年第___学期考试类型:(闭卷)考试考试时间:90 分钟年级专业_____________学号_____________ 姓名_____________1、判断题(20分,每题5分)1. 虽然龟的最高寿命是175岁,而小鼠的寿命只有几年,但它们的细胞在体外培养时分裂的极限基本相同。
()答案:错误解析:龟细胞体外培养可分裂100次以上,而小鼠细胞在体外分裂的次数一般不超过30次。
2. 所有组成核小体的组蛋白,在进化上都是高度保守的。
()答案:错误解析:H1有一定的种属和非政府玉蕊科特异性,在演化过程上不如上才其他组蛋白那么保守。
3. 原核细胞中的核糖体都是70S的,而真核细胞中的核糖体都是80S的。
()答案:错误解析:真核细胞线粒体和叶绿体内的核糖体的近似于70S。
4. 亲脂性信号分子的受体仅位于细胞核内。
()答案:错误解析:亲脂性波形分子的地处受体也可以位于细胞质基质中。
2、名词解释题(20分,每题5分)1. 细胞分子开关[安徽师范大学2019研]答案:细胞分子开关是通过指激活机制或失活机制精确控制细胞内一系列信号传递级联反应的蛋白质分子,可分为两类:一类分子开关蛋白的活性由磷酸化和去磷酸化控制,蛋白激酶使之磷酸化而开放,蛋白磷酸酯酶使之去吡啶磷酸化而关闭,许多酶由可逆蛋白激酶控制的开关蛋白是蛋白激酶本身;另一类分子开关蛋白是GTP结合蛋白,结合GTP而活化,结合GDP而失活。
解析:空2. pull hypothesis答案:pull hypothesis的中文名称是侧向假说,是指认为染色体向赤道板方向运动微管由于动粒是牵拉的结果的一种假说。
牵拉假说认为动粒微管越长,拉力越大,当来自两级的动粒微管拉力相等时,染色体就被稳定在赤道板上有上用。
解析:空3. 脂筏[北京师范大学2018研]答案:脂筏是脂双层内含有特殊脂质的蛋白质的微区,具有低流动性,呈现有序液相,富含胆固醇和鞘磷脂,形成相对的具有一定功能的筏结构漂浮于二维流动的细胞质膜中。
脂筏的结构与功能3陈 岚 许彩民33 袁建刚 潘华珍(中国医学科学院基础医学研究所中国协和医科大学基础医学院医学分子生物学国家重点实验室,北京100005)摘要 脂筏是膜脂双层内含有特殊脂质及蛋白质的微区.小窝是脂筏的一种类型,由胆固醇、鞘脂及蛋白质组成,以小窝蛋白为标记蛋白.脂筏的组分和结构特点有利于蛋白质之间相互作用和构象转化,可以参与信号转导和细胞蛋白质运转.一些感染性疾病、心血管疾病、肿瘤、肌营养不良症及朊病毒病等可能与脂筏功能紊乱有着密切的关系.关键词 脂筏,小窝,小窝蛋白,信号转导,糖肌醇磷脂2锚固蛋白学科分类号 Q51 自从1972年Nicolson提出膜流动镶嵌结构模型以来,至今有许多关于膜结构的报道,发现膜中有富含胆固醇及鞘脂的微区(microdomain),这个区曾有过许多名称:不溶于去垢剂的糖脂富含区(detergent2ins oluble glycolipid2rich domain,DIFs);富含糖脂的膜(glycolipid2enriched membrane,GEMs);低浓度T riton不溶复合物(low2density triton2ins oluble complex,LDTI).2001年在西班牙召开了欧洲研究讨论会(European Research C onference,EURESCO),会上对“微区、脂筏(lipid raft)及小窝(caveolae)”作了专题的讨论,Meer于次年综述了此专题[1].下面简要介绍小窝及脂筏的结构与功能.1 小窝与小窝蛋白(caveolin)1953年Palade用非离子去污剂提取内皮细胞及平滑肌细胞膜时,发现在低温下用非离子去污剂提取不溶的部分,经浮力密度分离,仍可再分成两部分,较沉的是骨架蛋白,上浮的是膜的另一些组分,从形态观察似在质膜上的囊泡,他设想这种囊泡可游走穿梭在细胞之间,所以起名为质膜囊泡(plasmalemmal vesicles).1955年Y amado报道了与Palade相似的结果,将这种囊泡称小窝(caveolae),是拉丁字小洞(cave)的意思.Y amado所称的小窝即是现在所说的“脂筏”之一,而且是其中最主要的一种.小窝由胆固醇、鞘脂(糖化神经鞘脂、神经鞘磷脂)及蛋白质组成.大约有50~100nm,形态有多种多样,根据其功能不同有瓶型、囊泡型及管型,多数是瓶型.在细胞表面有开放型,如形成胞吐囊泡;也有封闭型,如形成胞吞囊泡.从冷冻刻蚀图观察,可见小窝表面有曲纹覆盖.经分析,曲纹主要由小窝蛋白结合胆固醇而成.小窝的组装分两个步骤:首先在高尔基体由鞘脂及胆固醇形成去污剂不溶的脂质核,内质网合成的糖肌醇磷脂2锚固蛋白(glycosylphosphatidylinosital2anchored protein,GPI2 anchored protein)和小窝蛋白嵌入脂核体,形成初步的组装体,然后运往细胞表面.在此过程中小窝蛋白及胆固醇起关键作用,如果敲除小窝蛋白的基因,即不能形成小窝.胆固醇是在内质网合成,如合成受抑制,也不能形成小窝,胆固醇与小窝蛋白的比例大约在415∶1.所以小窝蛋白可作为小窝的标记蛋白[2]. 小窝蛋白大约至少有4种:小窝蛋白1α、1β、2、3.在膜内,小窝蛋白21易形成同型二聚体;在脂肪细胞中,小窝蛋白21、22共表达,形成异二聚体;小窝蛋白23主要分布在肌肉,其他细胞较少.小窝蛋白的结构中都含有3个半胱氨酸(位于134,144,157)并在此部位进行酰化.小窝蛋白是一种绞架蛋白(scaffolding protein),绞架区基本已清楚,它可直接与胆固醇及鞘脂结合,又可与信息分子(Src2kinase,H2ras,eNOS,G2protein等)结合[3].小窝能募集多种蛋白质可能与此结构有关.2 脂 筏脂筏是指膜脂双层内含有特殊脂质及蛋白质的 3国家自然科学基金重点资助项目(30130070). 33通讯联系人. Tel:010*********,E2mail:caiminxu@ 收稿日期:2002209203,接受日期:2002211201微区,微区内陷可形成囊泡,近年发现脂筏不仅存于质膜,亦可在高尔基体膜上.脂筏的概念早在发现小窝时即有了,经长期的争论,直至1988年Simon 才正式提出“脂筏”之称.脂质的双层有不同的脂筏:外层的微区主要含有鞘脂、胆固醇及GPI 2锚固蛋白,因为鞘脂含有长链饱和脂肪酸,T m 温度较高,流动性差,而且粘稠,邻近的磷脂区其脂肪酸多不饱和,T m 温度较低,所以出现分相;膜内侧也有相似的微区,与外侧的脂质不完全相同,主要是在此区有许多酰化的蛋白质,特别是信号转导蛋白(图1).虽然两层分别有脂筏,但它们是偶联的,因为用非离子去污剂提取时,不仅有外层的GPI 2锚固蛋白,还有许多信息分子共同被提出.用一种GPI 2锚固蛋白的抗体介导锚固蛋白聚集,与此同时Src 家族的酪氨酸激酶也被激活.如换成糖鞘脂的抗体,也有同样的现象.GPI 蛋白及糖鞘脂都存于膜外侧,Src 酪氨酸激酶在膜内侧,这表明脂筏内外层之间是有联系的[4].Fig 11 Model of lipid rafts图1 脂筏模式图 用不同的去污剂或改变溶解时的温度,所得到的脂质及蛋白质都有差异,说明细胞膜上的脂筏并不都等同.脂筏可能有三类:小窝、富含糖鞘脂膜区(glycosphingolipid enrich membrane )、富含多磷酸肌醇(PIP2)膜区.不同的脂筏有其各自的特异蛋白,并有不同的功能.脂筏的部分脂质及蛋白质的组分见表1.T able 1 partial lipid and protein components of lipid raft [5]表1 脂筏的部分脂质及蛋白质的组成[5]脂质蛋白质受体蛋白胆固醇、鞘脂、糖鞘脂(如GM1)、PIP2、花生四烯酸小窝蛋白、flotillins 、stomatins 、MAL/BENE 、LAT/PAG 、VIP36、GPI 2锚固蛋白类(如叶酸受体,疯牛病蛋白,碱性磷酸酶等)、非受体酪氨酸激酶(Src 2家族)、G蛋白、一氧化氮合酶(eNOS )、Ras 、Fyn 、PKCα受体酪氨酸激酶(RTKs ,如EGF 2R 、PD GF 2RInsulin 2R 等) 脂筏内的蛋白质,有的是经跨膜直接插入膜,但更多的蛋白质需酰化,由酰化后的脂肪酸插入膜.Zacharias [6]的实验表明,用水母蛋白的两个变异体,分别融合蓝荧光蛋白(CFP )及绿荧光(YEP )蛋白,然后在多肽链上分别联接脂化(棕榈酸、豆蔻酸、异戊二烯)的特异共同序列,用荧光能量共振转移(fluorescence resonance energy transfer )测定邻近效应,发现棕榈酰化及豆蔻酰化的蛋白质在脂筏的小窝区,而异戊二烯化的蛋白质形成二聚体,不在小窝区,对胆固醇不敏感,可能是另一类的脂筏.说明不同的酰化蛋白插入不同的脂筏. Stuermer 在神经元细胞膜内,发现两个新蛋白质———flotillin 21及flotillin 22,其不溶于去污剂,与脂质形成脂筏.用其抗体进行免疫组化染色,在光镜及电镜下可见,flotillin 21,2排列在细胞膜的胞浆面,在延伸的轴突及丝足上都存在.它不仅存于神经细胞,而且在人Jurkat T 2淋巴细胞株的细胞上也可被发现,并且与Thy21,Fyn蛋白交联,参与淋巴细胞活化,如加入Thy21抗体,可见与flotillin21,2共聚,但在这个脂筏内没有小窝蛋白,这说明,含有flotillin的结构可组成另一类的脂筏.Salzer用钙诱导红细胞时,从形态及生化的分析发现有两种囊泡:一种称微囊泡(microvesicle),直径约150nm;另一种称纳诺囊泡(nanovesicle),直径约60nm.两种囊泡除都共有GPI2锚固蛋白外,其他的组分不完全相同.微囊泡含有两种蛋白质:stomatin及flotillin21,2.Stomatin是棕榈酰化蛋白质,以棕榈酸插入脂筏区,作用很像小窝蛋白.Flotillin21,2也在脂筏区,但它没有被酰化,在囊泡及质膜都存在.纳诺囊泡还有另两种蛋白质———会联蛋白(synexin)及抗药蛋白(s orcin).会联蛋白是膜联蛋白(annexin)家族的成员annexinⅦ(可介导分泌小泡的聚集),负责细胞内运转及膜的组装.它与钙结合后,转到膜上形成膜融合蛋白.抗药蛋白也可与钙结合,结合后,暴露出疏水基团,直接插入膜,有观点认为抗药蛋白与会联蛋白都在脂筏上,但分别在不同的区域或两者结合后进入脂筏.这说明在红细胞的囊泡中也有脂筏[7].3 脂筏的功能从结构及组分分析,脂筏有两个特点:a1许多蛋白质聚集在脂筏内,便于相互作用.b1脂筏的环境有利于蛋白质的变构,形成有效的构象.所以它具有许多功能.311 参与信号转导由于脂筏内有多种信号分子,它可以参与许多的信号转导通路.以T细胞为例:T细胞膜上有T 细胞抗原抗体(TCR),当它活化后可引起许多信息分子聚集形成复合体.最近Zhang[8]报道这种复合体形成的关键是脂筏提供了平台.在静止的T 细胞脂筏内富含Src家族激酶Lck、Fyn及跨膜的衔接子(linker activated T cell,LA T),还有少量的CD4和CD3,其中最重要的是LA T.它是一个跨膜蛋白,是伸向胞浆的一段肽链,富含酪氨酸,另外LA T的半胱氨酸还可酰化棕榈酸,以此插入脂筏.当TCR被激活,Src酪氨酸激酶活化引起LA T的多个酪氨酸被磷酸化,然后,LA T募集多种底物进行磷酸化,包括ZAP70、PLCγ1、PI3K、Vav,形成活化的大复合体.这些反应都必须在脂筏内完成,如果LA T不能进入脂筏,T2细胞便不能执行其重要的增殖及分化功能.其他免疫识别受体,如B细胞抗原受体(BCR)、肥大细胞高亲和IgE受体(FcζRI)等也都以脂筏作为传递信息的场所.Roy[9]报道调节小窝蛋白的表达,可直接影响信息蛋白分子(包括癌基因).他发现小窝蛋白21的启动子有胆固醇的调节位点,小窝蛋白在小窝内有运转及调节胆固醇水平的作用,所以调节小窝蛋白的表达,可以间接影响信息的传递.Parton制备了一个小窝蛋白的变异体(gavD GV),可抑制H2Ras介导Raf的活化,如加入胆固醇可改善抑制作用.他设想H2Ras是Ras的异构体,有棕榈酸化,需要富有胆固醇的脂筏才能活化Raf,这个筏区被小窝蛋白变异体破坏.HIa[10]发现在细胞质膜有溶血卵磷脂类(L Ps)的受体,这种受体是一个大家族,大约有12个左右,它与G蛋白偶联,称G蛋白偶联受体(G protein2coupled receptors,GPCRs).L Ps受体位于膜富含鞘磷脂的小窝区.L Ps包括溶脂酸(L PA)、溶血卵磷脂(L PC)、鞘磷酸胆碱(SPC)及鞘磷脂212磷酸(S2I2P).L Ps受体的作用包括:参与调节细胞迁移、增殖及存活;调节血管系统的成熟;激活eNOS及活化磷脂酶C(P KC)等.Nanjundan[11]从鼠肺细胞内提取出一种脂质磷酸水解酶(LL P),活化的LL P存于小窝内,它可水解L PA、PA和鞘氨醇212磷酸,水解后的产物皆为第二信使.312 参与细胞蛋白运转31211 参与跨细胞运转:文献[2]报道,分布在内皮细胞的小窝囊泡,开口于细胞的表面,细胞与细胞之间的囊泡可形成一个通道,小分子物质可相互交换,小窝成为一些小分子化合物运转的通路. 31212 参与细胞胞饮作用:小窝内包含许多受体蛋白,小窝将它们运送到细胞内可利用不同的途径:a1受体与配基结合后,被小窝囊泡运送到胞浆,配基与受体分离,受体随囊泡回到质膜;b1小窝囊泡载着受体与配基,通过胞浆,在细胞的反方向开口将配基释放到细胞外基质.有些毒素(如霍乱毒素)与细胞上的受体结合,也以这种方式进入宿主细胞内[2].31213 参与细胞分选:Martin[12]介绍了一个膜内在蛋白MAL.MAL属酰化蛋白质家族,存于Madin2 Darby犬肾细胞(Madin2Darby canine kidney, MDCK),有多个疏水基团插入极化的上皮细胞(polarized epithilia)的脂筏.MAL直接参与极化分选(apical s orting).在MDCK细胞内如加入MAL反义寡核苷酸,就无分选功能.MAL即像一个货车,可在质膜、内质网、高尔基体之间循环,运送蛋白.另外BENE蛋白,和MAL一样,也属酰化蛋白质家族.它与小窝蛋白21结合,位于小窝内,可能这两个蛋白质是脂筏的新成员.Millan[13]报道当T2淋巴细胞执行其功能时,发生极化形态.细胞的一端延伸,形成特殊的突起———“尾足”(uropod),随后选择性地将一些粘附分子(ICAM21,ICAM22,ICAM23,CD43,CD44)浓缩在尾足的脂筏区.血粘素(hemaglutinine,HA)在T淋巴细胞的高尔基体合成后,进入脂筏囊泡,随囊泡也被送到尾足的表面,说明尾足富含脂筏.细胞的另一端称前缘,对抗原及细胞因子很敏感,其复杂的免疫反应都在前缘脂筏区进行.4 脂筏与疾病脂筏的生理功能具有多向性,如果它的正常生理过程发生改变,则会引发多种疾病.近来由于一些小窝蛋白基因缺陷的动物模型的建立,为人们理解和发现与脂筏相关的疾病创造了条件.411 脂筏与感染性疾病细胞内吞作用可以发生在小窝结构上.许多病原体如病毒、寄生虫、细菌及其毒素都能经由小窝进入宿主细胞.这些病原体或毒素通过小窝的内化作用进入宿主细胞后,并不与溶酶体融合(与溶酶体融合导致内吞物的降解是经典细胞内吞作用的途径),这样,外源物质就可以避开机体的免疫反应和一系列的清除行为,得以在体内生存并发挥其致病作用.Abraham等[14]向人们展示了细菌在小鼠骨髓源性肥大细胞(mouse bone marrow2derived mast cells,BMMCs)上经由小窝包裹形成囊后进入噬菌细胞的机制.Oppenheim等的实验说明了SV40经由小窝内吞后,未与溶酶体结合,而转运至内质网的特殊感染途径.用药物阻断这一通路中某些蛋白质的组装,就可以抑制感染的发生[15,16].目前还有待在体内实验和分子水平上阐述小窝在感染性疾病中发挥的作用,借此,人们将利用更为合理的传送途径来设计药物以达到更为有效的阻断病原体感染的治疗目标.412 脂筏与心血管疾病内皮来源的一氧化氮生物利用率低下,在动脉粥样硬化症发生过程中起着关键作用.内皮型一氧化氮合成酶(eNOS)直接受到小窝蛋白的调节,与小窝关系密切.实验表明,利用氧化型的低密度脂蛋白作为胆固醇的接收体,可以导致小窝的耗竭,也可以抑制由乙酰胆碱诱导的eNOS的活性,但不影响eNOS的数量和环前列腺素的产量,也不影响eNOS的肉豆蔻酰化、棕榈酰化和磷酸化.去除氧化型低密度脂蛋白,小窝的结构可以得到恢复,并且eNOS亦可再与小窝结合[17].Bucci等在体内实验中证实了小窝蛋白21对eNOS信号通路起着首要的作用,一旦eNOS通路出现抑制,则会消除由NO介导的血管渗透和收缩反应.随后, Razani[18]和Drab两个研究小组利用敲除小窝蛋白2 1基因的小鼠,在离体大动脉血管收缩紧张度测量的实验中发现,小窝蛋白21的缺乏导致eNOS活性水平低并导致血管弹性降低,在动脉血管中也发现了这一现象.另外,糖化终末产物(advanced glycation end products,A GEs)通过其受体介导,在糖尿病并发症中发挥着重要的作用.Stitt等[19]发现,在视网膜微循环内皮细胞中,A GEs与其受体的相互作用是在小窝中进行的.由此表明,小窝的内化及信号转导功能或许可以调控A GE受体介导的血管功能障碍.413 脂筏与肿瘤许多原癌基因和抑癌基因是信号转导通路中处于不同阶段的蛋白质,当它们的生理功能发生改变或缺失时就会促进肿瘤形成.脂筏可以参与信号转导.由此自然让人联想到,脂筏结构和功能的变化与肿瘤发生有密切的关系.小窝蛋白21基因被作为一种候选的抑癌基因.小窝蛋白21的mRNA和蛋白质表达水平在肿瘤转化的N IH23T3培养细胞中出现下调,在乳腺癌转基因小鼠中以及其他起源于人肿瘤的细胞系中这种基因的表达也出现下调[20].一般来说,确定小鼠的一种原癌基因或抑癌基因需要分析原始鼠胚胎纤维原细胞的生长特性,因此,在对小窝蛋白21进行了细胞增殖和细胞周期评估后,发现小窝蛋白21敲除的小鼠胚胎纤维原细胞出现细胞周期整体活性增加,主要出现合成期的增加[18].Pramudji等[21]利用腺病毒载体介导小窝蛋白21启动子对原位前列腺癌动物模型进行基因治疗,结果出现新生物的坏死.肿瘤抗药性是导致对癌症患者化疗失败的原因.研究发现多药耐药(multidrug resistant, MDR)的癌细胞可表达高水平和高密度的小窝[22].这给人们了解多药效耐药细胞的产生机制和设计有针对性的药物起到了指导作用.414 脂筏与肌营养不良症小窝蛋白21和小窝蛋白22广泛分布在组织细胞上,而小窝蛋白23仅仅局限地分布于肌肉组织,这一点显现出小窝蛋白23可能在肌肉组织功能障碍中起到某些作用.杜兴肌营养不良(Duchenne muscular dystrophy, DMD)是一种严重的肌肉功能紊乱疾病,其特征是缺乏了肌营养不良蛋白.肌营养不良蛋白是一种细胞内的蛋白质,是DMD基因的表达产物,可以联系细胞内的细胞骨架和细胞外基质,这一作用对于保持正常肌肉功能和肌肉纤维的完整性是极为必要的.免疫电镜观察到肌营养不良蛋白局限在肌肉细胞的小窝上[23].还有一些形态学和生物化学的观察发现,在DMD病理过程中有小窝和小窝蛋白23的参与.很早的时候,人们利用电镜和冰冻蚀刻技术在DMD患者的骨骼肌中发现小窝数目的增多.与此结果相一致的是,最近,在mdx小鼠(一种肌营养不良蛋白缺陷的DMD动物模型)和患者的骨骼肌中发现,小窝蛋白23的表达增高了2~3倍.在超表达小窝蛋白23的转基因小鼠中,表现出杜氏肌营养不良样症状[24].这种转基因模型的建立,有助于人们更好地研究和理解人类DMD的病理发生机制.肌带型肌营养不良(limb2girdle muscular dystrophy,L GMD21C)是另一种肌营养不良症,是由于小窝蛋白23基因突变造成的,这种突变基因编码的蛋白质很不稳定,易被降解,所以存在甚少.G albiati等[25]模拟人的基因突变点,构建缺失小窝蛋白23基因的小鼠,证实小窝蛋白23是形成肌细胞内小窝的主体.正常动物肌细胞内小窝蛋白23与肌营养不良蛋白及糖蛋白形成复合体,存于脂筏.基因突变的小鼠,无脂筏,所以不能形成复合体.说明小窝蛋白23的突变是产生L GMD21C的分子基础.由此可见,正常生理条件下,小窝蛋白的表达是一个动态平衡的过程.415 脂筏与朊病毒病朊病毒病包括人克雅氏病(Creutzfeldt2Jakob Disease,C JD)、羊瘙痒症和牛海绵样脑病等,其发病原因是细胞朊蛋白(prion protein,PrP C)的构象转变为异常形式(PrP SC).PrP C是结合在细胞膜外表面,带有GPI锚结构的糖蛋白.研究表明,小鼠的神经瘤母细胞(N2a)的PrP C聚集在小窝样结构域中,经scrapie感染后N2a(ScN2a)的PrP SC也聚集在相应的结构中.细胞胆固醇水平的降低可以抑制PrP C向PrP Sc的转化.这就提示人们,富含胆固醇的小窝样结构域很可能就是朊病毒传播增殖的场所.Kaneko等[26]证实PrP Sc是由GPI 锚固形式的PrP C转变而来的,并不是来自跨膜的PrP C.由于GPI是小窝结构的标志性蛋白质之一,这也证实了上述实验的结论.416 脂筏与肺部疾病肺泡Ⅰ型上皮细胞富含小窝蛋白21.在敲除小窝蛋白21基因的动物中,肺泡空间明显变得狭小,肺泡间隔由于不可控制的上皮细胞增殖和纤维化而变厚,导致严重的肺功能紊乱,这种小窝蛋白21基因敲除的小鼠在游泳实验中很容易疲劳[18].5 结束语人们对脂筏结构与功能的研究由来已久,特别是近几年虽然有许多问题已较清楚,但仍存在不少有待深入研究的问题:a1膜内外层的脂筏组分有何异同,特别是内膜很难提取,至今对其组分还不清楚;b1蛋白质如何识别不同的脂筏,进入一个有效区;c1脂筏内跨膜蛋白与脂筏内的酰化蛋白之间如何相互作用;d1脂筏在膜内如何游走,虽然对脂筏与细胞骨架关系的了解有了良好的开端,但是尚缺乏更深入的研究.期望应用先进的技术,解决在无试剂干扰的条件下,观察天然单细胞膜脂与膜蛋白的活动.从对小窝蛋白的研究,可看到它与癌症、动脉粥样硬化、糖尿病并发症、早老性痴呆、疯牛病及肌营养不良等疾病的关系.同时,对脂筏结构和功能的研究将会给生物学及医学带来更多的信息与启示.参 考 文 献1 Meer G V.The different hues of lipid rafts.Science,2002,296 (5569):855~8572 Anderson R G W.The caveolae membrane system.Ann Rev Biochen,1998,67:199~2253 Massimino M L,Griffoni C,Spisni E,et al.Involvement of caveolae and caveolae2like domain in signaling cell survival and angiogenesis.Cell Signal,2002,14(2):93~984 Brown D A,London E.Structure and function of sphingolipid and cholesterol2rich membrane rafts.J Biol Chem,2000,275(23): 17221~172245 G albiati F,Razani B,Lisanti M P.Emerging themes in lipid rafts and caveolae.Cell,2001,106(4):403~4116 Zacharias D A,Violin J D,Newton A C,et al.Partitioning of lipid2modified monomeric GFPs into membrane microdomains of live cells.Science,2002,296(5569):913~9167 Salzer U,Hinterdorfer P,Hunger U,et al.Ca2+dependent vesicle release from erythrocytes involves stomatin2specific lipid rafts,synexin(annexinⅦ),and sorcin.Blood,2002,97(7): 2569~25778 Zhang W,Sloan2Lancaster J,K itchen J,et T:the ZAP270 tyrosine kinase substrate that links T cell receptor to cellular activation.Cell,1998,92(1):83~929 Roy S,Luetterforst R,Harding A,et al.Dominant2negative caveolin inhibits H2Ras function by disrupting cholesterol2rich plasma membrane domains.Nat Cell Biol,1999,1(2):98~10510Hla T,Lee M J,Ancellin N,et al.Lysophospholipids2receptor revelations.Science,2001,294(5548):1875~187811Nanjundan M,Possmayer F.Pulmonary lipid phosphate phosphohydrolase in plasma membrane signaling platforms.Biochem J,2001,358(Pt3):637~64612Martin2Belmonte F,Puertollano R,Millan J,et al.The MAL proteolipid is necessary for the overall apical delivery of membrane proteins in the polarized epithelial Madin2Darby canine kidney and fischer rat thyroid cell lines.Mol Biol Cell,2000,11(6):2033~204513Millan J,Montoya M C,Sancho D,et al.Lipid rafts mediate biosynthetic transport to the T lymphocyte uropod subdomain and are necessary for uropod integrity and function.Blood,2002,99(3):978~98414Shin J S,Abraham S N.Caveolae2not just craters in the cellular landscape.Science,2001,293(5534):1447~144815Norkin L C,Anderson H A,Wolfrom S A,et al.Caveolar endocytosis of simian virus40is followed by brefeldin A2sensitive transport to the endoplasmic reticulum,where the virus disassembles.J Virol,2002,76(10):5156~516616Richards A A,Stang E,Pepperkok R,et al.Inhibitors of COP2 mediated transport and cholera toxin action inhibit simian virus40 infection.Mol Biol Cell,2002,13(5):1750~176417Alison B,Philip W S,Ivan Y,et al.Smart oxidized low densitylipoprotein displaces endothelial nitric2oxide synthase(eNOS)from plasmalemmal caveolae and impairs eNOS activation.J Biol Chem, 1999,274(45):32512~3251918Razani B,Engelman J A,Wang X B,et al.Caveolin21null mice are viable,but show evidence of hyper2proliferative and vascular abnormalities.J Biol Chem,2001,276(41):38121~38138 19Stitt A W,Burke G A,Chen F,et al.Advanced glycation end2 product receptor interactions on microvascular cells occur within caveolin2rich membrane domains.FASEB J,2000,14(15): 2390~239220Razani B,Schlegel A,Lisanti M P.Caveolin proteins in signaling, oncogenic transformation and muscular dystrophy.J Cell Sci, 2000,113(Pt12):2103~210921Pramudji C,Shimura S,Ebara S,et al.Timme in situ prostate cancer gene therapy using a novel adenoviral vector regulated by the caveolin21promoter.Clin Cancer Res,2001,7(12):4272~427922Lavie Y,Fiucci G,Liscovitch M.Up regulation of caveolae and caveolar constituents in multidrug resistant cancer cells.J Biol Chem,1998,273(49):32380~3238323G albiati F,Razani B,Lisanti M P.Caveolae and caveolin23in muscular dystrophy.Trends Mol Med,2001,7(10):435~44124G albiati F,Volont D,Chu J B,et al.Transgenic overexpression of caveolin23in skeletal muscle fibers induces a Duchenne2like muscular dystrophy phenotype.Proc Natl Acad Sci USA,2000, 97(17):9689~969425G albiati F,Engelman J A,Volonte D,et al.Cavelin23null mice show a loss of caveolar,changes in the microdomain distribution of the dystrophin glycoprotein complex,and T2tubule abnormalities.J Biol Chem,2001,276(24):21425~2143326Kaneko K,Vey M,Scott M,et al.COOH2terminal sequence of the cellular prion protein directs subcellular trafficking and controls conversion into the scrapie isoform.Proc Natl Acad Sci USA, 1997,94(6):2333~2338Structure and Function of Lipid R afts3CHEN Lan,XU Cai2Min33,YUAN Jian2G ang,PAN Hua2Zhen(N ational L aboratory of Medical Molecular Biology,Basic Instit ute of Medical Sciences,The Chi nese Academy of Medical Sciences,Peki ng U nion Medical College,Beiji ng100005,China)Abstract Lipid rafts are liquid2ordered membrane microdomains with a unique protein and lipid composition found on the plasma membrane.Caveolae,a type of lipid rafts,is characterized by high levels of cholesterol, sphingolipids and proteins,and is identified by the presence of the protein caveolin.The structure and component of lipid rafts is used of reciprocity and comformational change between proteins.Lipid rafts are associated with signal transduction and cellar proteins movement.The disfunction of lipid rafts is related to some diseases such as infection,heart disease,cancer,muscular dystrophy and prion protein diseases.K ey w ords lipid raft,caveolae,caveolin,signal transduction,GPI2anchored protein 3This work was supported by a grant from The National Natural Sciences Foundation of China(30130070). 33Corresponding author.Tel:86210265296445,E2mail:caiminxu@ Received:September3,2002 Accepted:November1,2002。