
家禽维生素E营养研究进展
摘要:维生素E是动物机体内一种非常重要的脂溶性抗氧化剂,具有诸多的生物学功能。本文综述了维生素E的生物学特性、吸收代谢特点、生物学功能以及对家禽生长性能、繁殖性能、免疫功能、肉产品质量、缓解热应激等方面影响的最新研究进展。
关键词:家禽;维生素E 研究进展
前言
维生素E(Vitamin E,简称VE),又称生育酚,是人们最早发现的维生素之一。1922年,Evans和Bishop首次发现维生素E是影响大鼠正常繁殖必需的脂溶性膳食因子[1]。随后,Sure 做了相似的实验,并把这种物质命名为“维生素E”[2]。1936年,Evans等从麦胚油中分离出具有VE活性的晶体状化合物,由于化合物功能与生育有关,故命名为生育酚,英文名为tocopherol。1937~1938年间,化学家Fernholz首次提出了α-生育酚的准确化学式,即其化学结构式为异戊二烯的6-羟基杂满(苯并二氢吡喃)的衍生物。1938年,瑞士化学家Karrer 首次人工合成了这种维生素。1968年,美国食品和营养委员会正式确定维生素E是人类营养的必须成分。现在,人们不仅清楚了维生素E的结构与性质,而且对维生素E的吸收,转运,代谢机制也进行了大量研究。随着家禽养殖集约化程度的不断提高,以及社会对食品安全越来越多的关注,人们对禽肉、禽蛋等产品的要求越来越高,经过长期大量的研究,作为天然生物抗氧化剂能防止自由基对细胞和生物膜的破坏,维生素E不仅为家禽繁殖所必需,还可以促进家禽的生长,提高家禽的健康水平、肉产品品质等。本文对维生素E的结构与性质、吸收与代谢以及主要生物学功能做了介绍,并综述了维生素E对家禽生产性能、繁殖性能、免疫功能、肉产品质、抗应激能力等方面的最新研究进展,为完善安全高效的家禽配合饲料配方及推进家禽的健康养殖提供参考资料。
1 维生素E的生物学特性
1.1化学结构
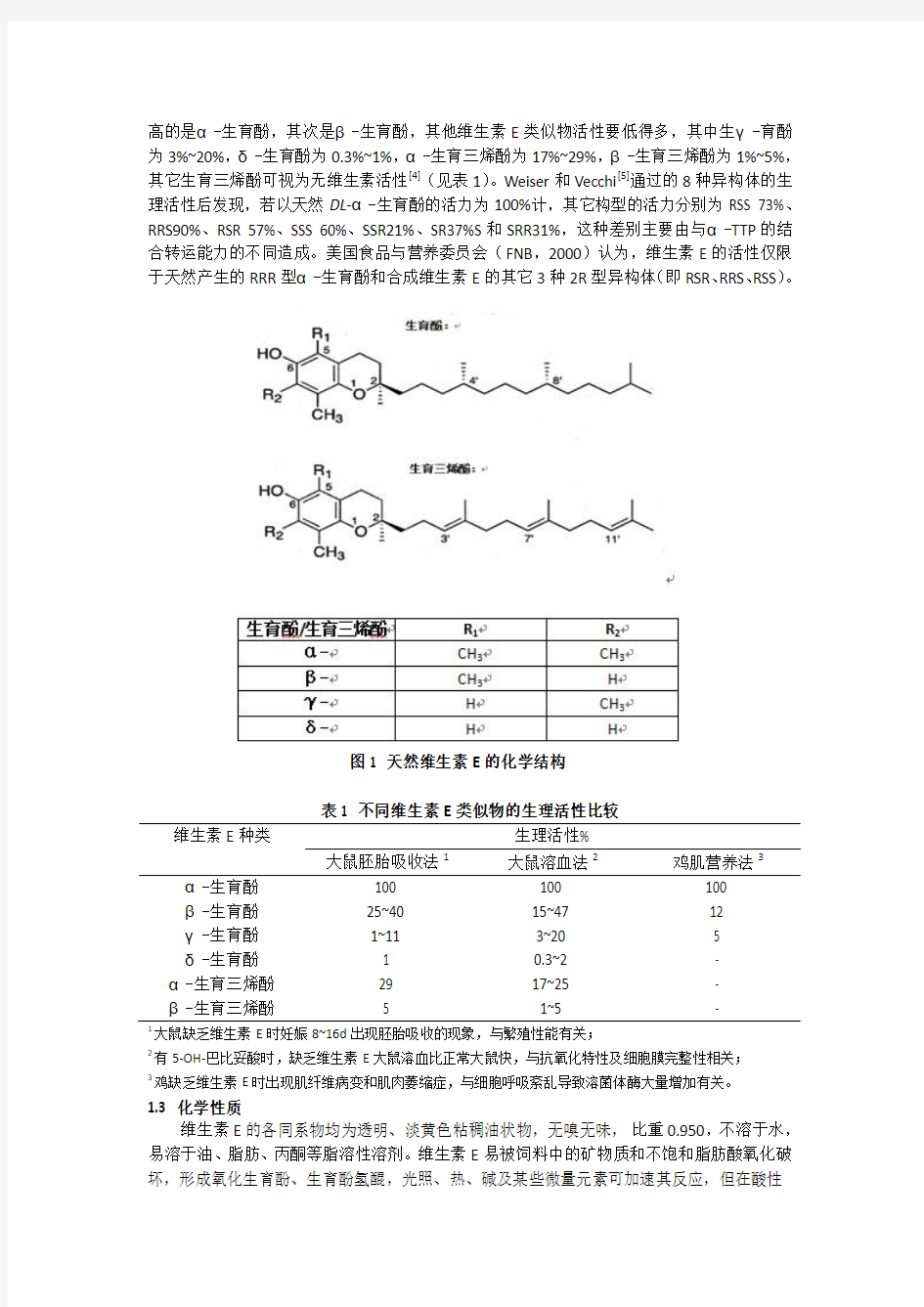
维生素E是苯丙二氢呋喃的衍生物,具有β-色满环和一个植基侧链,根据侧链结构的不同分为生育酚(tocopherol)和生育三烯酚(tocotrienol)。生育酚的侧链为一个饱和的16个碳原子的类异戊二烯,而生育三烯酚的侧链上有三个不饱和的键,位置在3’、7’、11’碳原子上。根据甲基数目和位置的不同,生育酚和生育三烯酚分别含有四种不同的形式,分别为α-、β-、γ-、δ-生育酚和α-、β-、γ-、δ-生育三烯酚(图1)。由于维生素E的植基部分有三个手性碳原子存在,手性碳原子上甲基的不同位置使得生育酚和生育三烯酚产生不同的消旋形式,根据R\S构型命名规则,人工合成的每一种类似物都有组分相等的8种异构体,如DL-α-生育酚有RRR、RSS、RRS、RSR、SSS、SSR、SRS和SRR等8种立体异构体。天然α-生育酚只有一种构型为RRR。
1.2生理活性
尽管这8种维生素E类似物结构相似,但由于与α-生育酚转运蛋白(α-TTP)的相对亲和力存在差异,因此在动物体内相应的生理学活性有很大的区别。维生素E在被小肠吸收后,需要经肝脏中特殊的转运蛋白转运到相应的部位才能发挥其生理作用,肝脏中存在特殊的α-TTP可选择性的转运α-生育酚,但缺乏其它维生素E类似物的转运蛋白。比较不同维生素E类似物与α-TTP结合的亲和力,以生育酚与TTP的相对亲和力为100%计,则β-、γ-、δ-生育酚与α-TTP的相对亲和力分别为38.1%、8.9%、1.6%,其余更低,故它们的生理活性也就越低[3]。通过各种试验观察不同形式的维生素E对生物功能的影响发现,活性最
高的是α-生育酚,其次是β-生育酚,其他维生素E类似物活性要低得多,其中生γ-育酚为3%~20%,δ-生育酚为0.3%~1%,α-生育三烯酚为17%~29%,β-生育三烯酚为1%~5%,其它生育三烯酚可视为无维生素活性[4](见表1)。Weiser和Vecchi[5]通过的8种异构体的生理活性后发现,若以天然DL-α-生育酚的活力为100%计,其它构型的活力分别为RSS 73%、RRS90%、RSR 57%、SSS 60%、SSR21%、SR37%S和SRR31%,这种差别主要由与α-TTP的结合转运能力的不同造成。美国食品与营养委员会(FNB,2000)认为,维生素E的活性仅限于天然产生的RRR型α-生育酚和合成维生素E的其它3种2R型异构体(即RSR、RRS、RSS)。
图1 天然维生素E的化学结构
表1 不同维生素E类似物的生理活性比较
维生素E种类生理活性%
大鼠胚胎吸收法1大鼠溶血法2鸡肌营养法3α-生育酚100 100 100
β-生育酚25~40 15~47 12
γ-生育酚1~11 3~20 5
δ-生育酚 1 0.3~2 -
α-生育三烯酚29 17~25 -
β-生育三烯酚 5 1~5 -
1大鼠缺乏维生素E时妊娠8~16d出现胚胎吸收的现象,与繁殖性能有关;
2有5-OH-巴比妥酸时,缺乏维生素E大鼠溶血比正常大鼠快,与抗氧化特性及细胞膜完整性相关;
3鸡缺乏维生素E时出现肌纤维病变和肌肉萎缩症,与细胞呼吸紊乱导致溶菌体酶大量增加有关。
1.3 化学性质
维生素E的各同系物均为透明、淡黄色粘稠油状物,无嗅无味,比重0.950,不溶于水,易溶于油、脂肪、丙酮等脂溶性溶剂。维生素E易被饲料中的矿物质和不饱和脂肪酸氧化破坏,形成氧化生育酚、生育酚氢醌,光照、热、碱及某些微量元素可加速其反应,但在酸性
环境及酯化后较为稳定。对可见光稳定,但可被紫外光破坏。
1.4维生素E的来源
植物能合成维生素E,且所有谷类粮食中都含有丰富的维生素E,特别是种子胚芽中。绿色饲料、叶和优质干草也是维生素E很好的来源。植物油是各种生育酚的主要来源,但其所含生育酚的种类和数量不同。除β-生育酚主要含于棉籽油中外,α-生育酚、γ-生育酚和δ-生育酚都不同程度出现于各种植物油中。生育三烯酚也存在于大多数植物油中,但其含量远低于生育酚,经常被忽略,一个例外是棕榈油含有高水平的生育三烯酚。在浸提后的油饼类原料中维生素E较缺乏。
虽然天然维生素E具有其天然活性,但是其天然RRR型α-生育酚含量有限,不能满足实际需要。而且然维生素E极不稳定,在饲料中易氧化,另外市场价格相对较高,故目前在饲料中广泛使用的是人工合成的性质稳定的DL-α-生育酚醋酸酯。维生素E有2种常用单位:其中1 mg DL-生育酚醋酸酯=1 IU DL-生育酚醋酸酯,1 mg D-生育酚=1.49 IU D--生育酚。
2 维生素E的吸收与代谢
2.1维生素E的吸收
脂溶性维生素通常是伴随着脂肪的吸收而吸收。摄入的天然维生素E和脂质、胆盐可以形成微胶囊粒,被动扩散进入肠道粘膜[6],而维生素E酯类衍生物在被吸收前必须被胰脂肪酶水解[7]。然后在粘膜细胞的高尔基体内形成乳糜微粒,并通过胞吐作用进入淋巴系统。通过淋巴吸收的α-生育酚,约99%是以乳糜微粒的形式吸收的,而且主要是在十二指肠吸收的。乳糜微粒包裹的α-生育酚被吸收进入循环系统后,于运输的过程中在脂蛋白脂肪酶(LPL)作用下,释放出一部分生育酚而被肝外组织,如脂肪组织,骨骼肌等组织吸收利用[8,9],大部分被乳糜微粒包裹的生育酚进入肝脏,主要分布在主质细胞,少量分布在内皮细胞,星状细胞,枯否氏细胞[10]。由于肝脏中存在特异性的α-TTP可选择性的转运α-生育酚,故在肝脏中TTP可优先与α-生育酚结合,然后以囊泡的形式在细胞膜中转运,参与极低密度脂蛋白(VLDL)的合成,再通过血液系统从肝脏把α-生育酚运往各个组织。在转运的过程中,约50%的VLDL又进入肝脏,其余一半的VLDL被血液系统中的脂蛋白脂肪酶(LPL)降解成低密度脂蛋白(LDL),其中的α-生育酚为肝外组织摄取利用,过量的α-生育酚以及VLDL分解后的多余成分被转入高密度脂蛋白(HDL),然后循环回到肝脏。由于选择性运输的存在,其他维生素E类似物则很少在肝脏细胞中停留,多数通过胆汁、尿液或其他途径排泄出体外。
在α-生育酚进入组织器官的过程中,B类Ⅰ型清除剂受体(scavenger receptor class B typeⅠ,SR—BⅠ)对α-生育酚从膜蛋白到特定组织的转运具有重要的作用。有研究表明,敲除SR—BⅠ基因大鼠的睾丸、卵巢、肺部和大脑的α-生育酚的浓度显著下降,而细胞膜的α-生育酚浓度却大大增加[11]。Reboul 等[12]研究发现,使用SR-BⅠ抑制剂可显著降低VE 的吸收,SR-BI的过量表达可提高α-生育酚的生物学利用率。这些结果也暗示α-生育酚的吸收转运并非完全是一个被动扩散的过程。且Anwar[13]等的体外细胞脂质吸收模型也证明了这一点。动物体中还有其他一些因子参与α-生育酚转运过程,例如3种TAPs(tocopherol-associated protein),即TAP1/SPF(SEC14L2),,TAP2 (SEC14L3),和TAP3(SEC14L4),三者均属于Sec 14脂结合蛋白超家族[14]。目前到底有多少相关蛋白、载体或受体参与了生育酚的选择性吸收、转运,以及它们是如何调控α-生育酚的选择性吸收、转运的作用机制还没有完全清楚。
2.2维生素E的代谢
肝脏是VE储存的重要器官,也是VE代谢的主要场所。维生素E作为重要的抗氧化物,与自由基作用,失去氢原子形成氧化产物生育酚自由基。生育酚自由基有4种去向:a.被体内其他抗氧化物如维生素C,还原为非氧化形式;b.再与另一自由基作用,进一步氧化生成非自
由基的生育醌;c.与其他生育酚自由基形成没有活性的物质,如生育酚二聚体、三聚体;d.作为过氧化物,氧化其他脂肪酸。早在20世纪50年代,Smion等[15]在尿样中分离鉴定了生育酸(tocopheronic acid)和生育酸内酯(tocopheronolactone),研究发现它们是生育酚在体内氧化分解的最终代谢物,这两种代谢物又称作Smion代谢物。目前解释这两种代谢产物来源及代谢机制的理论主要有两种[16],一是生育酚苯并二氢吡喃母环分解机制,该理论推测生育酚苯并二氢吡喃母环的降解是通过与自由基氧化反应生成相应的生育酚醌或生育酚氢醌,然后以生育酸或生育酸内酯的形式排出体外;二是生育酚侧链基团降解机制,在尿中检测到生育酚的非氧化产物α-CEHC(2,5,7,8-四甲基-2-羧基-6-羟基苯并氢化吡喃)和γ-CEHC,α-CEHC 在其苯并二氢吡喃母环被氧化后也可以Smion代谢物的形式排除体外,即生育酚也可通过细胞色素介导的ω-氧化途径和β-氧化途径进行的非抗氧化途径分解代谢。
3维生素E的生物学功能
3.1抗氧化作用
生物膜对细胞生命活动是很重要的,细胞、细胞器膜结构的完整性是发挥其正常功能的基础。由于生物膜上富含多不饱和脂肪酸(PUFA)极易受到自由基的攻击而发生脂质过氧化反应。所谓脂质过氧化反应是一种由自由基介导的链式反应过程,其结果是导致PUFA的断裂、氧化并产生有毒性的LOOH和其他代谢中间产物,其反应的终产物丙二醛(MDA)可以和膜上的蛋白、磷脂上的氨基交联,生成西夫(Shiff)碱,导致细胞变形性改变,生物膜活性降低,从而影响膜的物理化学性质,使细胞的多种功能受到损害[17]。维生素E作为动物体内重要的抗氧化剂,其抗氧化作用主要是:阻止自由基反应的进行,从而保护细胞膜磷脂和血浆脂蛋白中的多不饱和脂肪酸免受氧自由基的攻击,保证细胞膜及细胞器膜结构和功能的完整性[18];增强谷胱甘肽过氧化物酶(GSH-Px)、过氧化氢酶(CAT)等酶的活性[19,20]。
在维生素E的八种形式中,α-生育酚是最具有活性的,能直接参与中断脂质过氧化的过程,发挥其抗氧化的作用。其抗氧化作用的机理如图2[21]所示。具体地说,在脂质过氧化反应中,首先由外界或机体自身的自由基(X·)在光照、热能、微量过渡金属离子等作用下引发,也可以由其它自由基或自由基产物引发。自由基首先从不饱和脂肪酸分子上夺取一个氢,使脂质生成脂质自由基(L·),由于氧的存在,过氧化脂质自由基跟着生成过氧化脂质自由基(LOO·)(反应几乎是扩散型,k=108~109 M-1s-1)。如果过氧化自由基不被清除,它将进一步扩增自由基链,重复反应(1a)~(2),如此循环不止;而各种自由基连锁反应的结果使LOOH积累,而完好的LH不断减少,最后因各种自由基相互结合为稳定的化合物而使反应终止。然而,在α-生育酚存在的情况下,α-生育酚迅速与过氧化自由基反应(速度k=105~106 M-1s-1),结合形成稳定的化合物,而自身形成较稳定的生育酚游离基,反应速度远大于过氧化自由基与不饱和脂肪酸之间的反应。因此,α-生育酚能够在特定反应位置阻断自由基反应链的延续,从而终止脂质过氧化反应,这一过程,主要依赖β-色酮环上第6位的羟基提供电子给自由基。此时,尽管α-生育酚将过氧化脂质自由基(LOO·)转变成了脂质氢过氧化物(LOOH),后者仍然是一种很具氧化活性的物质,依然潜在氧化威胁。α-生育酚也不能进一步清除该物,只能由其它抗氧化物予以清除(见图2反应5~8)。即虽然VE具有很强的抗氧化、清除自由基的能力,但是它有局限性,VE必需结合其它抗氧化物或机体自身抗氧化酶系统才能达到彻底清除机体氧化的威胁。
3.2维持生育功能
维生素E是动物维持生育必不可少的营养物质,在畜禽繁殖上发挥重要的作用,研究表明维生素E使垂体前叶促性腺分泌细胞亢进,促性腺激素分泌增加,促进性腺发育,调节性机能, 从而提高卵巢机能,增加卵泡黄体细胞,增强动物的繁殖能力。维生素E也能影响类固醇激素、前列腺素PG2的合成,促进精子的形成和增强其活力,促进胚胎发育[22]。缺乏时,雄性家禽睾
丸变性萎缩,精子运动异常甚至不产生精子,造成不育症,种蛋孵化率降低。但是过量的维生素E抑制前列腺素的合成,影响雌性动物卵巢健康,从而影响繁殖性能[23]。维生素E还可以通过抗氧化作用保护生物膜的的正常功能,从而维持生殖器官功能的正常发挥。有研究者认为成年鸡即使采食低剂量的维生素E也不会表现出临床病理症状,但种蛋孵化率下降很明显,在孵化早期(孵化的第4天易出现胚胎死亡,其原因是种蛋孵化的第3~4天,正值尿囊形成,胚胎由无氧呼吸转变为有氧呼吸,容易产生大量氧自由基,引起氧化应激导致胚胎死亡[24]。
注:LH——多不饱和脂肪酸;L·——脂质自由基;LOO·——脂质过氧自由基;LOOH——过脂质氢过氧化物;α-TO·——α-生育酚氧化自由基;k——常数,单位;LTA——A星白细胞三烯;LTB——B型白细胞三烯;HPETE——过氧羟基二十四碳四烯酸;HETE——羟基二十碳四烯酸;HPODE——过氧羟基十八碳二烯酸;HODE——羟基十八碳二烯酸(引自Brigelius-FldheR[22])
图2 脂质过氧化作用及α-生育酚抗脂质过氧化反应
3.3免疫调节功能
维生素E对免疫系统的调节作用是近年来研究的热点。研究确认,维生素E可增强机体免疫力和对疾病的抵抗力,而维生素E的免疫调节作用机制主要是它的抗氧化作用。维生素E的主要作用是保护细胞膜及细胞器膜不被氧化破坏,而动物免疫应答主要是在细胞膜水平发生的。缺乏VE的膜脂质过氧化反应可引起膜流动性改变,从而影响淋巴细胞膜上受体分布,改变淋巴细胞对靶细胞或抗原的识别与结合。另一重要机制是维生素E可以作为免疫调节物来调节细胞素、前列腺素、凝血素及促细胞生长素合成,前列腺素E2(PGE2)是花生四烯酸的氧化代谢产物,它可以作为负向免疫调节剂抑制一些淋巴细胞的合成,而维生素E与多不饱和脂肪酸(PUFA)的关系即维生素E抑制不饱和脂肪酸的氧化,从而使前列腺素E2(PGE2)浓度降低来促进淋巴细胞的合成,提高机体的免疫能力。王丽等[25]报道,在基础日粮中添加2.43 IU/kgBW维生素E,对围产期奶牛的免疫功能具有较好的效果,在产后7 d奶牛血液中三种免疫球蛋白(IgG、IgM、IgA)含量显著高于添加低VE水平的日粮。
3.4细胞信号转导以及基因表达调控功能
维生素E除了直接的抗氧化功能外,还有特异性的分子功能,但主要是α--生育酚。它能抑制蛋白激酶C的活性,蛋白激酶C在血小板增殖和分化中起重要作用[26]。维生素E还可以上调胞浆磷脂酶A2、环氧合酶-1的表达,增强其活性[27],维生素E在体内还可以影响基因的转录与表达。α--生育酚可以负调控清除剂受体SR-A和CD36基因的转录,也微弱的影响相关组织生长因子(CTGF)[28]和α—原肌球蛋白的表达[29]。最近研究发现,TAP是一种配体依赖型转录因子,而α--生育酚为其最有活性的配体[30],但TAP是否影响SR-A、CD36、CTGF、α—原肌球蛋白的转录还不清楚。
4维生素E在家禽生产上的应用
4.1维生素E对家禽生长性能的影响
维生素E能对动物的生长起着重要的作用。呙于明等[31]综合了试验中生长性能认为,在含α-生育酚13mg/kg的玉米-豆粕-豆油日粮中添加维生素E 5 mg/kg以上,使其总量达到18 mg/kg以上,可明显的促进生长,添加维生素E 50 mg/kg以上使其总量达到63 mg/kg以上,可显著改善饲料利用率。还认为,玉米-豆粕-豆油日粮中添加维生素E10 mg/kg以上,对于维持较高水平的血浆α-生育酚含量是必要的。日粮中添加200 mg/kg维生素E可提高肉鸡日增重和饲料转化率,提高家禽生长性能(Rebole等,2006;李新花等,2006;李绍钰等,2001;李彦等,2008)[32-35]。但李绍钰等[34]报道,肉仔鸡日粮中添加高剂量维生素E 对增重量、饲料报酬和屠宰性能没有显著影响。
4.2维生素E对家禽繁殖性能的影响
4.2.1维生素E对家禽产蛋的影响
蛋的形成涉及雌激素在肝脏诱导卵黄前体蛋白的合成和卵黄前体蛋白在卵巢中的吸收。而卵黄前体蛋白源(vg)是由肝脏合成,其释放又需要经过细胞膜,维生素E作为细胞膜的抗氧化剂可保护细胞膜不受破坏,保证其正常的运输功能,使vg能正常的释放出来,以维持较高的产蛋量。Lin YF等[36]在产蛋期往台湾土种母鸡的玉米-豆粕饲粮中分别添加0、40、80、120和160mg/kg的dl-α-生育酚,对鸡的初产日龄无影响,但可显著降低料蛋比和提高产蛋量,添加80mg/kg的试验组效果最好。田河等[37]研究表明,添加100mg/kg维生素E可以显著降低鸡料蛋比并提高产蛋率。马兆臣[38]研究表明,饲料中添加350mg/kg和450mg/kg 维生素E的处理组的皖西白鹅的产蛋率显著高于添加0、100mg/kg维生素E的处理组。以上研究表明,维生素E可以增加鸡蛋、鸭蛋中维生素E的含量,降低料蛋比,提高鸡、鹅产蛋量和产蛋率。
4.2.2维生素E对家禽受精的影响
Hennig A等[39]推荐对种母鸡添加20mg/kg维生素E,可较好的提高受精蛋的数量。LinYF 等[36]研究表明,向产蛋期种鸡饲料中添加80mg/kg dl-α-生育酚时,受精率相比对照组提高7.7%。田河[37]研究表明,添加100、175、250mg/kg维生素E,鸡蛋受精率均高于对照组,分别提高2.11%、0.05%、0.15%。赵国先等[40]报道在常规日粮与非常规日粮中添加180 mg/kg 维生素E,均可显著提高种蛋受精率。陈岩锋等[41]报道在含13mg/kg维生素E的鸭基础日粮中对各处理组分别添加0、10、20、40、60mg/kg维生素E,随添加量的增加受精率有上升的趋势。马兆臣[38]研究表明,饲料中添加350和450mg/kg维生素E处理组的皖西白鹅的受精率高于添加0、100mg/kg维生素E的处理组。以上研究表明,添加适当的维生素E可以提高家禽的受精率。
4.2.3维生素E对家禽孵化的影响
Lin YF等[36]研究表明,添加80mg/kg dl-α-生育酚时,鸡蛋孵化率相比对照组提高了13.4%,达到各处理组中的最高值。赵国先等[40]报道在常规日粮中添加180~300 mg/kg,非常规日粮中添加300 mg/kg维生素E可显著降低死胎率,显著提高孵化健雏率。许万祥[42]
研究表明,VE缺乏时可导致入孵后胚胎发育受阻及循环系统紊乱。陈岩锋等[41]报道,在含13mg/kg(估值)维生素E的种鸭基础日粮中添加40mg/kg左右的维生素E的处理组的孵化率显著高于未添加组。马兆臣[38]研究表明,饲料中添加350mg/kg和450mg/kg维生素E的处理组的皖西白鹅的孵化率高于添加0、100mg/kg维生素E的处理组。以上研究表明,维生素E可以提高家禽的孵化率。
4.2.4维生素E对家禽精液品质的影响
Lin YF等[43]对台湾的土种斗鸡的豆粕-玉米型饲粮分别进行添加0、20、40、80、160mg/kg dl-α-生育酚,结果表明39周龄后的公鸡在给予的dl-α-生育酚在40~160mg/kg之间时精子活力最大,在给予80mg/kg dl-α-生育酚时精液浓度最大,而缺乏dl-α-生育酚超过39周会导致精液品质下降。Biswas A等[44]将135只印度Kadaknath公鸡分成9组,设10mg/kg(T1)、100mg/kg(T2)、200mg/kg(T3)三个水平,结果表明,添加10mg/kg维生素E可提高精液中谷草转氨酶(GOT)和谷丙转氨酶(GOP)活性,添加100mg/kg维生素E可减少死精、弱精,提高精液中葡萄糖和维生素E含量。以上研究表明,添加适当维生素E可以增加精子的抗氧化能力,提高家禽精液品质。
4.2.5维生素E对家禽生殖器官的影响
维生素E可通过调节垂体前叶分泌促性腺激素,调节鸡的性功能,刺激睾丸组织的生长发育,增强公鸡的睾丸生精能力;维生素E在母鸡体内具有保护生殖器官、刺激卵巢发育而增加排卵数的功能。陈鑫[45]对试验鸭饲料进行添加20、220、420mg/kg的处理,试验结果表明,随维生素E添加量的增加,输卵管重量、输卵管长度及卵巢的重量有增加的趋势。4.3维生素E对家禽肉质的影响
目前研究认为,日粮中添加VE可提高动物组织线粒体、微粒体中VE含量,提高亚细胞结构对铁诱导氧化反应的稳定性,进而提高畜禽肉品及其加工产品在加工过程中的氧化稳定性,延长商品货架寿命,获得显著经济效益。
4.3.1维生素E对肌肉的氧化稳定性的影响
VE在细胞膜中可解除自由基,从而阻止肌细胞膜磷脂中脂肪酸的氧化。研究表明,日粮中添加VE能增加其在组织中的含量,降低脂类氧化水平,提高鸡肉氧化稳定性[46-48]。肉品稳定性与组织中α-生育酚含量有关,组织中α生育酚浓度受日粮中维生素E添加水平及其添加时间长短影响[49]。李同树等[50]报道,在鲁西黄鸡的基础日粮中添加50mg/kgα-生育酚醋酸酯,鸡肉的α-生育酚含量显著增加,能达到改善肉质的效果。Nacim等[51]报道,肉鸡屠宰前20天向饲粮中添加100~300 mg/kg的维生素E,机体VE含量显著高于对照组(VE 添加量为25 mg/kg)。脂肪氧化水平可由氧化硫巴比妥酸反应物(TBARS)来衡量。Yuming Guo 等[52]报道,提高日粮VE水平能显著降低4℃下贮存4 d的腿肉TBARS值,且该值与日粮VE 水平呈显著负相关。Bartov等[53]研究不同日龄期间饲喂高浓度VE对7周龄肉仔鸡腿肉氧化稳定性的影响,结果表明,贮存后肉的TBARS值与VE添加量呈极显著负相关,补饲VE的鸡肉稳定性显著提高,且0~7周连续补饲显著高于阶段式补饲。因此,饲料中添加VE能有效抑制肌肉脂肪氧化,保持肉质稳定。
4.3.2维生素E对肉品颜色的影响
肉品颜色虽然与其食用品质无直接关系,但它却是肌肉内部的生理学、生物化学和微生物学变化的外部表现,也是消费者判断肉质和鲜度的主要依据。肌肉失色是肌间脂肪发生脂质过氧化与氧合肌红蛋白被氧化为正铁肌红蛋白的综合作用,而脂质氧化的产物可进一步催化氧合肌红蛋白向正铁肌红蛋白的转化,使肌肉呈现褐色。肌肉失色的速度与天然色素氧化速度、氧消耗量、高铁肌红蛋白降低系统的效率有关。维生素E可通过直接阻止脂质氧化并间接延缓了氧合肌红蛋白的氧化,从而起到了保护肉色、延长货架期的作用。李同树等[50]报道,饲喂添加维生素E的饲粮,鲜肉在4℃的条件下保存24h,其肉色的褪色率试验组小于对
照组,这说明维生素E能改善肉色,并且对肉色具有保护作用。然而Nacim等[51]报道,屠宰前20天向饲粮中添加200~300 mg/kg的维生素,E不能增强腿肌肉色的稳定性,但可提高腿肌的新鲜度。
4.3.3维生素E对肉品滴水损失的影响
滴水损失是量化肌肉系水力的指标。肌肉的系水力直接关系到肉品的质地、风味和组织状态,而且肌肉的失水率越高,其总色素和可溶性营养成分流失越多。维生素E作为一种脂溶性抗氧化剂,能够防止肌细胞膜上的磷脂被氧化,维持了细胞膜的完整性,防止胞浆外流,从而减少滴水损失。日粮中添加VE可提高肌肉系水力,并可明显降低Ca2+的释放量,降低糖酵解速度,抑制线粒体中磷脂酶A的活性,从而防止PSE肉产生。周林[54]、李同树[50]研究表明,在肉鸡日粮中添加VE可减少胸肌滴水损失,并降低肌肉失水率,改善肌肉嫩度。
4.3.4维生素E对肉品风味的影响
风味是肉品的另一个重要特征。脂肪氧化是产生肉质腐败气味的主要原因。已证实肌肉系统中,脂肪是发生氧化酸败的中心位点。肌肉和脂肪中过氧化起始于细胞膜和亚细胞器的膜水平,线粒体和微粒体膜上有富含极易氧化的磷脂,磷脂具有高度不饱和特性,双键易断裂,产生各种氧化产物如醛、酮、醇等复杂混合物,这些产物是产生酸败气味的主要原因。维生素E可以对因水解磷脂使肉产生异味的磷脂酶A2有抑制作用,从而减少异味的产生[55]。
C. Narciso-Gaytán等[56]报道,向肉仔鸡饲粮中添加200~400mg/kg的维生素E,可显著降低储藏和加工过程中腿肌脂质的氧化,过氧化物值(PV)显著减小。并且这种作用在鸭上也得到了验证[57]。
4.4维生素E对家禽免疫功能的影响
VE是细胞膜和亚细胞膜抗自由基的第一道防线,对家禽的免疫系统具有重要作用,能够有效促进雏禽免疫器官脾脏、胸腺和法氏囊生长发育,增强机体的体液免疫和细胞免疫反应,从而提高家禽的健康水平,增强抗病能力,减少因生产中不良因素引起的死亡率增加、生产性能下降等情况的发生,提高经济效益。
4.4.1 VE与体液免疫
日粮中添加VE可增强机体体液免疫反应,有利于淋巴细胞特别是辅助性T淋巴细胞增殖和T、B淋巴细胞协同作用及巨噬细胞增殖,提高血液中免疫球蛋白的水平,增强对各种抗原和疫苗的抗体产生能力,具体表现在日粮中添加VE时抗体滴度增加,新城疫抗体效价显著提高。研究表明,在全价营养条件下给予肉鸡高剂量的VE有益于机体免疫功能,且这种作用对30 d肉鸡的效果更为明显,血清中IgG含量、淋巴细胞反应性、新城疫免疫应答及白细胞数的变化规律也基本一致[58]。文杰等[59]报道,日粮中添加VE可明显提高肉仔鸡血清H I抗体滴度,表明添加VE可以提高肉仔鸡的体液免疫水平,并认为其主要是通过激活原发性免疫反应来调节IgG的生物合成,也可通过激活B细胞而促进IgG的分泌。当育成金定蛋鸭饲粮中添加15、20 IU/kg VE时,血清中的免疫球蛋白(Ig G、Ig M)、IL 2和甲状腺素(T 3)的浓度较高[60]。Gore等[61]在入孵18 d的鸡胚中注射10 IU的VE,分别在孵化后7 d和14 d 注射绵羊红细胞,7 d时机体内总的免疫球蛋白、IgM、巨噬细胞所吞噬的绵羊红细胞数均有所增加;而14 d时则这些指标均高于对照组。
4.4.2 VE与细胞免疫
细胞免疫主要是由T细胞介导,是机体免疫应答的一个重要组成部分。研究证实VE可增强家禽的细胞免疫反应。肉仔鸡中粮中添加0、47和87 mg/kg VE时,不影响胸腺和脾、B细胞或巨噬细胞的比例,而特异影响T细胞的分化,7日龄时高水平VE(47和87 mg/kg)可增加CD4+/CD8+,饲喂高含量VE日粮的肉仔鸡可能优先生成CD4+淋巴细胞,而CD4+淋巴细胞是抗体产生过程中的主要辅佐细胞[62]。文杰[59]报道,日粮中高水平VE(80 mg/kg)可以提高28日龄肉鸡血液淋巴细胞转化率。VE缺乏时,嗜中性粒细胞膜中的脂质过氧化物增多,细
胞在释放出过多的过氧化氢后,寿命减短,嗜中性粒细胞的功能受损,淋巴细胞中有丝分裂反应减慢,血液中T辅助细胞数量减少,抑制性T细胞增多,免疫功能下降。
4.4.3 VE与免疫器官
脾脏、法氏囊、胸腺是家禽的主要免疫器官。研究表明,VE能够促进这些免疫器官的发育,其能促进脾脏细胞的分裂,使脾脏重量显著增加[63]。Gerrit等[64]报道饲料中添加VE 可以增加胸腺和法氏囊的重量,从而提高机体的免疫功能。李彦[35]在肉鸡日粮添加不同水平的VE和VA,结果发现增加日粮VE和VA的水平不影响肉鸡的脾脏和法氏囊指数,胸腺指数显著提高,这可能与VE添加量和饲养条件有关。VE缺乏时,家禽胸腺、脾脏和法氏囊的生长发育明显受抑制,并导致这些组织的损伤性变化[65]。
4.5维生素E对家禽热应激的影响
在现代养殖业中尤其在南方,家禽面临着严重的热应激的挑战。家禽处于热应激状态时,主要表现为采食量减少,消化率下降;维生素%的摄入量相对减少,而机体内代谢却增强,尤其是骨组织和肌肉分解代谢加剧,随后增重、产蛋下降,蛋品质变劣,繁殖力下降和死亡率上升等。
大量研究表明,维生素E可缓解热应激对家禽的不良影响,恢复和调节代谢,降低家禽的死亡率。在热应激时维生素E有助于维持和提高抗氧化酶活性、抗体水平,增加抵抗力,促进采食,表现出明显的抗应激效果。傅德顺[66]报道,在热应激状态下,蛋鸡日粮中维生素E水平从10 IU/kg提高到30~50IU/kg,蛋鸡产蛋率提高了1.5%~5.5%。王晓霞等[67]在蛋鸡日粮中添加不同水平VC和VE,结果表明,热应激期日粮添加1 000 mg/kg VC及250 mg/kg VE 可显著提高产蛋率,降低破软蛋率;显著提高血清中SOD及全血中GSH PX的活性。Vakili等[68]报道,在32℃条件下向含有不饱和脂肪酸的肉鸡日粮中添加100IU的维生素E和50mg/kg 锌,较对照组(不添加维生素E和锌)可显著提高肉鸡的日增重和饲料转化率,但对料肉比影响不显著。刘铀等[69]试验表明,提高日粮中维生素E水平可显著降低血清皮质醇含量,缓解机体的应激状况,同时指出,热应激期间最大程度的维持产蛋量的维生素E适宜剂量应控制为200~300mg/kg。方洛云等[70]试验也证明在饲料中添加维生素E明显降低肉仔鸡血清皮质醇的含量,缓解机体热应激状况。孙长勉等[71]则发现维生素E也可减轻热应激对肉鸡血清T3和T4浓度的影响,维持T3和T4浓度的相对稳定。以上研究表明饲粮中添加适量的VE可有效的缓解家禽热应激。
5家禽对维生素E的需要量
由于维生素E与其他许多营养素存在相互作用,如饲料中脂质的类型及含量、脂溶性维生素A的水平影响维生素E在小肠的吸收;与维生素C和硒在抗氧化作用方面存在协同作用;还有一些矿物元素通过影响抗氧化酶的活性影响机体对维生素E的需要,而这些作用不容忽视。另外,目前研究确定家禽对维生素E需要量主要以其生产性能(主要有生长速度、饲料转化率、产蛋率等)获得最大响应量时的需要量,而二者并非简单的线性关系。所以评估家禽对维生素E 的需要量非常困难。
根据早期的研究,美国、中国等确立了预防家禽维生素E典型缺乏症脑软化和渗出性素质的需要量,其中蛋鸡和肉鸡维生素E需要量在5~10 mg/kg,北京鸭在10~20 mg/kg(见表2,表3),该需要量相对较低,既未包括饲养实际中和饲料加工条件下的安全含量,也未包括应激、亚临床病以及维生素E的额外添加效果。通常在实际生产中会对家禽日粮进行强化,肉鸡如典型添加量是20~30 mg/kg。Shen等[72]研究表明,来航生长蛋鸡及肉仔鸡对日粮(含脂肪3.6%,亚油酸0.4%)维生素E的需要量为12.5~25 mg/kg;Bartov等[53]认为,24 mg/kg 维生素E能满足肉仔鸡最佳生产性能的需要。王爽等[73]研究表明,维生素E可提高0~14日龄北京鸭生产性能,以日增重、料肉比为评价指标,北京鸭维生素E最低需要量为10 IU/kg,
与NRC(1994)推荐量一致。然而,有大量研究表明[31,33,34,59],饲粮中添加100~200 mg/kg VE 可以显著提高前肉仔鸡的平均日增重和饲料转化率。
但在不同的生产条件下,对于不同的生产目的,研究工作者也进行了大量的研究,Nacim Zouari等[51]报道,肉鸡屠宰前20天向饲粮中添加100~300 mg/kg的维生素E,机体VE含量显著高于对照组(VE添加量为25 mg/kg),可显著降低储藏和加工过程中肌肉脂质的氧化。赵国先等[40]报道在常规日粮中添加180~300 mg/kg,非常规日粮中添加300 mg/kg维生素E可显著降低死胎率,显著提高孵化健雏率。张小纯等[74]报道,饲粮中添加维生素E能显著提高骡鸭的产肝性能,并且维生素E添加量为40 mg·kg-1时,骡鸭平均肝重、一级肝所占比例、肝体比最高,料肝比最低。傅德顺[66]实验表明,在热应激状态下,蛋鸡日粮中维生素E水平从10 IU/kg提高到30~50IU/kg,蛋鸡产蛋率提高了1.5%~5.5%。而刘铀等[69]报道热应激期间最大程度的维持产蛋量的维生素E适宜剂量应控制为200~300mg/kg。上述报道表明,不同的生产目的和生产条件下,对家禽维生素E需要量的报道不尽一致,差别较大。
因此,应根据家禽的生长阶段、日粮组成以及环境应激等因素寻求一个最佳添加量,使之产生所期望的效果。
表2 6国蛋鸡维生素E需要量(m g/kg)
国别蛋雏鸡(周龄)育成鸡(周龄)产蛋鸡(周龄)种母鸡公布年份
中国10(0~6)5(7~20) 5 10 1986
美国10(0~6)5(7~开产) 5 10 1994 澳大利亚15(0~7)5(8~18) 5 5 1987 日本10(0~9)5(10~20) 5 10 1992
前苏联10(1~8)5(9~21) 5 10 1985
法国10(0~6)10(7~20)10 25 1990
表3 6国肉鸡及鸭维生素E需要量(m g/kg)
国别肉仔鸡(周龄)肉种鸡生长鸭种鸭公布年份
中国10(0~8)- 15~20 10 1986
美国10(0~8)- 10 10 1994 澳大利亚5(0~8)- - - 1987 日本10(0~出栏)- - - 1992
前苏联5~10(1~出栏)10 5 5 1985
法国20~30(0~8)- 20~25 10 1990
6结语
维生素E作为一种必需营养素对家禽的肉品质和机体的抗氧化、免疫及繁殖等功能具有重要影响。因此,随着家禽养殖集约化程度和人们对食品安全要求的不断提高,日粮中合理的添加维生素E对于改善动物健康状况,指导生产和提高经济效益有重要意义。但家禽对维生素E的最适需要量还需要深入的研究。
参考文献
[1]EVANS H M,BISHO P K S. On the existence of a hitherto unrecognized dietary factor essential
for reproduction[J].Science.1922,56:650 651.
[2]SURE B. Dietary requirement for reproductionⅡ: The existence of a specific vitamin for
reproduction[J].Biol Chem,1924.58:693 709.
[3]Hosomi,A.;Arita,M.;Sato,Y.et al. Affinity of a-tocopherol transfer protein as a determinant of
the biological activities of vitamin E analogs[J]. FEBS Lett .1997,409:105–108.
[4]Drevon C A. Absorption, transport and metabolism of vitamin E[J].Free Rad. Res. Commun.
1991,14:229-246.
[5]Weiser H , Vecchi M. Stereoisomers of tocopherol acetate.Ⅱ. Biopotencies of all eight
stereoisomers, individually or in mixtures, as determined by rat resorption-gestation test[J].
Int. J.Vit. Nutr. Res.1982,52:351-370.
[6]Traber,M.G.,Goldberg,I.,Davidson,E.,Lagmay,N.,Kayden,H.J. Vitamin E uptake by human
intestinal cells during lipolysis in vitro[J].Gastroenterology ,1990,98,96–103.
[7]Scherf H, Machlin L J, Frye TM, et al Vitamin E biopotency: comparison of various natural
derived and chemically synthesized tocopherols[J]. Anmi Food SciTech,1996,59:115.
[8]Traber,M.G.,Olivecrona,T.,Kayden,H.J. Bovine milk lipoprotein lipase transfers tocopherol to
human fibroblasts during triglyceride hydrolysis in vitro[J].J.Clin.Invest, 1985,75,1729–1734.
[9]Martinez,S.,Barbas,C.,Herrera,E. Uptake of alpha-tocopherol by the mammary gland but not
by white adipose tissue is dependent on lipoprotein lipase activity around parturition and during lactation in the rat[J].Metabolism ,2002,51,1444–1451.
[10]Bjorneboe,A.,Bjorneboe,G.E.,Drevon,C.A. Serum half-life, distribution, hepatic uptake and
biliary excretion of alpha-tocopherol in rats[J].Biochem. Biophys, 1987,921,175–181.
[11]郑景洲,许友卿.维生素E的功能、吸收与代谢(2)[J].生物学通报.2005,40(12):1-2.
[12]Reboul,E.,Klein,A.,Bietrix,F.,Gleize,B.,et al. Scavenger receptor class B type I(SR-BI)is involved
in vitamin E transport across the enterocyte[J].Biol.Chem,2006,281,4739–4745.
[13]Anwar,K.,Kayden,H.J.,Hussain,M.M. Transport of vitamin E by differentiated Caco-2 cells[J].
Lipid Res,2006,47,1261–1273.
[14]Saito,K.;Tautz,L.;Mustelin, T. The lipid-binding SEC14 domain[J]. Biochem.Biophys,2007 ,1771:
719–726.
[15]Smion E J,E isengart A,Sundhemi L,et al. The metabolism of vitamin E.ⅡPurification and
characterization of urinary metabolites of tocopherol[J].J BiolChem,1956,221:807.
[16]李军生. 维生素E吸收与代谢机制的研究进展[J]. 中国现代应用药学杂志,2005,22(6):
457-461.
[17]董晓慧,杨原志. 自由基与维生素E的抗氧化作用[J]. 饲料研究,2003,6:15-18.
[18]刘佃辛.维生素E在生物膜中的作用[J].生理科学进展,1993,124(1):83~85
[19]王改琴,李维,王恬. 维生素E对家禽抗氧化、免疫功能和繁殖性能的影响研究进展[J]. 家
畜生态学报,2009,30(6):1-5
[20]Sahoo D K,Roy A,Chainy G B N. Protective effects of vitamin E and curcumin on
L-thyroxine-induced rat testicular oxidative stress[J].Chemico-Biological Interactions,2008, 176: 121-128.
[21]Brigelius-FlohéR. Vitamin E: The shrew waiting to be tamed[J].Free Radical Biology and
Medicine,2009,46:543-554.
[22]刘树立,盛占武, 王华. 饲用维生素E的研究进展[J].新饲料,2007,(6):37-40.
[23]朱虹,罗海玲,孟慧. 维生素E对动物繁殖性能的影响及其机理研究进展[A]. 2007-2008年
全国养羊生产与学术研讨会议论文集,2008,65-66.
[24]Moran E T Jr. Nutrition of the developing embryo and hatchling[J].Poultry Science,2007,86:
1043-1049.
[25]王丽,侯先志,王海荣等. 不同维生素E水平对围产期奶牛血液免疫指标的影响[J].饲料工
业,2010,31(15):30-33.
[26]Freedman J E,Farhat J H,Loscalzo J,et al. Alpha-tocopherol inhibits aggregation o f human
platelets by a protein kinase C dependent mechanism[J].Circulation,1996,94:2434 -2440. [27]Chan A C,Wagner M,Kennedy C,et al. Vitamin E up regulates phospho lipase A2,arachidonic
acid release and cyclooxygenase in endothelia l cells[J].Aktuel Emahrungsmed,1998,23:1-8.
[28]Villacorta,L.,Graca-Souza,A.V.,Ricciarelli,R.,Zingg,J.M.,Azzi,A.,a-Tocopherol induces
expression of connective tissue growth factor and antagonizes tumor necrosis factor mediated downregulation in human smooth muscle cells[J].Circ.Res,2003,92:104–110. [29]Aratri,E,Spycher,S.E,Breyer,I.,Azzi,A., Modulation of a-tropomyosin expression by
a-tocopherol in rat vascular smooth muscle cells.[J]FEBS Lett,1999,447:91–94.
[30]Yamauchi,J.,Iwamoto,T.,Kida,S.,Masushige,S.,et al., Tocopherol-associated protein is a
ligand-dependent transcriptional activator[J]. Biochem.Biophys.
https://www.doczj.com/doc/d21390241.html,mun,2001,285:295–299.
[31]呙于明,陈继兰.0-3周龄肉仔鸡日粮中维生素E的适宜供给量研究[J].畜牧兽医学
报,1999,(4):289-295
[32]Rebole A,Rodriguez M L,Ortiz L T,et al. Effect of dietary high oleic acid sunflower seed,
palm,oil and vitamin E supplementation on broiler performance,fatty acid composition and oxidation susceptibility of meat[J].British Poultry Science,2006,47:581-591.
[33]李绍钰,魏凤仙,郝国庆等.维生素E的对肉鸡生产性能及肉质的影响[J].河南农业科
学,2001,6:43-45.
[34]李新花,王晓霞,禇耀诚.日粮维生素E、C的添加水平对AA肉仔鸡生产性能及血清抗氧化
酶活力的影响[J].北京农学院学报,2006,21(1):33~37.
[35]李彦,杨在宾,杨维仁等.日粮维生素A和维生素E水平对肉鸡抗氧化和免疫性能的影响[J].
动物营养学报2008,20(4):417 422.
[36]Lin YF,Chang SJ,Hsu AL. Effects of supplemental vitamin E during the laying period on the
reproductive performance of Taiwan native chickens[J].Br Poult Sci,2004,45(6):807-814. [37]田河,石君,宁志利.不同剂量的维生素E对产蛋后期蛋种鸡繁殖性能的影响.新饲
料,2009:36-38
[38]马兆臣.维生素E和硒对皖西白鹅繁殖性能和生殖免疫的影响[D].安徽:安徽农业大学.硕
士学位论文;2007.
[39]Hennig A,Marckwardt E,Richter G. Relation between vitamin E supply and the fertility of
laying hens[J].Arch Tierernahr,1986,36(6):519-29.
[40]赵国先,张振红. 不同日粮条件下添加维生素E对蛋种鸡繁殖性能的影响[J].中国饲料,
2011,3:23-25.
[41]陈岩锋,陈晖,郑丽祯.日粮中添加维生素E对种鸭繁殖性能的影响(初报)[J].福建畜牧兽
医,2001,23(2):5.
[42]许万祥.维生素对禽胚胎发育和孵化率的影响[J].中国饲料,1997,(24):7-9.
[43]Lin YF,Chang SJ,Yang JR,Lee YP,Hsu AL. Effects of supplemental vitamin E during the mature
period on the reproduction performance of Taiwan Native Chicken cockerels[J].Br Poult Sci.
2005,46(3):366-373.
[44]Biswas A,Mohan J,Sastry KV. Effect of higher dietary vitamin E concentrations on physical
and biochemical characteristics of semen in Kadaknath cockerels[J].Br Poult
Sci.2009,50(6):733-738.
[45]陈鑫.低温和维生素E对笼养育成蛋鸭生长及生化指标的影响[D].硕士学位论文.哈尔滨:
东北农业大学.2007.
[46]Ruiz J A,Perez-Vendrell A M,Esteve-Garcia E. Effect of beta-carotene and vitamin Eon
oxidative stability in leg meat of broilers fed different supplemental fats[J].Journal of Agriculture and Food Chemistry,1999,47(2):448-454.
[47]Jensen C,Skibsted L H,Jakobsen K,et al. Supplementation of broiler diets with all-rac-alpha-or
a mixture of natural source RRR-alpha,ganlnla,delta-tocopheryl acetate. Effect on the
oxidative stability of raw and precooked broiler meat products[J].Poultry Science,1995,74(12):2048-2056.
[48]Sarraga C,Garcla Regueiro J A. Membrane lipid oxidation and proteolytic activity in thigh
muscles from broilers fed different diets[J].Meat Science,1999,52(2):213-219.
[49]候永清,李绍钰.饲料中添加维生素E对畜禽肉质的影响[J].饲料博览,2001,(9):30-32.
[50]李同树,韩瑞丽,邢永国,等.日粮维生素E水平对鲁西黄鸡产肉性能与肉质的影响[J].动物
营养学报,2003,15(4):44-48.
[51]Nacim Zouari,Fatma Elgharbi,Nahed Fakhfakh,et,al. Effect of dietary vitamin E
supplementation on lipid and colour stability of chicken thigh meat[J]. African Journal of Biotechnology.2010,15(9):2276-2283.
[52]Guo Yuming,Tang Qing,Yuan Jianmin,et al. Effects of supplementation with vitamin E on the
performance and the tissue peroxidation of broiler chicks and the stability of thigh meat against oxidative deterioration[J].Animal Feed Science and Technology,2001,89:165-173. [53]Bartov I,Frigg M. Effect of high concentrations of dietary vitamin E during various age
periods on performance, plasma vitamin and meat stability of broiler chicks at 7 weeks of age[J].British Poultry Science,1992,33(2):393-402.
[54]周林.生物素和维生素对肉鸡屠宰性能和肉品质的影响[J].饲料博览,2000,(8):6-7.
[55]高海燕,张丽明. VE对鸡肉品质的影响[J].兽药与饲料添加,2008,13(3):11-13.
[56]C. Narciso-Gaytán, D. Shin, A. R,et.al. Dietary lipid source and vitamin E effect on lipid
oxidation stability of refrigerated fresh and cooked chicken meat[J]. Poult Sci,2010,89:2726-2734.
[57]Schiavone,A.,Marzoni,M.,Castillo,A.,Nery,J.and Romboli,I. Dietary lipid sources and vitamin E
affect fatty acid composition or lipid stability of breast meat from Muscovy duck[J].Can.J.Anim.Sci. 2010,90:371-378.
[58]刘铀,刘艳芬.维生素E对肉鸡免疫功能的增强作用[J].中国兽医杂志,1999,(6):11-13.
[59]文杰,林济华.日粮维生素E、抗坏血酸水平对肉仔鸡生长及免疫功能的影响[J].畜牧兽医
学报,1996,27(8):481-488.
[60]吴江利,王安,张养东.维生素E对育成金定蛋鸭生长及免疫和抗氧化指标的影响[J].动物
营养学报,2008,20(6):686-691.
[61]Gore A B,Qureshi M A. Enhancement of humoral and cellular immunity by vitamin E after
embryonic exposure[J].Poultry Science,1997,76:984-991.
[62]Erf G F,Bot t jc W G,Bers i T K,et al. Effects of dietary vitamin E on the immune system in
broilers: altered proportions of CD4+ T cells in the thymus and splecn[J].Poultry Science, 1998, 77:529-537.
[63]Combs G F J,N oguchi T,Scot t M L. Mechanism of action of selenium and vitamin E in
protection of biological membances[J].Federation proceedings,1975,34(11):2090-2095.
[64]Gerrit J V,Lemm ens A G,A nt on C B. Copper status in rats fed diets supplemented with
either vitamin E, vitamin A, or carotene[J].Biology Trace Element Research,1993,37:253-259.
[65]刘乾,李云.维生素E对畜禽免疫功能的影响[J].饲料研究,2002,(1):22-23.
[66]傅德顺.现代蛋鸡的饲养管理(下)[J].禽畜业,1999,111(7):36-39.
[67]王晓霞,滑静等. VC及VE对热应激蛋鸡生产性能及抗氧化功能的影响[J].北京农学院学
报,2000,15(4):37-38.
[68]R.Vakili,Ali Asghar Rashidi ,S.Sobhanirad. Effects of dietary fat, vitamin E and zinc
supplementation on tibia breaking strength in female broilers under heat stress.[J] African Journal of Agricultural Research,2010,5(23): 3151-3156.
[69]刘轴,刘艳芬,林红英,罗东君. 维生素E防治肉鸡热应激机理的初步研究[J].中国家
禽,1999, 21(6):6-9.
[70]方洛云,邹晓庭,许梓荣. 维生素E对家禽热应激和肉质的影响[J].饲料工业, 2000,21(2):
39-40.
[71]孙长勉,栾新红,曹中赞,郑雅文,胡建民. 维生素E对热应激条件下肉鸡甲状腺激素分泌的
影响[J]. 沈阳农业大学学报,2002,33(5):360-362.
[72]Shen Y. On the requirement of Vitamin in fast and slow growing chickens: experiments with
broiler and Leghorn-type chickens[J].J Anim Phydiol Anim Nutr,1992,67(3):113-122.
[73]王爽,侯水生等. 生长前期北京鸭维生素E需要量的研究[J].中国饲料.2010,8:25-26.
[74]张小纯,丁雪梅,张晶等. 不同维生素E水平对骡鸭产肝性能及血液生化指标的影响[J].
饲料博览.2010,6:1-4.