真核生物转录特点与加工
- 格式:ppt
- 大小:785.00 KB
- 文档页数:23

原核生物和真核生物基因表达调控、复制、转录、翻译特点的比较1.相同点:转录起始是基因表达调控的关键环节①结构基因均有调控序列;②表达过程都具有复杂性,表现为多环节;③表达的时空性,表现为不同发育阶段和不同组织器官上的表达的复杂性;2.不同点:①原核基因的表达调控主要包括转录和翻译水平。
真核基因的表达调控主要包括染色质活化、转录、转录后加工、翻译、翻译后加工多个层次。
②原核基因表达调控主要为负调控,真核主要为正调控。
③原核转录不需要转录因子,RNA聚合酶直接结合启动子,由sita因子决定基因表的的特异性,真核基因转录起始需要基础特异两类转录因子,依赖DNA-蛋白质、蛋白质-蛋白质相互作用调控转录激活。
④原核基因表达调控主要采用操纵子模型,转录出多顺反子RNA,实现协调调节;真核基因转录产物为单顺反子RNA,功能相关蛋白的协调表达机制更为复杂。
⑤真核生物基因表达调控的环节主要在转录水平,其次是翻译水平。
原核生物基因以操纵子的形式存在。
转录水平调控涉及到启动子、sita因子与RNA聚合酶结合、阻遏蛋白、负调控、正调控蛋白、倒位蛋白、RNA聚合酶抑制物、衰减子等。
翻译水平的调控涉及SD序列、mRNA的稳定性不稳定(5’端和3’端的发夹结构可保护不被酶水解mRNA的5’端与核糖体结合可明显提高稳定性)、翻译产物及小分子RNA的调控作用。
真核生物基因表达的调控环节较多:在DNA水平上可以通过染色体丢失、基因扩增、基因重排、DNA甲基化、染色体结构改变影响基因表达。
在转录水平主要通过反式作用因子调控转录因子与TATA盒的结合、RNA聚合酶与转录因子-DNA复合物的结合及转录起始复合物的形成。
在转录后水平主要通过RNA修饰、剪接及mRNA运输的控制来影响基因表达。
在翻译水平有影响起始翻译的阻遏蛋白、5’AUG、5’端非编码区长度、mRNA的稳定性调节及小分子RNA。
真核基因调控中最重要的环节是基因转录,真核生物基因表达需要转录因子、启动子、沉默子和增强子。
1.真核生物基因为什么要进行RNA转录后加工?(P209)原核生物没有细胞器的分化,转录与翻译同时进行。
真核生物有细胞器的分化,基因表达在时间和空间上存在明显间隔。
转录在细胞核内进行,翻译在细胞质内完成。
真核生物基因的初始转录产物被非编码序列或间隔区段分开,转录产物不连续,需要转录后加工。
2.细胞内RNA原初转录物一般都需要经过哪些过程的加工修饰?(P209)真核生物细胞内转录的RNA原初转录物要经过一系列变化,包括:①5’端形成帽子结构;②3’端形成一段PolyA;③切去内含子;④反式剪接;⑤部分核苷酸修饰;⑥RNA 编辑;⑦RNA的再编辑;⑧RNA链的断裂等过程。
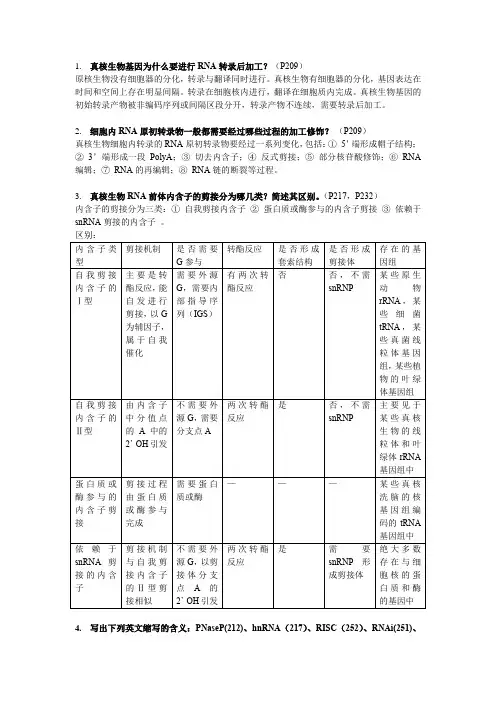
3.真核生物RNA前体内含子的剪接分为哪几类?简述其区别。
(P217,P232)内含子的剪接分为三类:①自我剪接内含子②蛋白质或酶参与的内含子剪接③依赖于snRNA剪接的内含子。
区别:4.写出下列英文缩写的含义:PNaseP(212)、hnRNA(217)、RISC(252)、RNAi(251)、剪接体(220)、自我剪接(228)、反义RNA(251或上课PPT)、RNA干涉(251)、siRNA(252)、选择性剪接(235)、核酶(229)PNaseP:催化切除5’端额外核苷酸的酶hnRNA:核内不均一RNARISC:沉默复合物RNAi:RNA干涉剪接体:是mRNA前提在剪接过程中组装形成的多组分复合物,由多种snRNA和蛋白质因子组成,即剪接体是具有催化剪接过程的核塘核蛋白复合体。
自我剪接:rRNA的内含子能够自我剪接,无需剪接体反义RNA:与mRNA互补的RNA分子,也包括与其它RNA互补的RNA分子RNA干涉:在双链RNA引导的抑制过程中存在某种扩增效应,且有某种没活性参与其中。
siRNA:短干涉RNA,发生转录后基因沉默的小的双链RNA选择性剪接:一个基因的初始转录产物在不同的分化细胞、不同的发育阶段乃至不同的生理状态下,可以有不同的剪接方式,得到不同的成熟mRNA和蛋白质产物核酶:RNA本身具有酶的活性称为核酶5.名词解释:套索结构(219)、转酯反应(227)、Dicer酶(253)、顺式剪接(239)、反式剪接(239)套索结构:RNA剪接过程中的中间结构,其中有形成的带尾巴的环形结构转酯反应:在剪接体上完成剪接反应的生化本质是磷酸二酯键的转移,又称转酯反应Dicer酶:能将双链RNA特异性切成大小均一的片段的酶称为Dicer酶顺式剪接:存在与同一基因中的两个或多个外显子和内含子的剪接,称为顺式剪接反式剪接:几个外显子不在同一基因甚至不在同意染色体上的剪接叫反式剪接6.什么是RNA的自我剪接?自我剪接有哪些类型?(217或232)RNA的自我剪接:能自发进行剪接,无需酶或蛋白质参与。


细菌和真核生物转录机制的差异
1不同的转录机制
细菌和真核生物的转录机制存在一定的差异。
这些差异主要体现在以下几个方面:
1.1启动子
首先,启动子是转录反应开始的地方,其作用是连接RNA聚合酶与DNA模板上。
细菌只有一种RNA聚合酶,它能识别相对简单的DNA 启动子。
而真核生物有三类不同的RNA聚合酶,它们能够识别不同的DNA启动子。
1.2转录调节
其次,细菌的转录调节主要依赖启动子及其上游的启动子元件,而真核生物的调节除了启动子以外,还受到基因间まゝ数距离,和内部及外部环境因素的影响。
1.3转录加工
最后,细菌转录后不会经过加工就会被进行翻译,而真核生物经过mRNA加工后再被翻译,包括5'修饰(5'cap)、流式处理(polyA tail)、剪切(splicing)、转录本结构调节(RNA stability)等。
总之,细菌和真核生物的转录机制的主要差异体现在启动子的多样性、转录调节的复杂性及转录加工的必要性上。


真核生物RNA的转录与原核生物RNA的转录过程在总体上基本相同,但是,其过程要复杂得多,主要有以下几点不同(图3-27)。
⒈真核生物RNA的转录是在细胞核内进行的,而蛋白质的合成则是在细胞质内进行的。
所以,RNA转录后首先必须从核内运输到细胞质内,才能指导蛋白质的合成。
⒉真核生物一个mRNA分子一般只含有一个基因,原核生物的一个mRNA分子通常含有多个基因,而除少数较低等真核生物外,一个mRNA分子一般只含有一个基因,编码一条多态链。
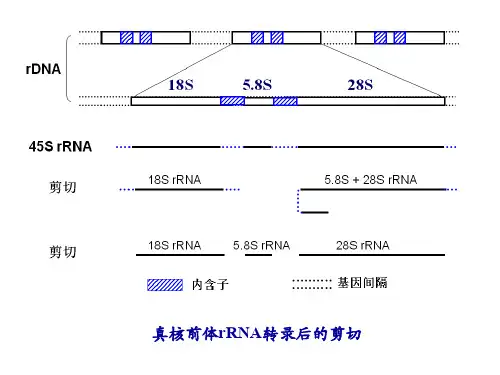
⒊真核生物RNA聚合酶较多在原核生物中只有一种RNA聚合酶,催化所有RNA的合成,而在真核生物中则有RNA聚合酶Ⅰ、RNA聚合酶Ⅱ和RNA聚合酶Ⅲ三种不同酶,分别催化不同种类型RNA的合成。
三种RNA聚合酶都是由10个以上亚基组成的复合酶。
RNA聚合酶Ⅰ存在于细胞核内,催化合成除5SrRNA 以外的所有rRNA的合成;RNA聚合酶Ⅱ催化合成mRNA前体,即不均一核RNA(hnRNA)的合成;RNA 聚合酶Ⅲ催化tRNA和小核RNA的合成。
⒋真核生物RNA聚合酶不能独立转录RNA 。
原核生物中RNA聚合酶可以直接起始转录合成RNA ,真核生物则不能。
在真核生物中,三种RNA聚合酶都必须在蛋白质转录因子的协助下才能进行RNA的转录。
另外,RNA聚合酶对转录启动子的识别,也比原核生物更加复杂,如对RNA聚合酶Ⅱ来说,至少有三个DNA的保守序列与其转录的起始有关,第一个称为TATA框(TATA box),具有共有序列TATAAAA,其位置在转录起始点的上游约为25个核苷酸处,它的作用可能与原核生物中的-10共有序列相似,与转录起始位置的确定有关。
第二个共有序列称为CCAAT框(CCAAT box),具有共有序列GGAACCTCT,位于转录起始位置上游约为50-500个核苷酸处。
如果该序列缺失会极大地降低生物的活体转录水平。
第三个区域一般称为增强子(enhancer),其位置可以在转录起始位置的上游,也可以在下游或者在基因之内。

原核生物和真核生物基因表达调控、复制、转录、翻译特点的比较1.相同点:转录起始是基因表达调控的关键环节①结构基因均有调控序列;②表达过程都具有复杂性,表现为多环节;③表达的时空性,表现为不同发育阶段和不同组织器官上的表达的复杂性;2.不同点:①原核基因的表达调控主要包括转录和翻译水平。
真核基因的表达调控主要包括染色质活化、转录、转录后加工、翻译、翻译后加工多个层次。
②原核基因表达调控主要为负调控,真核主要为正调控。
③原核转录不需要转录因子,RNA聚合酶直接结合启动子,由sita因子决定基因表的的特异性,真核基因转录起始需要基础特异两类转录因子,依赖DNA-蛋白质、蛋白质-蛋白质相互作用调控转录激活。
④原核基因表达调控主要采用操纵子模型,转录出多顺反子RNA,实现协调调节;真核基因转录产物为单顺反子RNA,功能相关蛋白的协调表达机制更为复杂。
⑤真核生物基因表达调控的环节主要在转录水平,其次是翻译水平。
原核生物基因以操纵子的形式存在。
转录水平调控涉及到启动子、sita因子与RNA聚合酶结合、阻遏蛋白、负调控、正调控蛋白、倒位蛋白、RNA聚合酶抑制物、衰减子等。
翻译水平的调控涉及SD序列、mRNA的稳定性不稳定(5’端和3’端的发夹结构可保护不被酶水解mRNA的5’端与核糖体结合可明显提高稳定性)、翻译产物及小分子RNA的调控作用。
真核生物基因表达的调控环节较多:在DNA水平上可以通过染色体丢失、基因扩增、基因重排、DNA甲基化、染色体结构改变影响基因表达。
在转录水平主要通过反式作用因子调控转录因子与TA TA盒的结合、RNA聚合酶与转录因子-DNA复合物的结合及转录起始复合物的形成。
在转录后水平主要通过RNA修饰、剪接及mRNA运输的控制来影响基因表达。
在翻译水平有影响起始翻译的阻遏蛋白、5’AUG、5’端非编码区长度、mRNA的稳定性调节及小分子RNA。
真核基因调控中最重要的环节是基因转录,真核生物基因表达需要转录因子、启动子、沉默子和增强子。


真核生物转录后加工方法1 真核生物转录后加工方法真核生物转录后加工(Transcription Post-Processing)是指在真核生物转录前实施多种变异来改变或增强其转录本表达。
它可以改变转录本结构或增强蛋白质表达效率。
真核生物转录后加工方法是可以精确协调转录因子与转录本的相互作用,从而达到增强特定基因的表达的目的。
转录后加工是研究基因表达和调控过程的主要工具,也用于开发转基因技术来改变宿主品系,或分离提高表达特定编码蛋白质功能的基因。
1.1 真核生物转录后加工技术真核生物转录后加工技术包括各种类型的转录本变异,如遗传变异(突变、转录因子缺失、RNA聚合酶选择性识别)、外源RNA干扰、DNA整合、去氧核糖核酸(RNAi)、CRISPR/Cas流程等。
1.1.1 遗传变异遗传变异是真核生物转录后加工的一种常见的技术,它涉及到各种形式的变异突变,如突变、转录因子缺失、RNA聚合酶选择性识别等。
这些技术可用于改变转录本结构,如增加剪接位点来增强表达量,减少反式转录等。
1.1.2 外源RNA干扰外源RNA干扰技术用特定的外源RNA来干扰细胞内特定的mRNA,从而减少特定基因的表达。
它也可以被用来调节特定基因的表达,或作为一种突变诊断的技术。
1.1.3 DNA整合DNA整合是让因子和特定的DNA序列整合到宿主RNA聚合酶II靶位点以调控特定基因的表达。
目前,常见的DNA整合方式包括荧光融合和杂交方法。
1.1.4 RNAiRNAi是通过使用特定的21~23碱基接头的双链RNA裂解靶标mRNA来减少特定基因的表达,从而抑制特定基因的功能。
1.1.5 CRISPR/Cas技术CRISPR/Cas技术是通过使用特定的RNA-目标-核酸(sgRNA)和多聚蛋白或酶来精确结合特定的DNA片段以调节特定基因的表达。
1.2 真核生物转录后加工作用真核生物转录后加工技术广泛应用于各种科学研究领域,其作用主要有:(1)用于研究转录调控机制(2)用于纯化特定编码蛋白质的基因(3)用于改变宿主基因组,进而改变宿主品系(4)用于生产新型转基因植物,提取编码蛋白的特定基因(5)用于开发医学应用1.3 真核生物转录后加工技术的局限性尽管真核生物转录后加工技术在研究转录调控和改变宿主品系、开发转基因技术方面发挥了重要作用,但它也具有一定的局限性。

原核生物和真核生物基因表达调控、复制、转录、翻译特点的比较1.相同点:转录起始是基因表达调控的关键环节①结构基因均有调控序列;②表达过程都具有复杂性,表现为多环节;③表达的时空性,表现为不同发育阶段和不同组织器官上的表达的复杂性;2.不同点:①原核基因的表达调控主要包括转录和翻译水平。
真核基因的表达调控主要包括染色质活化、转录、转录后加工、翻译、翻译后加工多个层次。
②原核基因表达调控主要为负调控,真核主要为正调控。
③原核转录不需要转录因子,RNA聚合酶直接结合启动子,由sita因子决定基因表的的特异性,真核基因转录起始需要基础特异两类转录因子,依赖DNA-蛋白质、蛋白质-蛋白质相互作用调控转录激活。
④原核基因表达调控主要采用操纵子模型,转录出多顺反子RNA,实现协调调节;真核基因转录产物为单顺反子RNA,功能相关蛋白的协调表达机制更为复杂。
⑤真核生物基因表达调控的环节主要在转录水平,其次是翻译水平。
原核生物基因以操纵子的形式存在。
转录水平调控涉及到启动子、sita因子与RNA聚合酶结合、阻遏蛋白、负调控、正调控蛋白、倒位蛋白、RNA聚合酶抑制物、衰减子等。
翻译水平的调控涉及SD序列、mRNA的稳定性不稳定(5’端和3’端的发夹结构可保护不被酶水解mRNA的5’端与核糖体结合可明显提高稳定性)、翻译产物及小分子RNA的调控作用。
真核生物基因表达的调控环节较多:在DNA水平上可以通过染色体丢失、基因扩增、基因重排、DNA甲基化、染色体结构改变影响基因表达。
在转录水平主要通过反式作用因子调控转录因子与TATA盒的结合、RNA聚合酶与转录因子-DNA复合物的结合及转录起始复合物的形成。
在转录后水平主要通过RNA修饰、剪接及mRNA运输的控制来影响基因表达。
在翻译水平有影响起始翻译的阻遏蛋白、5’AUG、5’端非编码区长度、mRNA的稳定性调节及小分子RNA。
真核基因调控中最重要的环节是基因转录,真核生物基因表达需要转录因子、启动子、沉默子和增强子。
1.相同点:转录起始是基因表达调控的关键环节①结构基因均有调控序列;②表达过程都具有复杂性,表现为多环节;③表达的时空性,表现为不同发育阶段和不同组织器官上的表达的复杂性;2.不同点:①原核基因的表达调控主要包括转录和翻译水平。
真核基因的表达调控主要包括染色质活化、转录、转录后加工、翻译、翻译后加工多个层次。
②原核基因表达调控主要为负调控,真核主要为正调控。
③原核转录不需要转录因子,RNA聚合酶直接结合启动子,由sita因子决定基因表的的特异性,真核基因转录起始需要基础特异两类转录因子,依赖DNA-蛋白质、蛋白质-蛋白质相互作用调控转录激活。
④原核基因表达调控主要采用操纵子模型,转录出多顺反子RNA,实现协调调节;真核基因转录产物为单顺反子RNA,功能相关蛋白的协调表达机制更为复杂。
⑤真核生物基因表达调控的环节主要在转录水平,其次是翻译水平。
原核生物基因以操纵子的形式存在。
转录水平调控涉及到启动子、sita因子与RNA聚合酶结合、阻遏蛋白、负调控、正调控蛋白、倒位蛋白、RNA聚合酶抑制物、衰减子等。
翻译水平的调控涉及SD序列、mRNA的稳定性不稳定(5’端和3’端的发夹结构可保护不被酶水解mRNA的5’端与核糖体结合可明显提高稳定性)、翻译产物及小分子RNA的调控作用。
真核生物基因表达的调控环节较多:在DNA水平上可以通过染色体丢失、基因扩增、基因重排、DNA甲基化、染色体结构改变影响基因表达。
在转录水平主要通过反式作用因子调控转录因子与TATA盒的结合、RNA聚合酶与转录因子-DNA复合物的结合及转录起始复合物的形成。
在转录后水平主要通过RNA修饰、剪接及mRNA运输的控制来影响基因表达。
在翻译水平有影响起始翻译的阻遏蛋白、5’AUG、5’端非编码区长度、mRNA的稳定性调节及小分子RNA。
真核基因调控中最重要的环节是基因转录,真核生物基因表达需要转录因子、启动子、沉默子和增强子。
真核生物和原核生物复制的不同点:①真核生物DNA的合成只是在细胞周期的S期进行,而原核生物则在整个细胞生长过程中都可进行DNA合成②原核生物DNA的复制是单起点的,而真核生物染色体的复制则为多起点的。
真核生物dna 转录的特点真核生物DNA转录是指在真核生物细胞中,DNA作为模板合成RNA的过程。
在这个过程中,DNA的信息被转录为RNA分子,这些RNA分子可以进一步被翻译为蛋白质。
真核生物DNA转录具有以下几个特点:1. 包含多个转录因子:真核生物DNA转录需要多个转录因子的参与。
其中最重要的是RNA聚合酶,它是一个复合酶,由多个亚单位组成。
RNA聚合酶能够识别DNA上的启动子序列,并在该位置开始合成RNA链。
2. 转录起始位点的多样性:在真核生物DNA上,一个基因通常含有多个外显子和内含子。
DNA转录时,RNA聚合酶结合到基因的启动子上,并在转录起始位点开始合成RNA链。
但是,不同基因的转录起始位点位置可能不同,甚至一个基因内的不同转录变体也可能有不同的转录起始位点。
3. 转录后修饰:真核生物DNA转录产生的RNA分子不同于细菌中的mRNA,需要经过转录后修饰才能成为成熟的mRNA。
这些转录后修饰包括剪接、5'帽子的添加、3'端的聚腺苷酸尾巴的添加等。
这些修饰能够增加RNA的稳定性、调控其翻译和定位等。
4. 转录调控:真核生物DNA转录的过程受到多种调控因素的影响。
其中一个重要的调控因素是转录因子。
转录因子是能够结合到DNA上的特定序列上,并调控基因转录的蛋白质。
转录因子可以促进或抑制RNA聚合酶的结合和转录活性。
5. 转录后的RNA处理:真核生物DNA转录产生的RNA分子还需要经过一系列的RNA处理过程。
这些处理过程包括RNA剪接、RNA修饰和RNA运输等。
RNA剪接是指将转录后的RNA链中的内含子剪除,并将外显子连接起来。
这样可以产生不同的转录变体,增加基因的功能多样性。
6. 转录速度:真核生物DNA转录的速度相对较慢,通常为几十到几百个核苷酸每分钟。
这是因为真核生物DNA转录需要多个转录因子的参与,并且还要经过转录后修饰等复杂过程。
相比之下,细菌中的DNA转录速度较快,可以达到几千个核苷酸每分钟。