pikakt信号通路图谱
- 格式:docx
- 大小:1.19 MB
- 文档页数:2

经典信号通路之PI3K-AKT-mTOR信号通路磷脂酰肌醇3-激酶(PI3Ks)信号通路相关磷脂酰肌醇3-激酶(PI3Ks)蛋白家族参与细胞增殖、分化、凋亡和葡萄糖转运等多种细胞功能的调节。
PI3K活性的增加常与多种癌症相关。
PI3K磷酸化磷脂酰肌醇PI(一种膜磷脂)肌醇环的第3位碳原子。
PI在细胞膜组分中所占比例较小,比磷脂酰胆碱、磷脂酰乙醇胺和磷脂酰丝氨酸含量少。
但在脑细胞膜中,含量较为丰富,达磷脂总量的10%。
PI的肌醇环上有5个可被磷酸化的位点,多种激酶可磷酸化PI肌醇环上的4th和5th位点,因而通常在这两位点之一或两位点发生磷酸化修饰,尤其发生在质膜内侧。
通常,PI-4,5-二磷酸(PIP2)在磷脂酶C的作用下,产生二酰甘油(DAG)和肌醇-1,4,5-三磷酸。
PI3K转移一个磷酸基团至位点3,形成的产物对细胞的功能具有重要的影响。
譬如,单磷酸化的PI-3-磷酸,能刺激细胞迁移(cell trafficking),而未磷酸化的则不能。
PI-3,4-二磷酸则可促进细胞的增殖(生长)和增强对凋亡的抗性,而其前体分子PI-4-磷酸则不然。
PIP2转换为PI-3,4,5-三磷酸,可调节细胞的黏附、生长和存活。
PI3K的活化PI3K可分为3类,其结构与功能各异。
其中研究最广泛的为I类PI3K, 此类PI3K为异源二聚体,由一个调节亚基和一个催化亚基组成。
调节亚基含有SH2和SH3结构域,与含有相应结合位点的靶蛋白相作用。
该亚基通常称为p85, 参考于第一个被发现的亚型(isotype),然而目前已知的6种调节亚基,大小从50至110kDa不等。
催化亚基有4种,即p110α, β,δ,γ,而δ仅限于白细胞,其余则广泛分布于各种细胞中。
PI3K的活化很大程度上参与到靠近其质膜内侧的底物。
多种生长因子和信号传导复合物,包括成纤维细胞生长因子(FGF)、血管内皮生长因子(VEGF)、人生长因子(HGF)、血管位蛋白I(Ang1)和胰岛素都能启始PI3K的激活过程。

Akt信号转导通路课件一、引言信号转导是细胞生物学研究的重要领域之一,通过信号转导,细胞可以感知外部环境的变化,并作出相应的生物学反应。
Akt信号转导通路是细胞信号转导的重要组成部分,对于细胞的生长、增殖、分化和生存等方面具有重要作用。
本文将对Akt信号转导通路进行详细介绍。
二、Akt信号转导通路的基本概念1.Akt信号转导通路简介Akt信号转导通路,又称为蛋白激酶B(ProteinKinaseB,PKB)信号转导通路,是细胞内重要的信号转导途径之一。
Akt信号转导通路在细胞的生长、增殖、分化和生存等方面具有重要作用。
2.Akt信号转导通路的主要组成部分(1)Akt蛋白:Akt蛋白是Akt信号转导通路的核心组成部分,具有蛋白激酶活性。
(2)PI3K(磷脂酰肌醇3激酶):PI3K是Akt信号转导通路的上游分子,能够将磷脂酰肌醇4,5二磷酸(PIP2)转化为磷脂酰肌醇3,4,5三磷酸(PIP3)。
(3)PDK1(磷脂酰肌醇依赖性激酶1):PDK1是Akt信号转导通路中的重要分子,能够磷酸化Akt蛋白,激活其激酶活性。
(4)mTOR(哺乳动物雷帕霉素靶蛋白):mTOR是Akt信号转导通路的重要下游分子,能够调控细胞的生长、增殖和分化等生物学过程。
三、Akt信号转导通路的作用机制1.Akt的激活Akt的激活主要依赖于PI3K的激活。
当细胞受到外部信号的刺激时,PI3K被激活,将PIP2转化为PIP3。
PIP3能够招募Akt蛋白到细胞膜上,并使其磷酸化激活。
2.Akt的生物学功能(1)细胞增殖:Akt能够磷酸化细胞周期蛋白依赖性激酶(CDKs),促进细胞周期的进程,从而促进细胞增殖。
(2)细胞生存:Akt能够磷酸化促凋亡蛋白BAD,抑制其促凋亡作用,从而促进细胞生存。
(3)蛋白质合成:Akt能够激活mTOR信号通路,促进蛋白质的合成。
四、Akt信号转导通路在疾病中的作用1.癌症Akt信号转导通路在癌症的发生和发展中具有重要作用。


信号通路3 —PI3K/AKT/mTORAPExBIO一、PI3K/Akt/mTORPI3K/AKT/mTOR是调节细胞周期的重要细胞内信号通路。
PI3K/AKT/mTOR信号通路与细胞的休眠、增殖、癌变和寿命直接相关。
PI3K激活后磷酸化并激活AKT,将其定位在质膜中。
信号通过AKT传递到下游不同的靶点,如激活CREB,抑制p27,将FOXO定位于细胞质中,激活PtdIns-3ps,及激活mTOR(影响p70或4EBP1的转录)。
该通路的激活因子包括EGF、shh、IGF-1、胰岛素和CaM。
该信号通路的拮抗因子,包括PTEN、GSK3B、和HB9。
在多种癌症中,PI3K/AKT/mTOR通路是过度活化的,因此减少凋亡并促进增殖。
然而,该通路在成人干细胞尤其是神经干细胞的分化过程中促进细胞生长和增殖。
1. PI3KPhosphatidylinositide 3-kinases,是一种胞内磷脂酰肌醇激酶。
由调节亚基p85和催化亚基p110构成。
与v.sre和v.ras等癌基因的产物相关。
PI3K本身具有丝氨酸/苏氨酸(Ser/Thr)激酶的活性,也具有磷脂酰肌醇激酶的活性。
2. Akt又称PKB(protein kinase B)。
是一种丝氨酸/苏氨酸特异性蛋白激酶,在多种细胞生长过程中发挥关键作用,如葡萄糖代谢、凋亡、细胞增殖、转录和细胞迁移。
Akt的Ser473可以被PDK1磷酸化。
PKB与PKA和PKC均有很高的同源性,该激酶被证明是反转录病毒安基因v-akt 的编码产物,故又称Akt。
3. mTORMammalian target of rapamycin。
mTOR与其它蛋白质结合,形成两种不同蛋白质复合物,mTOR复合物1(mTORC1,)和mTOR复合物2(mTORC2),它们调节不同的细胞过程。
mTORC1由mTOR、mTOR调节相关蛋白Raptor、MLST8和非核心组分PRAS40、DEPTOR 组成。

解读Akt(上)丨Akt磷酸化位点那么多,到底该选哪一个?摘要教你如何选择Akt各磷酸化位点,以及它们的功能及区别。
正文Akt家族简介Akt也被称为蛋白激酶B (protein kinase B,PKB),是一种分子量约为60kDa的丝/苏氨酸蛋白激酶,存在于哺乳动物中的三种同工型。
AKT可分为3种亚型(AKT1、AKT2、AKT3或PKBα, PKBβ, PKBγ),3种亚型的功能各异,但也有重叠。
AKT1 广泛地分布在组织中,Akt2主要分布在肌肉和脂肪细胞中,而Akt3则在睾丸和大脑中表达。
Akt 调控多个生物过程,包括细胞存活、增殖、生长和糖原代谢,Akt信号通路与恶性肿瘤、糖尿病、类风湿关节炎等多种疾病的发生发展密切相关,越来越受到广泛的关注【1-3】。
许多生长因子、激素与细胞因子通过结合其同源受体酪氨酸激酶(RTK)、细胞因子受体或GPCR,以及通过触发脂质激酶PI3K 的活化来激活Akt,从而在细胞质膜中生成PIP3。
Akt 通过其普列克底物蛋白同源 (PH) 结构域与 PIP3 相结合,导致 Akt 转位至细胞膜。
通过双磷酸化机制,可以激活Akt。
同样因为自身PH 结构域而转位至细胞膜的 PDK1,能够活化Akt的Thr308 位点使 Akt 磷酸化,PDK2再对Akt的Ser473 位点磷酸化才能最后导致其完全活化,也就是说羧基末端Ser473 位点的二次磷酸化对于活性来说也是必要的,并由mTOR-rictor 复合体 mTORC2 完成, 活化的Akt引起下游磷酸化级联反应和靶蛋白之间的相互作用详见图1 [4]。
图1: PI3K-Akt信号通路简图。
让我们来看下Akt的表达定位,一般在静止的细胞是位于细胞浆的,在被生长因子等激活之后,Akt由细胞浆转移至细胞膜内测,活化后的AKT进入细胞浆和细胞核,通过使下游底物蛋白磷酸化而激活(如图2)。
图2:使用 Phospho-Akt (Ser473) (D9E) XP® Rabbit mAb(绿色),对经 LY294002 处理(左)或经胰岛素处理(右)的 C2C12 细胞进行共聚焦免疫荧光分析。
PI3K/AKT/mTORPI3K/AKT/mTOR是调节细胞周期的重要细胞内信号通路。
PI3K/AKT/mTOR信号通路与细胞的生长、存活、增殖、凋亡、血管生成、自吞噬过程中发挥着重要的生物学功能。
该通路是由磷脂酰肌醇3- 激酶(PI3Ks)、丝氨酸/苏氨酸蛋白激酶(Akt)和哺乳动物雷帕霉素靶蛋白(mTOR)组成。
PI3K/Akt/mTOR通路过程PI3K激活后产生PIP3, PIP3促使PDK1(phosphoinositide dependent kinase-1)磷酸化含有PH结构域的信号AKT蛋白(Ser308),从而活化AKT。
AKT有很多下游效应,可通过磷酸化多种酶、激酶和转录因子等下游因子,进而调节细胞的功能。
mTOR,是PI3K/Akt 下游的一种重要的丝氨酸-苏氨酸蛋白激酶,调节肿瘤细胞的自噬的经典通路。
PI3K/Akt/mTOR信号通路图按靶点分类:*PI3KPI3K,是一种胞内磷脂酰肌醇激酶,也具有丝氨酸/苏氨酸(Ser/Thr)激酶的活性。
能够通过PI3K诱发PIP3生成的激活因子,则能够激活Akt 信号途径,包括受体酪氨酸激酶、整合素、B 细胞和T 细胞受体、细胞因子受体、G 蛋白偶联受体等等。
*Akt又称PKB或Rac,是一种丝氨酸/苏氨酸特异性蛋白激酶B,在细胞存活和凋亡中起重要作用,如葡萄糖代谢、凋亡、细胞增殖、转录和细胞迁移。
Akt的Thr308可以被PDK1磷酸化,而被部分激活。
或者473位点上的丝氨酸被mTORC2磷酸化,激发Akt的完全酶活性。
*mTORmTOR是细胞生长和增殖的重要调节因子。
mTOR与其它蛋白质结合,形成两种不同蛋白质复合物,mTORC1和mTORC2,参与调节不同的细胞过程。
*GSK-3。

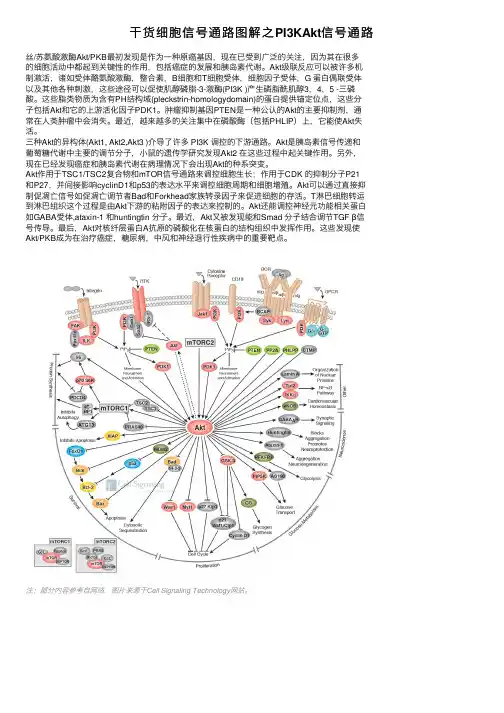
⼲货细胞信号通路图解之PI3KAkt信号通路丝/苏氨酸激酶Akt/PKB最初发现是作为⼀种原癌基因,现在已受到⼴泛的关注,因为其在很多的细胞活动中都起到关键性的作⽤,包括癌症的发展和胰岛素代谢。
Akt级联反应可以被许多机制激活,诸如受体酪氨酸激酶,整合素,B细胞和T细胞受体,细胞因⼦受体,G 蛋⽩偶联受体以及其他各种刺激,这些途径可以促使肌醇磷脂-3-激酶(PI3K )产⽣磷脂酰肌醇3,4,5 -三磷酸。
这些脂类物质为含有PH结构域(pleckstrin-homologydomain)的蛋⽩提供锚定位点,这些分⼦包括Akt和它的上游活化因⼦PDK1。
肿瘤抑制基因PTEN是⼀种公认的Akt的主要抑制剂,通常在⼈类肿瘤中会消失。
最近,越来越多的关注集中在磷酸酶(包括PHLIP)上,它能使Akt失活。
三种Akt的异构体(Akt1, Akt2,Akt3 )介导了许多 PI3K 调控的下游通路。
Akt是胰岛素信号传递和葡萄糖代谢中主要的调节分⼦,⼩⿏的遗传学研究发现Akt2 在这些过程中起关键作⽤。
另外,现在已经发现癌症和胰岛素代谢在病理情况下会出现Akt的种系突变。
Akt作⽤于TSC1/TSC2复合物和mTOR信号通路来调控细胞⽣长;作⽤于CDK 的抑制分⼦P21和P27,并间接影响cyclinD1和p53的表达⽔平来调控细胞周期和细胞增殖。
Akt可以通过直接抑制促凋亡信号如促凋亡调节者Bad和Forkhead家族转录因⼦来促进细胞的存活。
T淋巴细胞转运到淋巴组织这个过程是由Akt下游的粘附因⼦的表达来控制的。
Akt还能调控神经元功能相关蛋⽩如GABA受体,ataxin-1 和huntingtin 分⼦。
最近,Akt⼜被发现能和Smad 分⼦结合调节TGF β信号传导。
最后,Akt对核纤层蛋⽩A抗原的磷酸化在核蛋⽩的结构组织中发挥作⽤。
这些发现使Akt/PKB成为在治疗癌症,糖尿病,中风和神经退⾏性疾病中的重要靶点。


干货细胞信号通路图解之细胞代谢相关通路(1)Insulin Receptor Signaling:胰岛素是控制关键的能量代谢功能如葡糖和脂代谢过程中的主要激素。
胰岛素激活胰岛素受体酪氨酸激酶,后者磷酸化并召集不同的底物接头蛋白如IRS家族蛋白。
IRS上酪氨酸被磷酸化后成为众多信号传导关联蛋白的结合位点。
其中,PI3K在胰岛素功能中起主要的作用,大部分是通过活化Akt/PKB和PKCζ级联来完成的。
活化的Akt通过抑制GSK-3诱导糖原的合成,通过mTOR和下游元件影响蛋白合成,通过抑制几个促凋亡分子(Bad,Forkhead家族转录因子,GSK-3)影响细胞生存。
胰岛素促进肌肉和脂肪细胞对葡糖的吸收,这是因为它能促使还有GLUT4的囊泡转移到细胞膜上。
GLUT4的移位需要PI3K/Akt通路的参与和IR介导的CAP 的磷酸化和形成CAP:Cbl:CrkII复合体。
胰岛素信号通路也会影响细胞生长和有丝分裂,主要是通过Akt级联进行,也会有Ras/MAPK通路的参与。
另外,胰岛素信号通路可以通过中断CREB/CBP/Torc2的结合抑制肝脏中的糖异生。
胰岛素信号通路还可以通过激活SREBP-1C,USF1和LXR来促进脂肪酸的合成。
从Akt/PKB,PKCζ,p70 S6K和MAPK级联得到的负反馈信息会导致丝氨酸的磷酸化和IRS信号通路的失活。
(2)AMPK Signaling:AMP激活的蛋白激酶( AMPK, AMP-activated protein kinase)是细胞能量稳态平衡调节的关键分子。
这个激酶的激活响应细胞耗竭胞内ATP的压力,如低葡萄糖,缺氧,缺血和热激。
该激酶是一个异质三聚体,由一个α催化亚基和β、γ调节亚基组成。
AMP与γ亚基结合后,该复合体发生异构化并激活,成为上游主要激酶AMPKK,LKB1更适合的底物,在α亚基的Thr172位发生磷酸化。
当代谢激素包括adiponectin和leptin刺激时,AMPK响应细胞内钙的改变也能被CAMKK2在Thr172直接磷酸化。

PI3K/AKT旗号通路之阳早格格创做磷脂酰肌醇3激酶(PI3Ks)旗号介进删殖、瓦解、凋亡战葡萄糖转运等多种细胞功能的安排. 连年去创造, IA型PI3K战其下游分子蛋黑激酶B(PKB或者Akt)所组成的旗号通路取人类肿瘤的爆收死少稀切相闭. 该通路安排肿瘤细胞的删殖战存活, 其活性非常十分没有但是能引导细胞恶性转移, 而且取肿瘤细胞的迁移、黏附、肿瘤血管死成以及细胞中基量的落解等相闭, 暂时以PI3KAkt旗号通路闭键分子为靶面的肿瘤治疗战术正正在死少中.正在PI3K家属中, 钻研最广大的是能被细胞表面受体所激活的I型PI3K. 哺乳动物细胞中Ι型PI3K又分为IA战IB二个亚型, 他们分别从酪氨酸激酶连交受体战G蛋黑连交受体传播旗号.IA 型PI3K是由催化亚单位p110战安排亚单位p85所组成的二散体蛋黑, 具备类脂激酶战蛋黑激酶的单沉活性.PI3K通过二种办法激活, 一种是取具备磷酸化酪氨酸残基的死少果子受体或者连交蛋黑相互效率, 引起二散体构象改变而被激活; 另一种是通过Ras战p110曲交分离引导PI3K的活化. PI3K激活的截行是正在量膜上爆收第二疑使PIP3, PIP3取细胞内含有PH结构域的旗号蛋黑Akt战PDK1(phosphoinositidedependentkinase1)分离, 督促PDK1磷酸化Akt蛋黑的Ser308引导Akt的活化. Akt还能通过PDK2(如调整素连交激酶ILK)对于其Thr473的磷酸化而被激活.活化的Akt通过磷酸化效率激活或者压造其下游靶蛋黑Bad 、Caspase9、NFκB、GSK3、FKHR、 p21Cip1战p27 Kip1等, 从而安排细胞的删殖、瓦解、凋亡以及迁移等. PI3KAkt旗号通路的活性被类脂磷酸酶PTEN(phosphatase and tensin homolog deleted on chromosome ten)战SHIP(SH2containing inositol 5phosphatase)背安排, 他们分别从PIP3的3´战5´去除磷酸而将其转形成PI(4,5)P2战PI(3,4)P2而落解. 迄今为行, 尚已创造下调Akt活性的特同磷酸酶, 但是用磷酸酶压造剂处理细胞后, 创造Akt的磷酸化战活性均有所减少. 迩去创造Akt能被一种C终端安排蛋黑(CTMP)所得活, CTMP能分离Akt并通过压造Akt的磷酸化而阻断下游旗号的传播, CTMP的过表黑不妨顺转vAkt转移细胞的表型. 热戚克蛋黑90(HSP90)亦能分离Akt, 遏行Akt 被PP2A磷酸酶的去磷酸化而得活, 果此具备呵护Akt的效率.原旗号转导波及的旗号分子主要包罗Integrin,FAK,Paxillin,ILK,PIP3,S6,p70S6K,RTK,Gab1,Gab2,IRS1,PI3K,PTEN,AKT,PDK1,Cytokine Receptor,Jak1,CD19,BCR,Ag,BCAP,Syk,Lyn,GPCR,TSC1,TSC2,Gβγ,GαGTP,PP2A,PHLPP,CTMP,PDCD4,4EBP1,ATG13,mTORC1,TSC1,TSC2,PRAS40,XIAP,FoxO1,Bim,Bcl2,Bax,MDM2,p53,Bax,Bad,1433,Wee1,Myt1,p27Kip1,p21Waf1/Cip1,CyclinD1,GSK3,GS,Bcl2,mTORC2,LaminA,Tpl2,IKKα,eNOS,GABAAR,Huntingtin,Ataxin1,PFKFB2,PIP5K,AS160等.。
PI3K/AKT信号通路与心力衰竭摘要:丝氨酸/苏氨酸激酶(serine/threoninekinase,AKT)是真核细胞中参与细胞信号转导的关键分子目前已经证实PI3K(phosphatidylinositol-3-kinase,PI3K)/AKT信号通路在人类肿瘤、代谢紊乱、肾脏疾病以及精神障碍等疾病中发挥着重要的作用。
近年来的研究还发现P13K/AKT信号通路的激活会对心肌细胞的生长、代谢以及凋亡等活动产生影响,且该通路及其中的很多受体、激酶被证实与心力衰竭关系密切,这使该信号通路在心力衰竭的发病机制、诊断及治疗等方面的研究日益受到重视。
总结PI3K/AKT的结构特点、相关信号转导机制及其与心力衰竭的关系将有利于更好地理解心力衰竭的发病机制。
关键词:心力衰竭;磷脂酰肌醇3-激酶(PI3K);丝氨酸/苏氨酸激酶(AKT);信号通路ThePI3K/AKTSignalingPathwayandHeartFailureFANLiang-liang1,MALi-ning2,PENGYuan-liang1,XIANGRong1*(1.DepartmentofCellBiology,SchoolofLifeSciences,CentralSouthUniversity,Changsha410013,Hunan,China;2.HainanGeneralHospital,Haikou570311,Hainan,China)Abstract:Serine/threoninekinase(AKT)isakeymoleculewhichparticipatesinthecellularsignaltrans?ductionofeukaryoticcells.No withasbeenconfirmedthatthePI3K(phosphatidylinositol-3-kinase,PI3K)/AKTsignalingpathwayplaysanimportantroleinhumandiseases,suchasswelling,metabolicdisorders,kidneydiseasesandmentaldisorders.Researchinrecentyearshasalsodiscoveredthattheacti vationofPI3K/AKTsignalpathwaywouldhaveeffectonthegrowth,metabolismandapoptosisofmyocardialcell.Be?sides,thissignalingpathway,togetherwithmanyofitsreceptorsandkinaseshasbeenprovedtohavecloserelationwithhear tfailure.Becauseofallthesemechanisms,peoplearepayingmoreattentiontotheroleofthispathwayinresearchaboutthepathogenesis,diagnosisandtreatmentofheartfailure.Tosummarizethestructurecharacteristicsandtherel atedsignaltransductionpathwayofPI3K/AKTwillhelpusunderstandthemechanismandrel ationshipofPI3K/AKTpathwayandheartfailure.Keywords:heartfailure;PI3K(phosphatidylinositol-3-kinase);AKT (serine/threoninekinase);signalingpathway(LifeScienceResearch,2015,19(1):085?090)心力衰竭(heartfailure,HF)是指各种原因造成的心肌受损,使心脏收缩和(或)舒张功能出现障碍,心脏泵血功能下降,在足够的充盈压下不能射出相应血量来满足机体需要而引发的一组临床综合征。
PI3KAktmTOR信号通路和靶向药物PI3K/AKT/mTOR信号通路的梳理着实不易,寄希望该贴梳理的知识帮助到患者和家属。
很多癌种都会涉及这个信号通路,很多患者都面对这个信号通路的药物问题。
经常读报告看到mTOR抑制剂的靶向药物推荐,而且很多基因突变都与该种类型的靶向药物相关,我印象中的基因有PIK3CA,PTEN、NF1等等。
当然也不是这些基因随便某个位点出现了改变就可以用mTOR抑制剂,某些变异只是良性多态性,没有影响到蛋白的结构和功能,对于肿瘤的增殖没有影响,相对应的靶向药物推荐也就没有意义。
一、PI3K信号通路的突变类型我们不从信号通路的科学去阐释,如此繁多的术语和英文字符看的我自己都头疼,我们先用尽量简单的语言去看PI3K信号通路的突变类型有哪些,该信号通路的激活经常是通过关键节点的直接突变,如PIK3CA和AKT1的激活突变或扩增,以及PTEN这个负调控基因的失活突变。
另外PI3K信号通路的激活也可能源于RAS基因,下面的图1是PI3K信号通路的组成部分、和相互之间的抑制和激活关系。
图1:PI3K/AKT信号通路。
图1突变的机制包含受体酪氨酸激酶和原癌基因(如ERBB2,KRAS)的基因扩增/突变,PIK3CA、AKT、TSC1/2、mTOR的突变。
抑癌基因如PTEN、INPP4B和LKB1的失活突变。
mTOR激酶包含两个核蛋白TORC1和TORC2,分别位于AKT基因的上游和下游。
粉红色背景框的是原癌基因,而蓝色背景框的是抑癌基因。
需要注意的是,MEK和ERK蛋白是涉及到PI3K信号通路中的。
所以有时要把两种信号通路的靶向药物联合起来开展临床试验的原因。
下面的图2是具体的某些基因的突变形式,以及它们出现的肿瘤类型。
图2:PI3K信号通路相关的基因突变二、针对mTOR的靶向药物mTOR是一个丝氨酸——苏氨酸激酶,属于PI3K相关的激酶家族,参与介导生长、营养、能量获取等来调控细胞增殖、凋亡等。
经典信号通路之PI3K/AKT信号通路The PI3K/AKT and related pathways are important in inte RNA lizing the effects of external growth factors and of membrane tyrosine kinases. Activation of membrane kinases including epidermal growth factor receptor (EGFR) by external growth factors initiates receptor dimerization and subsequent events to activate these intracellular pathways. AKT is activated downstream of PI3K and has multiple targets. AKT and the cellular energy sensors LKB1 (STK11) and AMP-activated protein kinase (AMPK) exert opposing effects on mammalian target of rapamycin (mTOR), which is activated by AKT. ERK, extracellular signal regulated kinase; FKHR, forkhead; GDP, guanosine diphosphate; IRS, insulin receptor substrate; GSK3, glycogen synthase kinase 3; MAPK, mitogen-activated protein kinase; NF-B, nuclear factor-B; PIP2, phosphatidylinositol-3,4-diphosphate; PIP3, phosphatidylinositol-3,4,5-triphosphate; PKC, protein kinase C; STAT, signal transducer and activator of transcription.磷脂酰肌醇3-激酶(PI3Ks)信号参与增殖、分化、凋亡和葡萄糖转运等多种细胞功能的调节. 近年来发现, IA型PI3K和其下游分子蛋白激酶B(PKB或Akt)所组成的信号通路与人类肿瘤的发生发展密切相关. 该通路调节肿瘤细胞的增殖和存活, 其活性异常不仅能导致细胞恶性转化, 而且与肿瘤细胞的迁移、黏附、肿瘤血管生成以及细胞外基质的降解等相关, 目前以PI3K-Akt信号通路关键分子为靶点的肿瘤治疗策略正在发展中.在PI3K家族中, 研究最广泛的是能被细胞表面受体所激活的I型PI3K. 哺乳动物细胞中Ι型PI3K又分为IA和IB两个亚型, 他们分别从酪氨酸激酶连接受体和G蛋白连接受体传递信号.IA 型PI3K是由催化亚单位p110和调节亚单位p85所组成的二聚体蛋白, 具有类脂激酶和蛋白激酶的双重活性.PI3K通过两种方式激活, 一种是与具有磷酸化酪氨酸残基的生长因子受体或连接蛋白相互作用, 引起二聚体构象改变而被激活; 另一种是通过Ras和p110直接结合导致PI3K的活化. PI3K激活的结果是在质膜上产生第二信使PIP3, PIP3与细胞内含有PH结构域的信号蛋白Akt和PDK1(phosphoinositidedependentkinase-1)结合, 促使PDK1磷酸化Akt蛋白的Ser308导致Akt的活化. Akt还能通过PDK2(如整合素连接激酶ILK)对其Thr473的磷酸化而被激活.活化的Akt通过磷酸化作用激活或抑制其下游靶蛋白Bad 、Caspase9、NF-κB、GSK-3、FKHR、 p21Cip1和p27 Kip1等, 进而调节细胞的增殖、分化、凋亡以及迁移等.PI3K-Akt信号通路的活性被类脂磷酸酶PTEN(phosphatase and tensin homolog deleted on chromosome ten)和SHIP(SH2-containing inositol 5-phosphatase)负调节, 他们分别从PIP3的3´和5´去除磷酸而将其转变成PI(4,5)P2和PI(3,4)P2而降解. 迄今为止,尚未发现下调Akt活性的特异磷酸酶, 但用磷酸酶抑制剂处理细胞后, 发现Akt的磷酸化和活性均有所增加. 最近发现Akt能被一种C末端调节蛋白(CTMP)所失活, CTMP能结合Akt 并通过抑制Akt的磷酸化而阻断下游信号的传递, CTMP的过表达能够逆转v-Akt转化细胞的表型. 热休克蛋白90(HSP90)亦能结合Akt, 阻止Akt被PP2A磷酸酶的去磷酸化而失活, 因此具有保护Akt的作用.本信号转导涉及的信号分子主要包括Integrin,FAK,Paxillin,ILK,PIP3,S6,p70S6K,RTK,Gab1,Gab2,IRS-1,PI3K,PTEN,AKT,PDK1,Cytokine Receptor,Jak1,CD19,BCR,Ag,BCAP,Syk,Lyn,G PCR,TSC1,TSC2,Gβγ,GαGTP,PP2A,PHLPP,CTMP,PDCD4,4E-BP1,ATG13,mTORC1,TSC1,TSC2,PRAS40,XIAP,FoxO1,Bim,Bcl-2,Bax,MDM2,p53,Bax,Bad,14-3-3,Wee1,Myt1,p27Kip1,p21Waf1/Cip1,CyclinD1,GSK-3,GS,Bcl-2,mTORC2,LaminA,Tpl2,IKKα,eNOS,GABAAR,Huntingtin,Ataxin-1,PFKFB2,PIP5K,AS160等。
pikakt信号通路图谱P I3K/A K T信号通路磷脂酰肌醇3-激酶(PI3Ks)信号参与增殖、分化、凋亡和葡萄糖转运等多种细胞功能的调节. 近年来发现, IA型PI3K和其下游分子蛋白激酶B(PKB或Akt)所组成的信号通路与人类肿瘤的发生发展密切相关. 该通路调节肿瘤细胞的增殖和存活, 其活性异常不仅能导致细胞恶性转化, 而且与肿瘤细胞的迁移、黏附、肿瘤血管生成以及细胞外基质的降解等相关, 目前以PI3K-Akt信号通路关键分子为靶点的肿瘤治疗策略正在发展中.在PI3K家族中, 研究最广泛的是能被细胞表面受体所激活的I型PI3K. 哺乳动物细胞中Ι型PI3K又分为IA和IB两个亚型, 他们分别从酪氨酸激酶连接受体和G蛋白连接受体传递信号.IA 型PI3K是由催化亚单位p110和调节亚单位p85所组成的二聚体蛋白, 具有类脂激酶和蛋白激酶的双重活性.PI3K通过两种方式激活, 一种是与具有磷酸化酪氨酸残基的生长因子受体或连接蛋白相互作用, 引起二聚体构象改变而被激活; 另一种是通过Ras和p110直接结合导致PI3K的活化. PI3K激活的结果是在质膜上产生第二信使PIP3, PIP3与细胞内含有PH结构域的信号蛋白Akt和PDK1(phosphoinositidedependentkinase-1)结合, 促使PDK1磷酸化Akt蛋白的Ser308导致Akt的活化. Akt还能通过PDK2(如整合素连接激酶ILK)对其Thr473的磷酸化而被激活.活化的Akt通过磷酸化作用激活或抑制其下游靶蛋白Bad 、Caspase9、NF-κB、GSK-3、FKHR、p21Cip1和p27 Kip1等, 进而调节细胞的增殖、分化、凋亡以及迁移等.PI3K-Akt信号通路的活性被类脂磷酸酶PTEN(phosphatase and tensin homolog deleted on chromosome ten)和SHIP(SH2-containing inositol 5-phosphatase)负调节, 他们分别从PIP3的3′和5′去除磷酸而将其转变成PI(4,5)P2和PI(3,4)P2而降解. 迄今为止, 尚未发现下调Akt活性的特异磷酸酶, 但用磷酸酶抑制剂处理细胞后, 发现Akt的磷酸化和活性均有所增加. 最近发现Akt能被一种C末端调节蛋白(CTMP)所失活, CTMP能结合Akt并通过抑制Akt的磷酸化而阻断下游信号的传递, CTMP的过表达能够逆转v-Akt转化细胞的表型. 热休克蛋白90(HSP90)亦能结合Akt, 阻止Akt被PP2A磷酸酶的去磷酸化而失活, 因此具有保护Akt的作用.本信号转导涉及的信号分子主要包括Integrin,FAK,Paxillin,ILK,PIP3,S6,p70S6K,RTK,Gab1,Gab2,IRS-1,PI3K,PTEN,AKT,PDK1,Cytokine Receptor,Jak1,CD19,BCR,Ag,BCAP,Syk,Lyn,GPCR,TSC1,TSC2,Gβγ,GαGTP,PP2A,PHLPP,CTMP,PDCD4,4E-BP1,ATG13,mTORC1,TSC1,TSC2,PRAS40,XIAP,FoxO1,Bim,Bcl-2,Bax,MDM2,p53,Bax,Bad,14-3-3,Wee1,Myt1,p27Kip1,p21Waf1/Cip1,CyclinD1,GSK-3,GS,Bcl-2,mTORC2,LaminA,Tpl2,IKKα,eNOS,GABAAR,Huntingtin,Ataxin-1,PFKFB2,PIP5K,AS160等。
p i k a k t信号通路图谱 WTD standardization office【WTD 5AB- WTDK 08- WTD 2C】
P I3K/A K T信号通路
磷脂酰肌醇3-激酶(PI3Ks)信号参与增殖、分化、凋亡和葡萄糖转运等多种细胞功能的调节. 近年来发现, IA型PI3K和其下游分子蛋白激酶B(PKB或Akt)所组成的信号通路与人类肿瘤的发生发展密切相关. 该通路调节肿瘤细胞的增殖和存活, 其活性异常不仅能导致细胞恶性转化, 而且与肿瘤细胞的迁移、黏附、肿瘤血管生成以及细胞外基质的降解等相关, 目前以PI3K-Akt信号通路关键分子为靶点的肿瘤治疗策略正在发展中.
在PI3K家族中, 研究最广泛的是能被细胞表面受体所激活的I型PI3K. 哺乳动物细胞中Ι型PI3K又分为IA和IB两个亚型, 他们分别从酪氨酸激酶连接受体和G蛋白连接受体传递信号.IA 型PI3K是由催化亚单位p110和调节亚单位p85所组成的二聚体蛋白, 具有类脂激酶和蛋白激酶的双重活性.PI3K通过两种方式激活, 一种是与具有磷酸化酪氨酸残基的生长因子受体或连接蛋白相互作用, 引起二聚体构象改变而被激活; 另一种是通过Ras和p110直接结合导致PI3K的活化. PI3K激活的结果是在质膜上产生第二信使PIP3, PIP3与细胞内含有PH结构域的信号蛋白Akt和
PDK1(phosphoinositidedependentkinase-1)结合, 促使PDK1磷酸化Akt蛋白的
Ser308导致Akt的活化. Akt还能通过PDK2(如整合素连接激酶ILK)对其Thr473的磷酸化而被激活.活化的Akt通过磷酸化作用激活或抑制其下游靶蛋白Bad 、
Caspase9、NF-κB、GSK-3、FKHR、 p21Cip1和p27 Kip1等, 进而调节细胞的增殖、分化、凋亡以及迁移等.
PI3K-Akt信号通路的活性被类脂磷酸酶PTEN(phosphatase and tensin homolog deleted on chromosome ten)和SHIP(SH2-containing inositol 5-phosphatase)负调节, 他们分别从PIP3的3′和5′去除磷酸而将其转变成PI(4,5)P2和PI(3,4)P2而
降解. 迄今为止, 尚未发现下调Akt活性的特异磷酸酶, 但用磷酸酶抑制剂处理细胞后, 发现Akt的磷酸化和活性均有所增加. 最近发现Akt能被一种C末端调节蛋白(CTMP)所失活, CTMP能结合Akt并通过抑制Akt的磷酸化而阻断下游信号的传递, CTMP的过表达能够逆转v-Akt转化细胞的表型. 热休克蛋白90(HSP90)亦能结合Akt, 阻止Akt被PP2A磷酸酶的去磷酸化而失活, 因此具有保护Akt的作用.
本信号转导涉及的信号分子主要包括
Integrin,FAK,Paxillin,ILK,PIP3,S6,p70S6K,RTK,Gab1,Gab2,IRS-1,
PI3K,PTEN,AKT,PDK1,Cytokine Receptor,Jak1,CD19,BCR,Ag,BCAP,Syk,Lyn,GPCR,TSC1,TSC2,Gβγ,GαGTP,PP2A,PHLPP,CTMP,PDCD4,4E-BP1,ATG13,mTORC1,TSC1,TSC2,PRAS40,XIAP,FoxO1,Bim,Bcl-2,Bax,MDM2,
p53,Bax,Bad,14-3-3,Wee1,Myt1,p27Kip1,p21Waf1/Cip1,CyclinD1,GSK-3,GS,Bcl-2,mTORC2,LaminA,Tpl2,IKKα,eNOS,GABAAR,Huntingtin,Ataxin-1,PFKFB2,PIP5K,AS160等。