上海交大 中文翻译 生物化学课本:第17章-柠檬酸循环
- 格式:doc
- 大小:10.62 MB
- 文档页数:54

1.糖的有氧氧化.:指在机体氧供充足时,葡萄糖彻底氧化成CO2和H2O ,并释放出能量的过程。
是机体主要供能方式。
反应部位:胞浆及线粒体糖的有氧氧化过程第一阶段:酵解途径第二阶段:丙酮酸的氧化脱羧第三阶段:三羧酸循环第四阶段:氧化磷酸化2.糖的有氧氧化第一阶段:葡萄糖循糖酵解途径分解为丙酮酸此阶段与无氧分解的过程相似,不同的是3-磷酸甘油醛脱氢生成NADH+H+的去向不同。
无氧的情况下,NADH+H+在细胞浆中将丙酮酸还原生成乳酸;在有氧的情况下,NADH+H+经穿梭作用进入线粒体,氧化成水和能量(3 or 5 ATP)。
第二阶段:丙酮酸进入线粒体氧化脱羧生成乙酰CoA此阶段1分子Glu生成2分子NADH+H+ , 5分子ATP。
TPP丙酮酸————————→乙酰CoA丙酮酸脱氢酶系丙酮酸脱氢酶系包括三种酶和六种辅助因子:三种酶——E1-丙酮酸脱氢酶组分;E2-二氢硫辛酰转乙酰基酶;E3-二氢硫辛酸脱氢酶六种辅助因子——TPP、硫辛酸、CoA-SH、FAD、NAD+、Mg2+多酶复合体位于线粒体内;原核细胞在胞液中TPP的作用:脱羧酶辅酶,将底物移入(出)脱羧酶的活性中心。
此阶段反应特点:_ 反应速度快并且为不可逆反应。
_ 反应中生成的NADH+H+直接进入电子传递链进行氧化磷酸化生成水,产生2.5 ATP。
_ 生成的乙酰辅酶A进入三羧酸循环,CO2可由肺呼出或参与机体内代谢。
分步反应:① O E1CH3-C-COOH + TPP——→羟乙基TPP + CO2② E2羟乙基TPP + 硫辛酸——→乙酰硫辛酸+ TPP③ E2乙酰硫辛酸+ HS~CoA ——→乙酰-CoA + HS-L-HS④ E3HS-L-HS + FAD ——→硫辛酸 + FADH 2⑤ E3FADH2 + NAD+ ——→FAD + NADH+H+砷化物对硫辛酸的毒害作用,砷化物抑制丙酮酸脱氢酶复合体的机制也表现在对α-酮戊二酸脱氢酶复合体的抑制上。

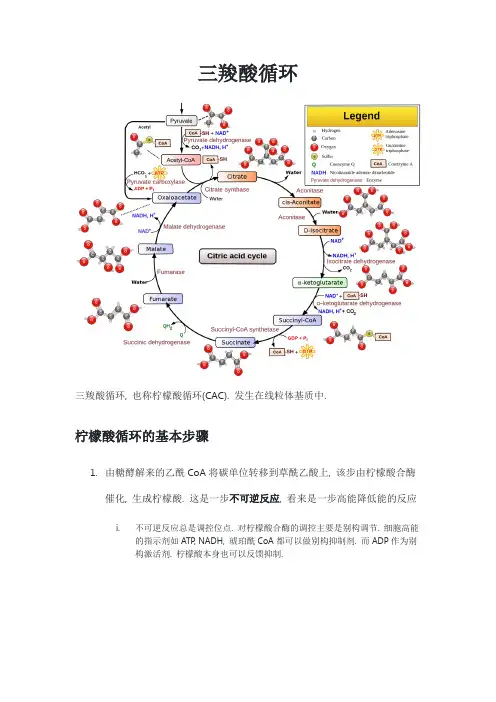
三羧酸循环三羧酸循环, 也称柠檬酸循环(CAC). 发生在线粒体基质中.柠檬酸循环的基本步骤1.由糖酵解来的乙酰CoA将碳单位转移到草酰乙酸上, 该步由柠檬酸合酶催化, 生成柠檬酸. 这是一步不可逆反应, 看来是一步高能降低能的反应i.不可逆反应总是调控位点. 对柠檬酸合酶的调控主要是别构调节. 细胞高能的指示剂如ATP, NADH, 琥珀酰CoA都可以做别构抑制剂. 而ADP作为别构激活剂. 柠檬酸本身也可以反馈抑制.2.柠檬酸异构为异柠檬酸, 由顺乌头酸酶催化, 该步可逆, 但由于下一步很快. 所以这一步也经常按正方向进行. 该步是必不可少的, 将底物变的更易氧化.(羟基从中间移至一侧)3.上述羟基被氧化, 异柠檬酸被氧化脱羧, 形成α-酮戊二酸, 由异柠檬酸脱氢酶催化. 该反应强烈放能, 也是不可逆反应.涉及氧化还原的反应往往有NAD+的参与. NAD+把异柠檬酸氧化.自己生成NADHi.植物对于它的调控有共价修饰. 动物中则多为别构调节. ATP是异柠檬酸脱氢酶的负别构效应物, 而ADP和钙离子是正别构效应物, 可能是因为钙离子代表了肌肉收缩的信号. NADH作为产物也可以竞争性反馈抑制.4.α-酮戊二酸氧化脱羧生成琥珀酰CoA, 由α-酮戊二酸脱氢酶系催化. 之后都不能再掉碳了. 同样涉及NAD+变为NADH, 这也是不可逆反应i.该酶系的调控是CAC的重要调控点,与丙酮酸脱氢酶系相似, 但少了共价修饰的调节. 它主要有别构调节和产物的竞争性反馈抑制. 钙离子和ADP可以别构激活, α酮戊二酸脱氢酶, 琥珀酰CoA和NADH分别作为产物竞争反馈抑制后两个酶.5.唯一一步底物水平磷酸化, 琥珀酰CoA推动GTP形成, 生成琥珀酸, 由琥珀酰CoA合酶催化. 该步可逆.6.琥珀酸脱氢生成延胡索酸, 由琥珀酸脱氢酶催化, 该酶在电子传递链中存在重要作用, 其就是复合体Ⅱ的主要成分(见电子传递链). 该步脱氢是用FAD做的.生成FADH27.延胡索酸生成苹果酸,由延胡索酸酶催化.8.苹果酸被氧化成草酰乙酸, 由苹果酸脱氢酶催化, 这个过程与苹果酸-天冬氨酸穿梭系统是一致的. NAD+变为NADH(见糖酵解)总的来说柠檬酸循环生成了3个线粒体的NADH, 1个FADH2, 1个GTP, 等同于10个ATP, 当然, 这只考虑了一个乙酰CoA, 而一个葡萄糖最后能形成两个乙酰CoA, 也就是等同于20个ATP.柠檬酸循环进来一个二碳单位, 同时途中掉过两次碳. 但这两个碳并不是进来的那个. CAC必须要有氧, 否则变化的NAD+, FAD无法再生.柠檬酸循环的回补反应CAC中的多种代谢产物可能被其他代谢通路用掉, 因此需要回补.1.草酰乙酸的回补.这是回补中最主要的途径, 由PEP被PEP羧化酶和生物素作用生成草酰乙酸, 或者由丙酮酸消耗1分子ATP被丙酮酸羧化酶和生物素反应, 生成草酰乙酸. 或者绕一圈, 由丙酮酸消耗NADPH, 变成苹果酸, 再生成一分子NADH 变成草酰乙酸. 这一步反应在后来的脂肪酸代谢也有重要的作用(见脂肪酸代谢)2.α-酮戊二酸的回补.谷丙转氨酶可以把谷氨酸转化成α-酮戊二酸.。


生物化学名词解释(英汉)第八章1,分解代谢反应(catabolic reaction):降解复杂分子为生物体提供小的构件分子和能量的代谢反应。
2,合成代谢反应(anablic reaction):合成用于细胞维持和生长所需分子的代谢反应。
3,反馈抑制(feedback inbition):催化一个代谢途径中前面反应的酶受到同一途径终产物抑制的现象4,前馈激活(feed-forward activition):代谢途径中一个酶被该途径中前面产生的代谢物激活的现象。
5,标准自由能变化(△GO):相应于在一系列标准条件(温度298K,压力1atm(=101.325KPa),所有溶质的浓度都是mol/L)下发生的反应自由能变化。
△GO′表示pH7.0条件下的标准自由能变化。
6,标准还原电动势(EO′):25℃和pH7.0条件下,还原剂和它的氧化形式在1mol/L浓度下表现出的电动势.第九章1,酵解(glycolysis):由10步酶促反应组成的糖分解代谢途径。
通过该途径,一分子葡萄糖转化为两分子丙酮酸,同时净生成两分子A TP和两分子NADH。
2,发酵(fermentation):营养分子(Eg葡萄糖)产能的厌氧降解。
在乙醇发酵中,丙酮酸转化为乙醇和CO2。
3,巴斯德效应(Pasteur effect):氧存在下,酵解速度放慢的现象。
4,底物水平磷酸化(substrate phosphorlation):ADP或某些其它的核苷-5′—二磷酸的磷酸化是通过来自一个非核苷酸底物的磷酰基的转移实现的。
这种磷酸化与电子的传递链无关。
5,柠檬酸循环(citric acid cycle):也称为三羧酸循环(TAC)、Krebs循环。
是用于乙酰CoA 中的乙酰基氧化成CO2的酶促反应的循环系统,该循环的第一步是由乙酰CoA经草酰乙酸缩合形成柠檬酸。
6,回补反应(anaplerotic reaction):酶催化的,补充柠檬酸循环中间代谢物供给的反应,例如由丙酮酸羧化酶生成草酰乙酸的反应。



第九讲柠檬酸循环柠檬酸循环是指在有氧的情况下,葡萄糖酵解产生的丙酮酸氧化脱羧形成乙酰CoA。
乙酰CoA经一系列氧化、脱羧,最终生成CO2和H2O并产生能量的过程.它是糖、脂类和氨基酸代谢的最后共同途径,简称TCA循环。
丙酮酸的氧化脱羧是放能反应,不可逆,在真核细胞的线粒体基质中进行。
它的催化酶——丙酮酸脱氢酶系是一个多酶复合体,包括三种酶和五种辅助因子,分别是:丙酮酸脱氢酶、二氢硫辛酰转乙酰基酶、二氢硫辛酸脱氢酶、硫胺素焦磷酸酯TPP、硫辛酸、HS—CoA、NAD+、FAD。
生成的乙酰CoA进入TCA循环,参与TCA循环的酶共有九种,该循环主要可分为八个步骤:(1)柠檬酸合成酶催化乙酰CoA与草酰乙酸结合,生成六碳的柠檬酸,放出CoA。
(2)顺乌头酸酶催化柠檬酸先失去一个H2O而成顺乌头酸,再结合一个H2O转化为异柠檬酸。
(3)异柠檬酸脱氢酶催化异柠檬酸发生脱氢、脱羧反应,生成5碳的a-酮戊二酸,放出一个CO2,生成一个NADH+H+。
(4)a-酮戊二酸脱氢酶系催化a-酮戊二酸发生脱氢、脱羧反应,并和CoA结合,生成含高能硫键的4碳琥珀酰CoA,放出一个CO2,生成一个NADH+H+。
(5)琥珀酰辅酶A合成酶催化碳琥珀酰CoA脱去CoA和高能硫键,放出的能通过GTP转入ATP(6)琥珀酸脱氢酶催化琥珀酸脱氢生成延胡索酸,生成1分子FADH2。
(7)延胡索酸酶催化延胡索酸和水化合而成苹果酸。
(8)苹果酸脱氢酶催化苹果酸氧化脱氢,生成草酸乙酸,生成1分子NADH+H+。
柠檬酸循环的总反应方程式为:乙酰CoA+3NAD++FAD+GDP+Pi+2H2O→2CO2+3NADH+3H++FADH2+GTP+HS—CoA 循环中有四次脱氢反应,生成三分子NADH和一分子FADH2。
有一次底物水平磷酸化,生成一分子GTP,每完成一次循环,氧化分解掉一分子乙酰基,可生成12分子ATP。
TCA循环生物学意义:它是机体获得能量的最有效方式;是联系糖类、脂类、蛋白质三大物质代谢的纽带;循环中所产生的多种中间产物是生物体内重要物质;发酵工业上利用微生物三羧酸循环生产各种代谢产物。


乙酰辅酶A 柠檬酸异柠檬酸α-酮戊二酸琥珀酰辅酶A琥珀酸延胡索酸苹果酸草酰乙酸2C:乙酰CoA-SH 4C:草酰乙酸6c:柠檬酸不可逆反应2C+4C=6C柠檬酸合成酶乙酰辅酶A草酰乙酸柠檬酸限速酶不可逆反应草酰乙酸乙酰–CoA 柠檬酰-CoA Asp OAAanalog acetyl-CoA酸柠檬酸顺乌头酸异柠檬酸顺乌头酸酶顺乌头酸酶Prochiral C 前手性* 顺乌头酸酶能识别这两个羧甲基基团有助于柠檬酸结合活性位点柠檬酸的结构柠檬酸的立体结构该键没有固定不容易被攻击该键被固定容易被攻击完全激活位点柠檬酸的异构问题草酰乙酸L-柠檬酸异柠檬酸只有这种产物产生能够形成L-α-酮戊二酸但自然界不存在6C-1C=5C异柠檬酸脱氢酶异柠檬酸α-酮戊二酸不可逆反应异柠檬酸脱氢酶草酰琥珀酸异柠檬酸α-酮戊二酸5C-1C=4Cα-酮戊二酸氧化脱羧形成琥珀酰 - CoAα-酮戊二酸脱氢酶复合体α-酮戊二酸琥珀酰-CoA不可逆反应琥珀酰-CoA 琥珀酸琥珀酰-CoA合成酶GDP + Pi GTP CoA-SH琥珀酰合成酶结构琥珀酰CoA合成酶琥珀酰CoA琥珀酸合酶琥珀酸磷酸化酶琥珀酸脱氢形成延胡索酸琥珀酸延胡索酸琥珀酸脱氢酶FAD FAD H2延胡索酸水合成苹果酸延胡索酸 苹果酸延胡索酸酶H2O酸L-苹果酸草酰乙酸苹果酸脱氢酶NAD+NAD H + H +2C柠檬酸循环carbon atoms from acetyl CoAonly 柠檬酸循环柠檬酸异柠檬酸α-酮戊二酸琥珀酰CoA 琥珀酸延胡索酸苹果酸草酰乙酸乙酰CoA葡萄糖6-磷酸果糖3-磷酸甘油醛1,3-二磷酸甘油酸磷酸烯醇丙酮酸丙酮酸异柠檬酸α-酮戊二酸琥珀酰-CoA琥珀酸苹果酸6`磷酸葡萄糖1,6二磷酸果糖3-磷酸甘油酸1,3-二磷酸甘油酸丙酮酸乙酰CoAα-酮戊二酸琥珀酰-CoA琥珀酸延胡索酸草酰乙酸中心异化途径酶系八步连续反应乙酰-CoA草酰乙酸柠檬酸2CO23 NADH 1 FADH2 1 ATP/GTP异化/同化两用丙酮酸羧化酶填补反应。

柠檬酸循环的名词解释介绍柠檬酸循环(Krebs cycle),也称为三羧酸循环(tricarboxylic acid cycle)或卡尔文循环(Calvin cycle),是生物体内进行细胞呼吸的关键代谢途径之一。
它在有氧条件下通过氧化葡萄糖产生能量,并生成二氧化碳、水和能量富集的还原辅酶。
循环过程柠檬酸循环是一系列复杂的生化反应,涉及多个底物和酶的参与。
以下是柠檬酸循环的主要步骤:1.乳酸脱氢酶反应–乳酸通过乳酸脱氢酶转化为丙酮酸,同时产生NADH。
2.丙酮酸变羧化反应–丙酮酸通过丙酮酸脱羧酶的作用,变羧化为柠檬酸,并释放出二氧化碳。
3.柠檬酸异构反应–柠檬酸经过柠檬酸异构酶的作用,转化为异柠檬酸。
4.异柠檬酸变羧化反应–异柠檬酸通过异柠檬酸脱羧酶的作用,变羧化为α-酮戊二酸,并释放出二氧化碳。
5.α-酮戊二酸脱氢反应–α-酮戊二酸通过α-酮戊二酸脱氢酶的作用,产生NADH和脱羧产物。
6.脱羧产物再生–脱羧产物在多次反应中生成辅酶A,再经过复杂的反应路径得到柠檬酸。
7.总反应方程式–以上反应综合在一起,得到柠檬酸循环的总反应方程式:乳酸 + NAD+ + CoA-SH + ADP + Pi → Acetyl-CoA + NADH + H+ + ATP +H2O + CO2。
循环中的产物柠檬酸循环在每一次循环过程中产生以下重要的产物:1.ATP:通过底物级磷酸化反应(substrate-level phosphorylation),柠檬酸循环每循环一次可以产生1个ATP。
2.NADH和FADH2:在柠檬酸循环中,通过NAD+和FAD接受氢原子的转移,产生NADH和FADH2,这些将在后续的细胞呼吸过程中发挥重要的作用。
3.CO2:柠檬酸循环中产生的二氧化碳是细胞释放掉的废物,它将在呼吸过程中通过肺部排出体外。
循环调控柠檬酸循环的调控对于维持正常的细胞呼吸过程至关重要。
以下是柠檬酸循环的调控机制:1.NADH和ATP浓度:高浓度的NADH和ATP会抑制柠檬酸循环的进行,这是因为细胞内能源和氧气供应充足,不需要继续产生更多的能量。
乙醛酸循环的名词解释乙醛酸循环(Krebs cycle),也被称为三羧酸循环或柠檬酸循环,是生物化学中一个重要的代谢途径,被广泛应用于生物体内能量的产生和有机物的合成。
乙醛酸循环是在细胞质中被陆同发现的,他发现在含有六个碳原子的柠檬酸氧化,最终生成了含有五个碳原子的柠檬酸。
这个循环因而被称为三羧酸循环,因为它将一分子柠檬酸转化为另一分子柠檬酸的三步反应。
乙醛酸循环在线粒体的内膜上进行,这是细胞中的一个重要有机物代谢路线,提供了细胞内的氧化能量。
乙醛酸循环是一系列复杂的轮回反应,通过将脂肪、蛋白质和糖类分子分解为能量和机体所需的可用分子。
它启动于链脂酸的氧化。
脂肪酸在线粒体内经过β氧化反应被转化为乙醛酸,然后进入乙醛酸循环。
在乙醛酸循环中,乙酰辅酶A与草酰乙酸结合生成柠檬酸。
接下来的反应包括甲积酸的脱羧反应以及还原脱羧酶的作用。
乙醛酸循环中的还原脱羧酶将柠檬酸通过一系列的反应转化为α-酮戊二酸,同时产生能量(ATP、NADH和FADH2)。
最后,α-酮戊二酸再被转化为草酰乙酸,重新进入乙醛酸循环。
乙醛酸循环对生物体内能量的产生至关重要。
氧化磷酸化过程中产生的能量,主要来自于乙醛酸循环。
其中的还原酶可以将NAD+和FAD还原为NADH和FADH2,最终通过呼吸链的过程将产生的能量转化为三磷酸腺苷(ATP)。
这种ATP的产生方式被称为氧化磷酸化,是生物体内绝大部分能量来源。
此外,乙醛酸循环还参与合成一些有机物。
作为中间产物,乙醛酸循环可以提供合成葡萄糖、脂肪酸和氨基酸所需的苹果酸和柠檬酸。
克尔氏循环通过脱羧,合成了α-酮戊二酸。
α-酮戊二酸可以产生丙酮酸、甲酮酸和叶酸,这些有机物在生物体内具有重要的功能。
乙醛酸循环的运行是一个高度调控的过程。
它受到细胞内能量需求的影响,以及各种代谢产物的抑制和激活。
细胞内的ATP水平较高时,乙醛酸循环速率会降低,以避免产生过多的乙醛酸。
另外,ATP的产量还会影响到乙醛酸循环中的脱羧酶和还原脱羧酶的活性,进一步调控循环过程。
第17章柠檬酸循环
环形交叉路,或者转盘,是有助于交通流动的中心。
柠檬酸循环是细胞氧化碳原子的生化反应流中心。
碳燃料以乙酰辅酶A的形式加入生化反应流中心,柠檬酸循环也是生物合成的前体。
[(以上)Chris Warren/International Stock.]
概要
17.1 丙酮酸脱氢酶联结糖酵解途径和柠檬酸循环
17.2 柠檬酸循环氧化2-C单位
17.3 柠檬酸循环及其调控
17.4 柠檬酸循环提供生物合成的原料
17.5 乙醛酸循环使植物和细菌也能利用乙酸盐生长
葡萄糖经过厌氧的糖酵解途径代谢生成丙酮酸,只能得到一小部分的ATP。
葡萄糖的
1
大部分的ATP是耗氧代谢途径产生的。
耗氧代谢途径能够将糖酵解产物完全氧化成二氧化碳。
这一氧化作用由一系列反应(即柠檬酸循环,或称为三羧酸(TCA)循环、或Krebs 循环)完成。
柠檬酸循环是能源分子——碳水化合物、脂肪酸、和氨基酸氧化过程最后的共同途径。
大部分能源分子被加工成乙酰辅酶A的形式,进入三羧酸循环。
在有氧条件下,葡萄糖经糖酵解产生的丙酮酸被氧化脱羧生成乙酰辅酶A。
柠檬酸循环在真核生物中的线粒体里进行(图17.1),而糖酵解反应在细胞质中进行。
乙酰辅酶A
2
3
图17.1 线粒体。
电子显微镜图片清晰地显示线粒体的双膜结构。
线粒体内膜有很多内陷,这些内陷称为嵴。
丙酮酸的氧化脱羧和柠檬酸循环在线粒体基质进行。
[(左)Omikron/图片研究者]
柠檬酸循环捕获高能电子
柠檬酸循环是细胞的代谢中心。
任何分子被转化成乙酰辅酶A 或三羧酸循环的其他组分就可以进行有氧代谢。
这个循环也为许多生物分子(如氨基酸、核苷酸碱基、卜啉)的合成原料。
草酰乙酸(柠檬酸循环的组分之一)也是葡萄糖合成的原料。
在燃料分子转化成ATP 过程中三羧酸循环起什么作用?我们知道燃料分子是可以被氧化(即失去电子)的碳化物(第15章)。
柠檬酸循环含有一系列氧化-还原反应,导致乙酰基被氧化成两个CO 2。
此氧化产生的高能电子用来驱动ATP 的合成。
柠檬酸循环的功能是捕获碳燃料的高能电子。
图17.2显示柠檬酸循环的总况。
草酰乙酸(四碳化合物)与乙酰基缩合形成六碳原子的三羧酸。
这个六碳化合物连续丢失两分子CO 2,与此同时两次释放高能电子,形成四碳化合物。
这个四碳化合物被进一步加工,又生成草酰乙酸,启动新一轮三羧酸循环。
乙酰基(二碳单位)进入柠檬酸循环后被转化成两分子CO 2。
图17.2 柠檬酸循环总图。
柠檬酸循环氧化二碳单位,形成两分子CO2,1分子GTP,和高能电子(高能电子以NADH和FADH2的形式存在)。
注意柠檬酸循环本身既不产生大量ATP,也不利用氧气作为反应物质(图17.3)。
柠檬酸循环只是从乙酰辅酶A移去电子,并用这种电子形成NADH和FADH2。
三个氢负离子(即六个电子)转移给3分子NAD+,形成3个NADH;还有一对氢原子(因此是两个电子)转移给核黄素腺苷二磷酸(即FAD)形成一分子FADH2。
这些电子载体经氧化磷酸化(第18章)将电子传递给氧气,形成9分子ATP。
NADH和FADH2携带的电子经过一系列的膜蛋白传递,本身被氧化并产生跨膜质子梯度。
质子流过ATP合成酶将ADP和磷酸转化成ATP。
4
图17.3 细胞呼吸。
柠檬酸循环构成细胞呼吸的第一阶段,将碳燃料的电子转移给电子载体形成NADH和FADH2(左边)。
这些高能电子转移给氧气分子产生跨膜质子梯度(红色)。
随后利用质子梯度合成ATP(绿色)。
氧气还原和ATP合成构成氧化磷酸化。
柠檬酸循环与氧化磷酸化过程一起给有氧细胞提供了大量的能量——在人体内,超过90%的能量以这种方式制造。
有限数量的柠檬酸循环分子氧化能产生大量的NADH和FADH2,因此柠檬酸循环和氧化磷酸化产能的效率非常高效。
图17.2中指出,柠檬酸循环由一个四碳分子——草酰乙酸开始,依次反应至循环结束时重新生成一分子的草酰乙酸。
因此,一分子草酰乙酸可以参与很多乙酰基分子的氧化过程。
17.1丙酮酸脱氢酶联结糖酵解途径和柠檬酸循环
碳水化合物,多数是葡萄糖,经过糖酵解过程变成丙酮酸(第16章)。
在厌氧条件下,丙酮酸转化成乳酸或者乙醇(实际情况取决于生物体自身类型)。
在有氧条件下,丙酮酸由线粒体膜上特定的载体蛋白质运输到线粒体内。
在线粒体基质,丙酮酸在丙酮酸脱氢酶的作用下氧化脱羧生成乙酰辅酶A。
丙酮酸+ 辅酶A + NAD+ →乙酰辅酶A + CO2 + NADH + H +
这个不可逆反应是糖酵解和柠檬酸循环之间的连接纽带(图17.4)。
注意:此反应丙酮酸脱氢酶复合物催化产生二氧化碳、并将俘获的高能电子以NADH形式储存。
因此,因此丙酮酸脱羧酶促反应有柠檬酸循环反应所具有的很多关键特征。
图17.4 糖酵解和柠檬酸循环间的联系。
糖酵解产生的丙酮酸转化成乙酰辅酶A,后者是柠檬酸循环的燃料。
丙酮酸脱羧酶复合物是由3种活性不同的酶构成的、高度整合的、巨型复合物(表17.1)。
丙酮酸脱氢酶复合物属于同源复合物家族的成员。
这个家族包括柠檬酸循环酶酮戊二酸脱氢酶复合物。
这些复合物巨大,分子质量从400万到1000万道尔顿(图17.5),其复杂的结构能够将待转移基团(活性基团用绳状结构与酶复合物结构的核心连接)从一个活性位点转移到另一活性位点。
6
表17.1 大肠杆菌的丙酮酸脱氢酶复合物
酶简称链的数目辅基催化的反应丙酮酸脱氢酶组分E124 TPP 丙酮酸的氧化脱羧
二氢硫辛酸转乙酰酶E224 硫锌酰胺转移乙酰基团给辅酶A
二氢硫辛酰胺脱氢酶E312 FAD 重新形成氧化型硫锌酰胺
图17.5 大肠杆菌丙酮酸脱氢酶复合物的电子显微图谱。
7
机理:从丙酮酸合成乙酰辅酶A需要三种酶和五种辅酶
丙酮酸脱氢酶促反应机理很复杂(比丙酮酸脱氢酶的化学反应式复杂多了)。
这个反应需要丙酮酸脱氢酶复合体的三种酶活性和五种辅酶的参与。
辅酶硫胺焦磷酸(thiamine pyrophosphate,TPP)、硫辛酸(lipoic acid)、和FAD是酶催化的辅助因子,CoA和NAD+是定量反应的协助因子。
硫胺焦磷酸(TPP)硫辛酸
丙酮酸转化成乙酰辅酶A有三步:脱羧反应、氧化反应、和乙酰基团转移给辅酶A的化学反应。
这些反应步骤必需偶联才能保存脱羧反应所释放的自由能,驱动后续的NADH和乙酰
8。