
Phytoremediation and hyperaccumulator plants Wendy Ann Peer, Ivan R. Baxter, Elizabeth L. Richards, John L. Freeman, Angus S. Murphy
Abstract
Phytoremediation is a group of technologies that use plants to reduce, remove, de-grade, or immobilize environmental toxins, primarily those of anthropogenic ori-gin, with the aim of restoring area sites to a condition useable for private or public applications. Phytoremediation efforts have largely focused on the use of plants to accelerate degradation of organic contaminants, usually in concert with root rhizosphere microorganisms, or remove hazardous heavy metals from soils or wa-ter. Phytoremediation of contaminated sites is a relatively inexpensive and aes-thetically pleasing to the public compared to alternate remediation strategies in-volving excavation/removal or chemical in situ stabilization/conversion. Many phytoremediation plans have multi-year timetables, but since most sites in need of remediatrion have been contaminated for more than ten years, as such a ten year remediation plan does not seem excessive. Seven aspects of phytoremediation are described in this chapter: phytoextraction, phytodegradation, rhizosphere degrada-tion, rhizofiltration, phytostabilization, phytovolatization, and phytorestoration. Combining technologies offer the greatest potential to efficiently phytoremediate contaminated sites. The major focus of this chapter is phytoextraction of arsenic, cadmium, chromium, copper, mercury, nickel, lead, selenium, and zinc.
1 Introduction to phytoremediation
Phytoremediation is a term applied to a group of technologies that use plants to reduce, remove, degrade, or immobilize environmental toxins, primarily those of anthropogenic origin, with the aim of restoring area sites to a condition useable for private or public applications. To date, phytoremediation efforts have focused on the use of plants to accelerate degradation of organic contaminants, usually in concert with root rhizosphere microorganisms, or remove hazardous heavy metals from soils or water. Phytoremediation of contaminated sites is appealing because it is relatively inexpensive and aesthetically pleasing to the public compared to al-ternate remediation strategies involving excavation/removal or chemical in situ stabilization/conversion. Seven aspects of phytoremediation are described in this chapter: phytoextraction, phytodegradation, rhizosphere degradation, rhizofiltra-tion, phytostabilization, phytovolatization, and phytorestoration. However, the ma-jor focus is on phytoextraction.
Topics in Current Genetics, Vol. 14
M.J. Tamás, E. Martinoia (Eds.): Molecular Biology of Metal Homeostasis
and Detoxification
DOI 10.1007/4735_100 / Published online: 11 August 2005
? Springer-Verlag Berlin Heidelberg 2005
300 Wendy Ann Peer et al.
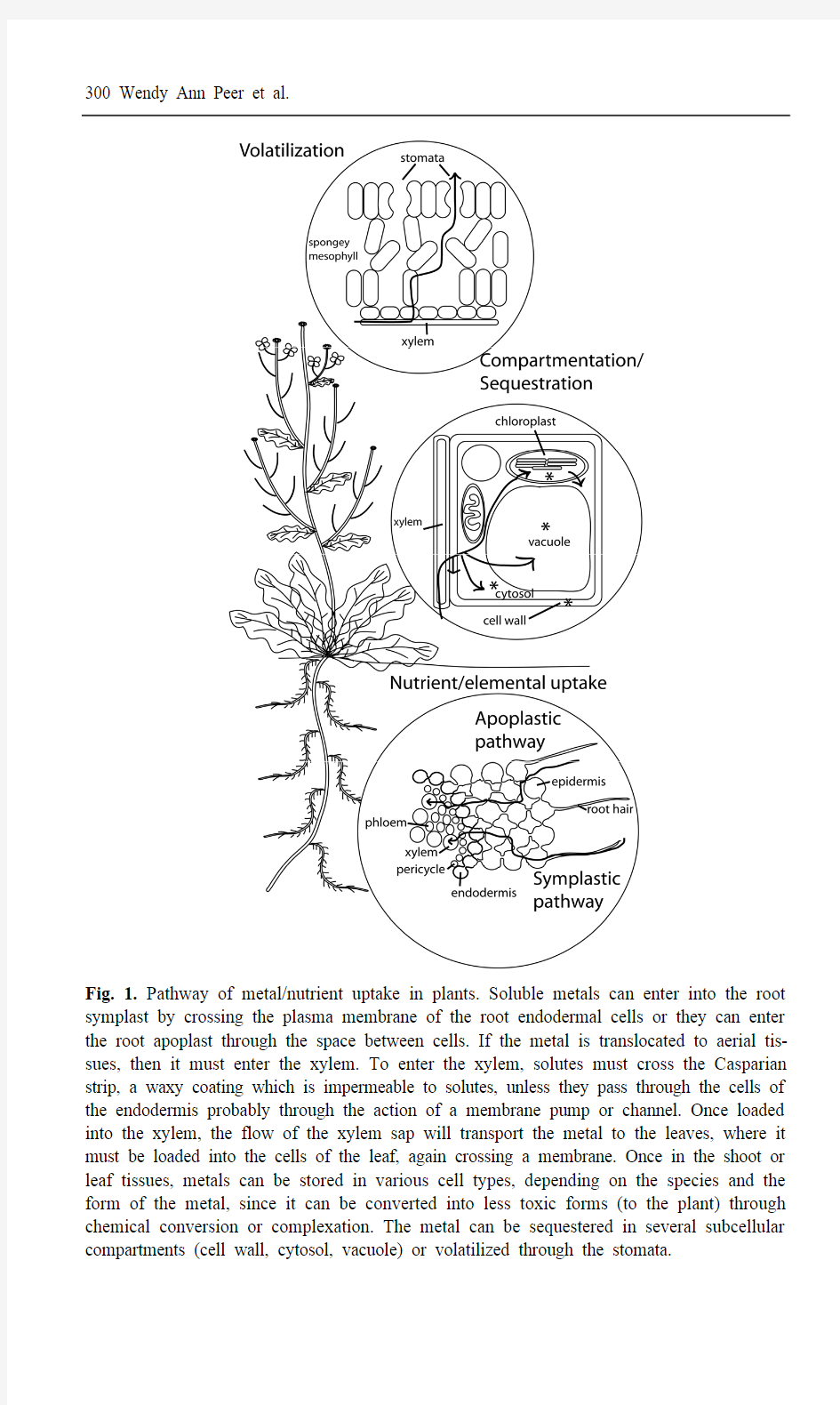
Fig. 1. Pathway of metal/nutrient uptake in plants. Soluble metals can enter into the root symplast by crossing the plasma membrane of the root endodermal cells or they can enter the root apoplast through the space between cells. If the metal is translocated to aerial tis-sues, then it must enter the xylem. To enter the xylem, solutes must cross the Casparian strip, a waxy coating which is impermeable to solutes, unless they pass through the cells of the endodermis probably through the action of a membrane pump or channel. Once loaded into the xylem, the flow of the xylem sap will transport the metal to the leaves, where it must be loaded into the cells of the leaf, again crossing a membrane. Once in the shoot or leaf tissues, metals can be stored in various cell types, depending on the species and the form of the metal, since it can be converted into less toxic forms (to the plant) through chemical conversion or complexation. The metal can be sequestered in several subcellular compartments (cell wall, cytosol, vacuole) or volatilized through the stomata.
Phytoremediation and hyperaccumulator plants 301 1.1 Phytoextraction
Phytoextraction involves the removal of toxins, especially heavy metals and met-alloids, by the roots of the plants with subsequent transport to aerial plant organs (Salt et al. 1998; Lombi et al. 2001a) (Fig. 1). Pollutants accumulated in stems and leaves are harvested with accumulating plants and removed from the site. Phyto-extraction can be divided into two categories: continuous and induced (Salt et al. 1998). Continuous phytoextraction requires the use of plants that accumulate par-ticularly high levels of the toxic contaminants throughout their lifetime (hyperac-cumulators), while induced phytoextraction approaches enhance toxin accumula-tion at a single time point by addition of accelerants or chelators to the soil. In the case of heavy metals, chelators like EDTA assist in mobilization and subsequent accumulation of soil contaminants such as lead (Pb), cadmium (Cd), chromium (Cr), copper (Cu), nickel (Ni), and zinc (Zn) in Brassica juncea (Indian mustard) and Helianthus anuus (sunflower) (Blaylock et al. 1997; Turgut et al. 2004). The ability of other metal chelators such as CDTA, DTPA, EGTA, EDDHA, and NTA to enhance metal accumulation has also been assessed in various plant species (Huang et al. 1997; Lombi et al. 2001b). However, there may be risks associated with using certain chelators considering the high water solubility of some chela-tor-toxin complexes which could result in movement of the complexes to deeper soil layers (Wu et al. 1999; Lombi et al. 2001b) and potential ground water and es-tuarian contamination.
1.2 Phytodegradation
In phytodegredation, organic pollutants are converted by internal or secreted en-zymes into compounds with reduced toxicity (Schnoor 1997; Salt et al. 1998; Suresh and Ravishankar 2004). For instance, the major water and soil contaminant trichloroethylene (TCE) was found to be taken up by hybrid poplar trees, Populus deltoides x nigra, which break down the contaminant into its metabolic compo-nents (Newman et al. 1997). TCE and other chlorinated solvents can be degraded to form carbon dioxide, chloride ion and water (Schnoor et al. 1995). Poplars have also been shown to take up the ammunition wastes 2,4,6-trinitrotoluene (TNT), hexahydro-1,3,5-trinitro-1,3,5 triazine (RDX), octahydro-1,3,5,7-tetranitro-1,3,5,7 tetrazocine (HMX) and partially transform them (Thompson et al. 1998; Yoon et al. 2002). Root exudates from Datura innoxia and Lycopersicon peruvianum con-taining peroxidase, laccase, and nitrilase have been shown to degrade soil pollut-ants (Schnoor et al. 1995; Lucero et al. 1999) and nitroreductase and laccase to-gether can break down TNT, RDX, and HMX (Schnoor et al. 1995). The plants are then able to incorporate the broken ring structures into new plant material or organic soil components that are thought to be non-hazardous.
302 Wendy Ann Peer et al.
1.3 Phytovolatilization
Plants can also remove toxic substances, such as organics, from the soil through phytovolatization. In this process, the soluble contaminants are taken up with wa-ter by the roots, transported to the leaves, and volatized into the atmosphere through the stomata (Tollsten and Muller 1996; Newman et al. 1997; Davis 1998) (Fig. 1). The best example of this is the volatilization of mercury (Hg) by conver-sion to the elemental form in transgenic Arabidopsis and yellow poplars contain-ing bacterial mercuric reductase (merA) (Rugh et al. 1996, 1998) (see Section 3.5). In a study where the movement of volatile organics was monitored by Fourier transform infrared spectrometry (FT-IR) in hybrid poplars (Populus deltoides x nigra), Tamarix parviflora (saltcedar), and Medicago sativa (alfalfa), chlorinated hydrocarbons were found to move readily through the plants, but less polar com-pounds like gasoline constituents did not (Davis et al. 1998). However, amounts of the contaminant transpired are in proportion to water flow and are relatively low, especially in the field. Rubin and Ramaswami (2001) found that poplar saplings can concentrate (100 ppb) and transpire methyl tertiary-butyl ether (MTBE), a compound added to gasoline which is commonly found as a groundwater pollut-ant. In a one week time period, they observed a 30% reduction in MTBE mass in hydroponic solution by saplings at both high (1600 ppb) and low (300 ppb) MTBE concentrations, which suggested that these plants could be successful in the phy-toremediation of this toxin from groundwater (Rubin and Ramaswami 2001). Se-lenium (Se) is a special case of a metal that is taken up by plants and volatilized (see Section 3.8). Se can also be volatilized following conversion to dimethylse-lenide by microbes and algae (Neumann et al. 2003) (see Chapter 13).
1.4 Rhizosphere degradation
Like phytodegradation, rhizosphere degradation involves the enzymatic break-down of organic pollutants, but through microbial enzymatic activity. These breakdown products are either volatilized or incorporated into the microorganisms and soil matrix of the rhizosphere. The types of plants growing in the contami-nated area influence the amount, diversity, and activity of microbial populations (Jones et al. 2004; Kirk et al. 2005). Grasses with high root density, legumes that fix nitrogen, and alfalfa that fix nitrogen and have high evapotranspiration rates are associated with different microbial populations. These plants create a more aerobic environment in the soil that stimulates microbial activity that enhances oxidation of organic chemical residues (Anderson 1993; Schnoor et al. 1995; Narayanan 1998; Jones et al. 2004; Kirk et al. 2005). Secondary metabolites and other components of root exudates also stimulate microbial activity, a byproduct of which may be degradation of organic pollutants (Pieper et al. 2004).
Phytoremediation and hyperaccumulator plants 303 1.5 Rhizofiltration
Rhizofiltration removes contaminants from water and aqueous waste streams, such as agricultural run off, industrial discharges, and nuclear material processing wastes (Salt et al. 1998; Suresh and Ravishankar 2004). Absorption and adsorp-tion by plant roots play a key role in this technique, and consequently large root surface areas are usually required. In research associated with Epcot Center, closed systems with recirculating nutrients have exhibited the benefits of rhizofil-tration and biofiltration using a variety of species (such as mosses and scented ge-raniums) (Negri and Hinchman 1996). Rhizofiltration was also shown to be useful in the San Francisco Bay study directed by Norman Terry (University of Califor-nia, Berkely) and supported by Chevron (Hansen et al. 1998). A wetland con-structed next to the bay was shown to remove 89% of the Se from selenite-contaminated wastewater released from various oil refineries. The water flowing into the wetland was measured to have 20–30 μg L?1selenite, while the outflow from the wetland had less than 5 μg L?1 selenite (Hansen et al. 1998). In a study of Se removal from agricultural subsoil drainage in the San Joaquin Valley (Gao et al. 2003), a flow-through wetland system was constructed with cells containing ei-ther a single species, or a combination of species [e.g. Schoenoplectus robustus (sturdy bulrush), Juncus balticus (baltic rush), Spartina alterniflora (smooth cordgrass), Polypogon monspeliensis (rabbit’s foot grass), Distichlis spicata (salt-grass), Typha latifolia (cattail), Schoenoplectus acutus (Tule grass), and Ruppia maritima (widgeon grass)]. Four years after planting, comprehensive analysis showed that 59% of the Se remained in the wetland, mostly in the organic detrital layer and surface sediment, 35% in the outflow, 4% in seepage and 2% to volatili-zation. Wetland plant uptake of Se varies with species type, and parrot’s feather (Myriophyllum aquaticum), iris-leaved rush (Juncus xiphioides), cattail, and sturdy bulrush were particularly noted for high Se uptake potential (Gao et al. 2003).
1.6 Phytostabilization
Erosion and leaching can mobilize soil contaminants resulting in aerial or water-borne pollution of additional sites. In phytostabilization, accumulation by plant roots or precipitation in the soil by root exudates immobilizes and reduces the availability of soil contaminants. Plants growing on polluted sites also stabilize the soil and can serve as a groundcover thereby reducing wind and water erosion and direct contact of the contaminants with animals. Significant phytostabilization pro-jects have been employed in France and the Netherlands (Ernst et al. 1996; Bouwman et al. 2001; Marseille et al. 2000). A 2005-2010 superfund basic re-search program (Maier 2004) is developing a phytostabilization revegetation strat-egy to remediate mine tailings in arid and semi-arid ecosystems. The researchers will monitor the bioavailability of metals for the native metal- and drought-tolerant plant species used, and determine the permanence of expected toxicity re-ductions.Plants with high transpiration rates, such as grasses, sedges, forage
304 Wendy Ann Peer et al.
Table 1. Summary of elemental levels in background soil, metalliferous (or contaminated) soil, critical load in soil above which biodiversity and ecosystem function are adversely af-fected, and Commission des Communautés Européennes (EU) and Environmental Protec-tion Agency (USA) permissible limits.
Element Background soil levels (ppm)a
Metalliferous soil levels (ppm)c
Critical load in loam/silt (ppm) CCE limits (ppm)h EPA lim-its k,i As 2.2 – 25
1 510 – – 0.01 ppm Cd 0.06 – 1.1
317 1.10 0.5i 5 ppb Cr 7 – 221
3 450d 64.41 1.5i 100 ppb Cu 6 – 80
3 783 48.78 50j 1.3 ppm Hg 0.02 – 0.41
12 000e 0.56 1.5 2 ppb Ni 4 – 55
11 260 54.64 1i 0.7 ppm Pb 10 – 84
49 910 75.68 5i 1.5 ppb Se 0.01 – 0.09b
>50f – – 0.05 ppm Zn 17 – 125
7 480 207.32 150j 5 ppm a https://www.doczj.com/doc/7e16276826.html, b Lakin 1972 c Reeves and Baker 2000 d https://www.doczj.com/doc/7e16276826.html, e https://www.doczj.com/doc/7e16276826.html, f https://www.doczj.com/doc/7e16276826.html, g Bannick et al. 2002; Sand ~50% of loam/silt levels; clay ~1.5X loam/silt levels h http://europa.eu.int i H 2O j Soil k https://www.doczj.com/doc/7e16276826.html,
plants, and reeds are useful for phytostabilization by decreasing the amount of ground water migrating away from the site carrying contaminants (Suresh and Ravishankar 2004). Combining these plants with hardy, perennial, dense rooted or deep rooting trees (poplar, cottonwoods) can be an effective combination (Berti and Cuningham 2000).
1.7 Phytorestoration
Phytorestoration involves the complete remediation of contaminated soils to fully functioning soils (Bradshaw 1997). In particular, this subdivision of phytoreme-diation uses plants that are native to the particular area, in an attempt to return the land to its natural state. An examination of phytorestoration compared to the other forms of phytoremediation brings to light an important issue: what degree of de-contamination do phytoremediation projects aim to achieve? There is a vast dif-ference between removing just enough soil pollutants to reach legally defined lev-els of compliance, remediating soils to a level at which they can be used again, and completely restoring land from its contaminated state to an environmentally
Phytoremediation and hyperaccumulator plants 305 uncontaminated state (Table 1). The objective of many phytoremediation projects is to restore the land to a legally acceptable level of contamination.
Lastly, a combination of phytoremediation approaches can be used for more ef-fective environmental restoration. This may help to simultaneously remove differ-ent types of wastes from the same site. For example, a remediation system could include plants that hyperaccumulate toxic metals and plants that stimulate the ac-tivity of microbes that specialize in organic contaminant degradation.
2 Definitions of tolerant, indicator, and hyperaccumulator species
When categorizing plants that can grow in the presence of toxic elements, the terms “tolerant,” “indicator”, and “hyperaccumulator” are used. A tolerant species is one that can grow on soil with concentrations of a particular element that are toxic to most other plants. While both indicator species and hyperaccumulators are also tolerant, studies have shown the genetic distinction of the mechanisms in-volved (Assun??o et al. 2001; Bert et al. 2003;Macnair et al. 1999). However, tol-erant species are not necessarily indicators or hyperaccumulators, as tolerant non-accumulators can exclude metals from entering the root tissue. Examples of toler-ant excluders include Holcus lanatus (Meharg and Macnair 1992a, 1992b), Agrostis capillaris, Mimulus guttatus, and Silene vulgaris (Pollard et al. 2002).
Indicator plants have been employed in biogeochemical prospecting. As early as 1865, F. Risse observed Zn accumulating plants, now known as Thlaspi caerulescens, growing near the German-Belgium border (Sachs 1865). This ob-servation led others to associate the sites where these plants grew with soil con-taining elevated Zn. In the 1950s and 1960s, Helen Cannon and other members of the United Sates Geological Society cataloged indicator plants that were poten-tially important for “bioprospecting” for ore (Cannon 1960). Examples include mosses, which have been recognized as bioindicators of high metal concentrations in water (Gstoettner and Fisher 1995) and Stanleya pinnata (Prince’s Plume), which is a recognized Se indicator.
Hyperaccumulators take up particularly high amounts of a toxic substance, usu-ally a metal or metalloid, in their shoots during normal growth and reproduction (Reeves 1992; Baker and Whiting 2002). Hyperaccumulation reported in senesc-ing plants generally represents a breakdown of homeostatic mechanisms and is clearly not a function of normal growth processes, although such accumulations could be technologically useful. The metal/metalloid concentration that must be accumulated by the plant before it is designated a “hyperaccumulator” depends upon the particular metal or metalloid in question. In early hyperaccumulator stud-ies, Brooks and coworkers (1977) defined nickel (Ni) hyperaccumulators as those accumulating greater than 1000 μg Ni g-1 dry weight in their leaves. Subsequently, Baker and Brooks (1989) defined threshold concentrations for other metals hyper-accumulated in plants as 100 μg g-1 dry weight for Cd, 1,000 μg g-1 dry weight for Ni, Cu, Co, Pb, and 10,000 μg g-1 dry weight for Zn and Mn. The defined levels of
306 Wendy Ann Peer et al.
these elements are typically at a concentration of one order of magnitude greater that those found in non-accumulator species (Salt and Kramer 2000). Hyperaccu-mulators are found in 45 different families, with the highest occurrence among the Brassicaceae (Reeves and Baker 2000). These plants are quite varied, from peren-nial shrubs and trees to small annual herbs.
While tolerance is necessary for accumulation, evidence suggests that tolerance and accumulation are independent traits. There is a strong positive correlation be-tween glutathione (GSH), cysteine (Cys), O-acetyl-L-serine (OAS) levels, com-pounds associated with tolerance, and Ni accumulation in shoot tissues of Thlaspi species from serpentine soils (Freeman et al. 2004). Thlaspi goesingense has con-stitutively high levels of serine acetyltransferase (SAT) and glutathione reductase activity associated with resistance to Ni-induced oxidative stress in this plant (Freeman et al. 2004). Elevated levels of reduced GSH contribute to a decrease in Ni-induced lipid peroxidation in T. goesingense shoots and lower levels of Ni-induced reactive oxygen species in roots. Salicylic acid (SA) was observed to both regulate glutathione accumulation through post-translational activation of SAT, and to increase glutathione reductase activity in A. thaliana (Freeman et al. 2005). Changes in the levels of these compounds and enzymes appear to be associated with Ni tolerance required for hyperaccumulation (For additional details about Ni tolerance in plants, see Chapter 9; for additional details about heavy metal chela-tion, see Chapter 10).
Overexpression of T. goesingense mitrochondrial SAT (TgSAT-m) in A. thaliana was found to confer increased Ni tolerance compared to controls (Free-man et al. 2004). These overexpressors also showed increased OAS, Cys and GSH biosynthesis (Freeman et al. 2004). However, A. thaliana SAT overexpressors and controls had the same shoot Ni content indicating that the SAT overexpressors neither hyperaccumulated nor excluded Ni, and chemical speciation showed that the Ni was not bound to thiols in these plants (Freeman et al. 2004). It is notewor-thy that this demonstration of Ni tolerance without accumulation represents an ex-ample of the distinction between metal tolerance mechanisms and metal hyperac-cumulation mechanisms and supports previous findings showing partial independent genetic control of hyperaccumulation and tolerance (Assun??o et al. 2001; Bert et al. 2003; Macnair et al. 1999).
2.1 How do plants take up and transport metal?
The process of metal accumulation involves several steps, outlined in Fig. 1, one or more of which are enhanced in hyperaccumulators.
2.1.1 Solubilization of the metal from the soil matrix
Many metals are found in soil-insoluble forms. Plants use two methods to desorb metals from the soil matrix: acidification of the rhizosphere through the action of plasma membrane proton pumps and secretion of ligands capable of chelating the metal. Plants have evolved these processes to liberate essential metals from the
Phytoremediation and hyperaccumulator plants 307 soil, but soils with high concentrations of toxic metals will release both essential and toxic metals to solution. To our knowledge, there are no reports of plants with the ability to solublize Pb from the soil matrix, where most of soil Pb exists in an insoluble form (Blaylock and Huang 2000). Experiments demonstrating Pb hyper-accumulation have used Pb(NO3)2, a soluble form of Pb, though it must be ques-tioned whether this is the most appropriate form of Pb for analysis. Aside from Pb, the solubilization mechanisms for hyperaccumulators are similar for metals dis-cussed, and therefore will not be addressed independently for each metal. While no hyperaccumulators have evolved to handle high concentrations of toxic metals if they are present in solution, phytoremediator plants could be modified to solubi-lize contaminants that are bound to the soil.
2.1.2 Uptake into the root
Soluble metals can enter into the root symplast by crossing the plasma membrane of the root endodermal cells or they can enter the root apoplast through the space between cells (Fig. 1). While it is possible for solutes to travel up through the plant by apoplastic flow, the more efficient method of moving up the plant is through the vasculature of the plant, called the xylem. To enter the xylem, solutes must cross the Casparian strip, a waxy coating, which is impermeable to solutes, unless they pass through the cells of the endodermis (Fig. 1). Therefore, to enter the xylem, metals must cross a membrane, probably through the action of a mem-brane pump or channel. Most toxic metals are thought to cross these membranes through pumps and channels intended to transport essential elements. Excluder plants survive by enhancing specificity for the essential element or pumping the toxic metal back out of the plant (Hall 2002; Meharg and Macnair 1992a, 1992b).
2.1.3 Transport to the leaves
Once loaded into the xylem, the flow of the xylem sap will transport the metal to the leaves, where it must be loaded into the cells of the leaf, again crossing a membrane (Fig. 1). The cell types where the metals are deposited vary between hyperaccumulator species. For example, T. caerulescens was found to have more Zn in its epidermis than in its mesophyll (Kupper et al. 1999), while A. halleri preferentially accumulates its Zn in its mesophyll cells instead of its epidermal cells (Kupper et al. 2000).
2.1.4 Detoxification/Chelation
At any point along the pathway, the metal could be converted to a less toxic form through chemical conversion or by complexation. Various oxidation states of toxic elements have very different uptake, transport, sequestration or toxicity character-istics in plants. Chelation of toxins by endogenous plant compounds can have similar effects on all of these properties as well. As many chelators use thiol groups as ligands, the sulfur (S) biosynthetic pathways have been shown to be
308 Wendy Ann Peer et al.
Fig. 2. Structures of some compounds that can bind metals: amino acids, glutathione, and phytochelatin.
Table 2. Summary of elements and the element-organic complexes that may be formed within plants.
ligand
Element Analogue Organic
thiol, glutathione, ADP-As, ascorbic acid Arsenic Phosphate
Phytochelatin,
Cadmium Zn, Fe Phytochelatin, glutathione, γ-glutamylcysteine, thiols Chromium Mn Thiols
metallothioniens, phytochelatin 2, phytochelatin 3 Copper Cu Citrate,
Mercury Unknown a Thiols
Nickel Fe Nicotianamine, histidine, thiols, citrate
Fe Glutathione
Lead Zn,
Selenium S Cystiene,
methionine, with and without methylation
γ-glutamylcysteine, thiols, cit-
glutathione,
Zinc Zn Phytochelatin,
rate, malate
a Enters cell through passive diffusion.
Phytoremediation and hyperaccumulator plants 309 critical for hyperaccumulator function (Ng and Anderson 1979; Pickering et al. 2003; Van Huysen et al. 2004) and for possible phytoremediation strategies (Figs.
2 and 3; Table 2). Oxidative stress is one of the most common effects of heavy metal accumulation in plants, and the increased anti-oxidant capabilities of hyper-accumulators allow tolerance of higher concentrations of metals (Freeman et al. 2004).
2.1.5 Sequestration/Volatilization
The final step for the accumulation of most metals is the sequestration of the metal away from any cellular processes it might disrupt. Sequestration usually occurs in the plant vacuole, where the metal/metal-ligand must be transported across the vacuolar membrane. Metals may also remain in the cell wall instead of crossing the plasma membrane into the cell, as the negative charge sites on the cell walls may interact with polyvalent cations (Wang and Evangelou 1994). Selenium may also be volatized through the stomata.
2.2 Strategies for phytoremediation using hyperaccumulators
The effectiveness of a phytoremediation plan is dependent on the selection of the appropriate plant or plants. Plants native to the target area should be considered since they are adapted to the local climate, insects, and diseases. Any plant used as a phytoremediator must be able to tolerate high concentrations of the toxic sub-stance of interest, in addition to any other pollutants found at the particular site, as candidate sites for phytoremediation usually have multiple contaminants. In the United States, more than 80% of the metal contaminated Superfund, Department of Defense, and Department of Energy sites are also contaminated by organic pol-lutants (Ensley 2000).
Plants used for phytoextraction should develop a large amount of biomass quickly and be easy to cultivate and harvest, preferably multiple times per year (Newman 1997; Tong et al. 2004). For example, Mcgrath and Zhao (2003) have calculated that a plant with a bioconcentration factor of 40 can halve the concen-tration of metal in the top 20 cm of soil in 10 crops if it produces 5 tonnes ha-1 crop-1, but a plant with a bioconcentration factor of 20 must produce at least 10 tonnes ha-1 crop-1 to have the same effect. Since most metal hyperaccumualtors are small plants with low biomass, efforts are being made to locate new hyperaccumu-lators, selectively breed for promising plant traits, and create transgenic phytore-mediators. For example, hyperaccumulation of arsenic in the fern Pteris vittata was recently discovered (Ma et al. 2001). Plant breeding approaches used to in-crease crop plant biomass and improved nutrient compositions can be potentially used to increase hyperaccumulator biomass.
Numerous research groups have emphasized transgenic solutions to obtaining appropriate plants for phytoremediation. The general strategy underlying this ap-proach is to introduce genes conferring the ability to tolerate and hyperaccumulate
310 Wendy Ann Peer et al.
Fig. 3. Sulfur and selenium assimilation pathways in plants
toxic metals into larger plants capable of rapid development (reviewed in Salt et al. 1998; Clemens et al. 2002). Ideally, the genes could be introduced into plants which are accustomed to growing in the climate where they will be used. Plants could also be engineered to hyperaccumulate multiple different metals for sites with numerous contaminants. Existing hyperaccumulators could be engineered with increased biomass and metal storage capacity. Many studies have begun to elucidate the molecular mechanisms underlying metal hyperaccumulation and tol-erance in plants (Ellis et al. 2004; Freeman et al. 2004; Pence et al. 2000). While it is possible that these discoveries mark the approaching widespread implementa-tion of transgenic hyperaccumulators at phytoremediation sites, there is still much controversy surrounding the agricultural use of genetically modified organisms. Opposition to the deployment of transgenic plants in the field, especially in the promiscuous cruciferous species most closely related to the majority of hyperac-cumulators, is potentially strong enough to prevent this type of phytoremediation technique from being widely used.
Phytoremediation and hyperaccumulator plants 311 If it were possible to deploy genetically engineered phytoremediators, several issues must be considered before transgenes are introduced into the plants. Tissue-specific gene promoters that optimize transgene expression in tissues of interest must be tested for efficacy. The introduction of expression microarray techniques (Atgenex; Birnbaum et al. 2003) and activation tagged promoter lines (Alvarado et al. 2003) to profile gene expression in different tissues should accelerate this proc-ess. If transgenes from non-plant organisms were to be introduced, the proteins encoded by these transgenes must often be engineered to include signal sequences to correctly target the new proteins to the appropriate cellular compartment. Bizily et al. (2003) and Pilon et al. (2003) have recently shown that proper targeting of trangenically expressed proteins in phytoremediator plants can increase efficiency and remediation capacity.
3 Common elemental contaminants
Elements naturally occur in the earth’s crust in a range of background levels that are generally below the critical load, i.e., the amount of the element above which there is a negative effect on biodiversity and ecosystem function (Table 1). How-ever, the concentrations of elements in localized, naturally occurring metalliferous soils or in depositions from anthropogenic activity (e.g. mining, waste disposal, etc.) are considerably higher (Table 1). In the following section, plant mechanisms of tolerance and/or hyperaccumulation of common elemental contaminants are discussed.
3.1 Arsenic (As)
As is a naturally occurring metalloid, which has been used in pesticides and wood preservatives, leading to As contaminated sites (Meharg and Hartley-Whitaker 2002). For example, a Canberra, Australia suburb has As contamination from a pesticide spill (Ng et al. 1998), and localized soil contaminations resulting from use of As in pressure-treated lumber have been widely reported. In the alluvial planes of Bangladesh and West Bengal, India, As contamination of ground water from microbial degradation of peat has resulted in widespread well contamination and health risk. Irrigation has dispersed As contamination to surrounding soils, re-sulting in As poisoning of humans and other animals (McArthur et al. 2001). Similar contamination is seen in regions with As in subsoils worldwide.
Arsenite [AsO2- or As(III)] and arsenate [AsO4-3 or As(V)] are the dominant in-organic arsenic moieties found in terrestrial plants. Both forms are phytotoxic, al-though via different mechanisms. Arsenate,the predominant form found in aerobic soils, is a phosphate analog. Formation of ADP-As complexes instead of ATP leads to cell death; arsenite can cause cell death by binding to and inhibiting en-zymes with sulfhydryl groups. Arsenate is often designated as the more phytotoxic of the two arsenic species (Quaghebeur and Rengel 2003; Wang et al. 2002) but
312 Wendy Ann Peer et al.
the relative toxicities are species-specific (Wang et al. 2002a). As tolerant clones of the grass H. lanatus have a smaller proportion of their total As as arsenite com-pared to non-tolerant clones (Quaghebeur and Rengel 2003), while the As tolerant fern Pteris vittata (Chinese brake fern) almost exclusively accumulates arsenite in its fronds (Webb et al. 2003). While non-accumulators have a phytotoxic thresh-old at approximately 5-100 mg kg-1 As dry weight, H. lanatus can accumulate up to 560 mg kg-1 As(Porter 1975), and P. vittata can accumulate up to 27,000 mg kg-1 As dry weight, with phytotoxic symptoms appearing around 10,000 mg kg-1 As dry weight (Gumaelius et al. 2004). Pteris cretica, Pteris longifolia, and Pteris umbrosa are fern species that also hyperaccumulate As.
3.1.1 Uptake into the root
Pteris vittata accumulates As in contaminated and non-contaminated soils sug-gesting that hyperaccumulation is a constitutive trait (Wang et al. 2002). Arsenic accumulation is correlated with the phosphorus concentration of the media sur-rounding P. vittata. Wang et al. (2002) found that phosphate starvation resulted in a 2.5 fold increase in As net uptake, while the presence of phosphate in the media decreased arsenate influx. This increase in P/As uptake under starvation condi-tions is mediated by an increase in phosphate transporter gene expression and a consequent increase in the amount of protein (Liu et al. 1998; Muchhal and Rag-hothama 1999; Poynton et al. 2004) (For an in depth description of metal-dependent transcriptional regulation, see Chapter 12). Arsenite does not compete with phosphate for uptake into P. vittata roots, suggesting that there is another mechanism for arsenite uptake (Wang et al. 2002).
3.1.2 Transport to the leaves
In Pteris vittata, arsenite is more efficiently translocated from roots to fronds than arsenate (Wang et al. 2002). In B. juncea, addition of the dithiol As chelator dimercaptosuccinate to nutrient solution increased arsenite transport from roots to shoots (Pickering et al. 2000), but it is not yet clear whether arsenite is complexed before xylem loading and transport in P. vittata (Wang et al. 2002). Consistent with arsenate competition for phosphate transport sites, high phosphorus levels were shown to result in significantly decreased As concentrations in P. vittata fronds (Wang et al. 2002).
3.1.3 Detoxification/Chelation
A detoxification pathway for arsenate (AsO4-3) by conversion to arsenite (AsO2-) upon its uptake into roots has been proposed (Meharg and Hartley-Whitaker 2002). Arsenate can be reduced to arsenite enzymatically by arsenate reductase as shown in vitro (Delnomdedieu et al. 1994) and non-enzymatically by glutathione (GSH) or ascorbic acid as in yeast (Mukhopadhyay et al. 2000) followed by the formation of an arsenite-thiol (AsO2--SH) complex. Phytochelatins (PCs) have also been proposed as As chelators in H. lanatus (Raab et al. 2004), and it has
Phytoremediation and hyperaccumulator plants 313 been suggested that arsenite-PC complexes are stored in the vacuole (Meharg and Hartley-Whitaker 2002). It does not appear that arsenite-PC complexes are the dominant form of arsenite in P. vittata as neither PC nor total S are present in suf-ficient quantities in P. vittata for the expected As:thiol ratio which is found in populations of As tolerant non-accumulators (Zhao et al. 2002, 2003). Therefore, if PCs are important for Pteris hyperaccumulation, they may function as a cyto-plasmic shuttle and not a storage complex (Raab et al. 2004). Furthermore, X-ray spectroscopy showed that, at most, 20% of the As in the fronds is coordinated to S suggesting that most of the As stored in the vacuole is aqueous, but uncomplexed with thiols (Lombi et al. 2002b).
3.1.4 Sequestration
Arsenic species are thought to be sequestered in extra- or sub-cellular compart-ments in P. vittata to prevent interaction between As species and cellular compo-nents. X-ray spectroscopy detected the majority of As intracellularlly in the frond epidermal cells, probably in the vacuole (Lombi et al. 2002b). Arsenite-PC com-plexes in H. lanatus are most likely vacuolar (Meharg and Hartley-Whitaker 2002; Quaghebeur et al. 2003).
3.1.5 Phytoremediation of As
Various studies propose the use of P. vittata for phytoremediation of soil and wa-ter. Field experiments have indicated that P. vittata could remediate contaminated soil sites in 10 years or less (Salido et al. 2003) and can reduce arsenic levels in water to less than 10 μg L-1 (ppb) (Blaylock et al. 2001). In Albuquerque, New Mexico, a study was conducted where the ferns significantly decreased the level of arsenic in samples of the city’s drinking water. While a study of the Pteris spe-cies could be beneficial in lending insight into hyperaccumulation mechanisms, using these plants for phytoremediation is not suggested. This is due to aforemen-tioned evidence that the plants convert arsenate to arsenite. Although this conver-sion could make the arsenic less harmful for the plants, it is more harmful to ani-mals and other organisms that might be exposed to the arsenite through plant contact. The ideal phytoremediator would accumulate arsenic at levels similar to P. vittata, but store it in a less toxic form. Arsenobetaine and arsenocholine, which are the major forms of arsenic found in fish (Lopez 2004), have low toxicity to humans and are readily excreted in urine. Transgenics with the ability to convert the inorganic forms of arsenic to these or similar compounds could be viable phy-tormediators.
Dhankar et al. (2002) were able to greatly increase the arsenic tolerance and ac-cumulation of Arabidopsis with only two genes. Constitutive overexpression of γ-glutamylcysteine synthetase (γ-ECS) from the glutathione biosynthesis pathway coupled with the leaf specific expression of arsenate reductase (arsC) from E. coli increased the fresh weight of arsenate challenged plants by ~5-fold and the shoot accumulation ~3-fold. While these significant improvements were not enough to make Arabidopsis into a viable phytoremediator, this shows promise for adding
314 Wendy Ann Peer et al.
arsenic tolerance and extraction capabilities to other hyperaccumulator species (Dhankher et al. 2002).
3.2 Cadmium (Cd)
Cd is a toxic metal and probable carcinogen associated with Zn mining and indus-trial operations where Cd has been used to prevent corrosion of machinery. Re-sulting air-borne Cd dust presents a significant health hazard. Ecotypes of T. caerulescens accumulate a wide range of Cd levels.The Ganges and Vivez eco-types can accumulate up to 10,000 mg kg-1 Cd dry weight and 12, 500 mg kg-1 Cd dry weight, respectively, without showing signs of toxicity; however, the Puy de Wolf and Prayon ecotypes only accumulate 2,300 mg kg-1 Cd dry weight and 4,800 mg kg-1 Cd dry weight, respectively (Lombi et al. 2000, 2001a, 2001b; Peer et al. 2003). Hyperaccumulation of Cd in Arabidopsis hallerii has also been re-ported (Cosio et al. 2004; Kupper et al. 2000). However, reports of hyperaccumu-lation of Cd in B. juncea are questionable, although some Cd accumulation in this species is evident (Salt et al. 1997).
3.2.1 Uptake into the root
Cd uptake is likely mediated through transporters or channels for other divalent ions (Cosio et al. 2004). Several of the Zn and Fe transporting ZIP genes in plants have been shown to transport Cd, although with a wide range of affinities (Grotz et al. 1998; Pence et al. 2000; Ramesh et al. 2003; Vert et al. 2001). Excess diva-lent cations in the media, such as Zn, can reduce Cd uptake in many plant species, including T. caerulescens Prayon (Lombi et al. 2002a, 2001a). Significantly, diva-lent cations and Ca channel blockers had no effect on the Cd uptake of T. caerulescens Ganges, suggesting that this ecotype may have developed a novel Cd uptake system (Lombi et al. 2001a, 2002a).
3.2.2 Transport to the leaves
Pi?eros and Kochian (2003) demonstrated that T. caerulescens and T. arvense mesophyl cells exhibit different plasma membrane ion transport properties, but the differences cannot be directly linked to the differences in Zn and Cd accumula-tion. Analysis of Cd/Zn transport capacity in leaf mesophyll protoplasts demon-strated that the constitutive transport capacity and affinity for these metals were indistinguishable in T. caerulescens Ganges, A. halleri, and T. caerulescens Prayon; however, Cd accumulation increased in Ganges protoplasts but decreased in A. halleri protoplasts in conjunction with Cd pre-exposure (Cosio et al. 2004). Therefore, there may be multiple Cd transport systems in leaves. This suggests that in addition to its novel Cd root uptake pathway, Ganges has developed mechanisms in leaves to facilitate hyperaccumulation.
Phytoremediation and hyperaccumulator plants 315
3.2.3 Detoxification/Chelation
Upon Cd exposure, Nicotiana tabaccum hairy roots had 5 times more reactive oxygen species (ROS)than T. caerulescens hairy roots (Boominathan and Doran 2003a). As GSH has been shown to act as an antioxidant in other species, it was hypothesized that increased GSH synthesis might account for increased tolerance in T. caerulescens. However, exposure to the GSH synthesis inhibitor buthionine sulfoximine(BSO) did not significantly affect ROS levels in T. caerulescens compared to controls, suggesting that GSH was not required for Cd tolerance in T. caerulescens (Boominathan and Doran 2003). Thlaspi caerulescens has constitu-tively high levels of antioxidant enzyme activity like catalase, 300-fold higher than N. tabaccum (Boominathan and Doran 2003), therefore, this may contribute to Cd tolerance. While Cd treatment does not induce phytochelatin (PC) synthesis in non-tolerant plants like A. thaliana, most metal tolerant plants do not accumu-late phytochelatin-metal complexes in response to metal toxicity (Cobbett and Goldsbrough 2002). Although T. caerulescens and T. arvense had increased PCs following Cd treatment, total PCs were lower in the hyperaccumulator T. caerulescens, and PC levels did not correlate with increased tolerance in this plant (Ebbs et al. 2002) (For a detailed discussion of phytochelatins, see Chapter 10). 3.2.4 Sequestration
Few studies have addressed the sequestration of Cd. Vázques et al. (1992) found Cd in the apoplast and vacuoles of T. caerulescens, and most Cd in T. caerules-cens hairy roots appears to be localized in the cell walls (Boominathan and Doran 2003). More recently, Cosio et al. (2005) demonstrated that ~35% Cd taken up ac-cumulates in the cell wall/apoplast in T. caerulescens leaves.
3.2.5 Phytoremediation of Cd
Pilot studies of Cd and Zn phytoremediation have been attempted with contami-nated soils collected from a Zn smelter site in Palmerton, PA (Brown et al. 1994), a Zn smelter in France, sewer sludge contaminated agricultural soil from the UK (Lombi et al. 2001b), and a site contaminated by mine tailings in Silver Bow Creek, MT (Ebbs et al. 1997). In each case, T. caerulescens removed Cd and Zn from the soils, but at rates that would require more than 15 years to remove most of the metals, and only from a narrow soil horizon. In the UK, agricultural soil, Cu toxicity limited the growth of T. caerulescens, demonstrating the need for phy-toremediator plants that can tolerate toxic concentrations of multiple pollutants (Lombi et al. 2001b). The remediation potential of T. caerulescens is also limited by its small stature and biomass. In one study, even though T. caerulescens accu-mulated the highest concentrations of Cd and Zn in its leaves, B. juncea removed more Zn and equivalent amounts Cd due to its larger size (Ebbs et al. 1997). Transgenic approaches to either make T. caerulescens grow larger or to make B. juncea accumulate more Cd and Zn could make Cd phytoextraction feasible.
316 Wendy Ann Peer et al.
Brassica juncea plants genetically modified with bacterial genes to overpro-duce γ-glutamylcysteine synthetase (ECS) or glutathione synthetase (GS) were found to accumulate 1.5 times more Cd and Zn compared to wild type B. juncea growing on metal-contaminated soil from a USEPA Superfund site near Leadville, CO (Bennett et al. 2003). Both ECS and GS overexpressors were able to remove up to 25% of the soil Cd, and Bennet et al. (2003) predict that such transgenic plants should be 1.5 to 3 fold more efficient in phytoextraction than wild type plants.
Zhu et al. (1999) overexpressed the E. coli gene gshI (with a chloroplast target-ing sequence) in B. juncea, which resulted in five times the ECS activity in the transgenic plants compared to wild type. Transgenic seedlings also had increased Cd tolerance, PCs, γ-glutamylcysteine (γ-GluCys), GSH, total non-protein thiols, and Cd accumulation (40-90%). These results indicate that ECS is important in Cd accumulation and tolerance and that overexpressing ECS could potentially be an effective phytoremediation strategy (Zhu et al. 1999).
Lee et al. (2003) overexpressed an Arabidopsis PC synthase (AtPCS1) in Arabidopsis resulting in 1.3 to 2.1-fold increase PCs; however, the transgenic lines were hypersensitive to Cd stress as measured by root growth and this hypersensi-tivity could be alleviated by the addition of glutathione. This suggests that the regulation of glutathione levels and perhaps the entire S assimilation pathway is important for Cd tolerance and accumulation. In a contrasting study, PC-deficient Arabidopsis(cad1-3) were transformed with wheat TaPCS1 cDNA (Gong et al. 2003). While the Cd sensitivity of the transgenic cad1-3 plants was complemented by TaPCS1 expression, these TaPCS1 expressing plants accumulated less Cd than the cad1-3 mutants but increased Cd transport to leaves (Gong et al. 2003). Fi-nally, Song et al. (2003) report that expression of the yeast vacuolar glutathione-Cd transporter YCF1 in Arabidopsis increased biomass and Cd uptake was ~2-fold greater than wild type. In addition, preliminary data suggest that expressing YCF1 in poplar also increases biomass (Song et al. 2003). Thus far, transgenic plants do not have sufficiently high levels of accumulation needed for significant Cd phytoremediation; however, a combined approach could result in an effective Cd phytoremediation technology.
3.3 Chromium (Cr)
Cr(III) is an essential nutrient for animals and is present in many oxidation states in the environment [Cr(II) to Cr(VI)], including the most common forms Cr(0), Cr(III), and Cr(VI). Cr(VI) and the steel component Cr(0) are usually byproducts of industrial processes. Cr(VI) is considered to be 1,000 times more toxic than Cr(III), and the World Health Agency and EPA has determined that Cr(IV) is a carcinogen. Cr(VI) contamination in the soil and groundwater has been detected in various parts of southern California and in Tennessee (EPA 2000; EPA 2004), where wells were closed because Cr(IV) levels exceeded EPA limits (Table 1). Areas in and around Glasgow, Scotland are also heavily contaminated by Cr used
Phytoremediation and hyperaccumulator plants 317 for ore processing in the 19th century which subsequently leached into the ground water supply and accumulated at toxic levels (Farmer et al. 1999).
3.3.1 Phytoremediation of Cr
Although Cr(IV) is oxidized to the relatively non-hazardous Cr(III) in soil (ATSDR, 2001), Cr(IV), and Cr(VI) in groundwater and estuaries can pose health hazards and disrupt ecosystems. Betula and Salix trees are able to take up Cr, and therefore, would be useful for phytoremediation of Cr(IV) contaminated ground-water (Pulford et al. 2001) while Cr(VI) in estuaries can be absorbed by coconut husks and bagasse (Krishnani et al. 2004; Parimala et al. 2004). A recent report of Salsola kali (tumbleweed) accumulating Cr(VI) may prove to be useful for phy-toremediation of Cr(VI) in the soil (Gardea-Torresdey et al. 2005).
3.4 Copper (Cu)
Cu is an essential element and enzyme co-factor for oxidases (cytochrome c oxi-dase, superoxide dismutase) and tyrosinases; however, animals and plants can ac-cumulate toxic levels of Cu. Cu contamination for soil and groundwater usually results from mine sites like Blackbird Creek, ID (Mebane 1997) or munitions re-search or disposal sites like the Picatinny Arsenal, in northern New Jersey (EPA 1989). The Deer Lodge Valley and Anaconda areas of the Beaverell pedon in Montana, a region affected by long-term Cu smelting, has soil polluted with high levels of Cu and lower concentrations of As, Zn, and Pb (Burt et al. 2000, 2003). Cu contamination is also problematic in European soils exposed to historical min-ing and smelting activity. During the 19th century, the Devon Great Consols Mine in the Tamar Valley of Devon, UK, was the world’s largest producer of Cu. As a result, Cu in the form of chalcopyrite has permeated the soil in this region (Lombi et al. 2004). In France, due to the continuous treatment of vine downy mildew with Bordeaux mix since the end of the 19th century, extensive deposits of Cu have accumulated in many vineyard soils (Brun et al. 2001). In Kayseri, Anatolia, high Cu concentrations were found in Zn-producing industrial sites (Aksoy et al. 2000), while a study in Denizli found that high levels of Cu characterize urban roadsides associated with road traffic (Celik et al. 2005).Cyprus (Kupros) has an extensive history of Cu mining extending back over 2,000 years. Cu mining wastes from this prolonged activity have had a significant impact on the environ-ment and biota of this island (Pyatt 2001). Greece is also afflicted by Cu contami-nation: lake ecosystems, such as Lake Pamvotis in north-western Greece, exhibit high concentrations of this metal (Papagiannis et al. 2004), and Cu poisoning from high Cu content in local food has occurred in areas such as Veria county (Zan-topoulos et al. 1999).
318 Wendy Ann Peer et al.
3.4.1 Phytoremediation of Cu
At superoptimal levels, Cu is highly toxic to plants and Cu ligands in plants are citrate, PC2, PC3, and metallothioniens (Murphy et al. 1999; Rauser 1999). Corre-spondingly, most Cu-tolerant plants are excluders, and no confirmed Cu accumu-lators have been identified. It was originally thought that Elsholtzia splendens was a Cu hyperaccumulator, but after further investigation it was concluded to be a tolerant excluder like Elsholtzia argyi (Jiang et al. 2004b), Silene vulgaris (Song et al. 2004) and Mimiulus guttatus (Harper et al. 1998).While 37 taxa of Cu hyper-accumulators have been reported, mainly from the Shaban Copper Arc of Congo, more research is needed to determine if the high Cu levels are due to hyperaccu-mulation or depositions of Cu dust on leaves (Song et al. 2004). Salix nigra was shown to accumulate more Cd and Cu than other Salix species, and field studies should determine the feasibility of this species for phytoremediation (Kuzovlina et al. 2004). In addition, soil amendments, like phosphate, increase Cu uptake, and therefore, may further phytoremediation efforts (Wu et al. 2004) (For a detailed discussion of Cu in plants, see Chapter 9). For aqueous Cu contamination, Eich-hornia crassipes, the water hyacinth, is estimated to absorb 21.62 kg Cu ha-1, and could potentially be used for phytoremediation of low level Cu contamination in waste water (Liao and Chang 2004).
3.5 Mercury (Hg)
Mercury is toxic to humans, and depending on the form it takes can cause severe neurological disorders (Carty and Malone 1979). Mercury exposure to humans is mostly through ingestion of fish, as Hg is biomagnified through the aquatic food chain, and amalgam dental fillings. Over the past century, several thousand tons of Hg have been released to the environment by human activity (Andren and Nriagu 1979). Many developing countries use elemental Hg-Au amalgamation mining practices, which results in significant Hg contamination of surrounding soil and water. Examples include artisanal Au mines in Suriname and the Amazon basin (Gray et al. 2002). Mercury in soil can be converted to cinnabar (HgS) as a result of sulfate reduction after the deposition and burial of mercury-contaminated soils. However, mercury release from solid forms, such as cinnabar, can also create en-vironmental hazards. Studies of contaminated soil from industrial mercury dump-ing at the headwaters of East Fork Poplar Creek in Oak Ridge, TN, and in a Flor-ida Everglades study indicate that organic matter could increase mercury mobilization from cinnabar and affect mercury bioavailability (Barnett et al. 1997; Ravichandran et al. 1998). These are candidate areas for phytoremediation.
The most toxic forms of mercury are organomercurials like methyl-Hg and phenyl mercuric acetate, followed by ionic Hg(II), with elemental Hg(0) as the least toxic form. Organomercurials and ionic Hg are toxic to plants, and to date Hg hyperaccumulating plants have not been identified. However, a Hg hyperaccumu-lating mushroom Amanita muscaria has been found that accumulates 96-1900 ng g-1 dry wt in the caps and 61-920 ng g-1 dry wt in the stalks depending on the col-
条码式寄存柜(储物柜)产品技术说明 一、适用范围 自助行李寄存柜,用于高效率、规范化且消费群体较密集的高档公共场所,如:超市、学校、机场、俱乐部、游泳馆、健身房、超市、大卖场、商场、事业单位等。客户可根据自身特点选择不同类型的机组,有条码型、指纹型、IC卡、条形码卡、磁卡、射频卡以及TM扣等,或者使用客户各自所持有的会员卡。 二、基本组成 自助行李寄存柜,是由嵌入式计算机进行控制,具有管理功能的小件物品寄存系统。主要由输入设备(条码阅读器)、输出设备(如中文液晶显示)、嵌入式计算机处理中心、管理软件以及电源组成。 三、结构、材料、钣金说明 寄存柜柜体选用0.8mm以上优质冷轧板,经冷加工成形后,用二氧化碳气体保护焊焊接装配而成,柜体结构坚固结实。箱体表面经除锈、除油、打磨、磷化处理后喷塑,塑面的颜色可由用户选定。?箱门背面增加纵向加强筋,提高箱门的防撞击能力;能有效防止和降低使用者因疏忽碰伤、磕伤,电控锁采用360度具有防撬、带防软片插入装置。 四、元器件说明 1)电控锁:经过10万次寿命测试无故障,性能可靠稳定。部件的表面均作热 涂锌防锈处理,装配成形后每把锁均作电气绝缘试验和开启试验,并按加工批量的1%作寿命试验。 2)扫描枪:台湾欣技C1000,性能稳定,读码速度快。 3)液晶显示屏:3.5寸彩色显示屏。例如:
表示:空箱,顾客可以存入的箱子 表示:满箱,有合法密码,箱内有物 4)打印机:自主研发使用日本精工打印机芯。 5)控制主板:自主研发生产。 6)切纸刀:自主研发生产、10万次寿命测试,性能稳定,配有走纸导向。 7)控制芯片:采用国际上著名大公司的工业级产品,有ATMEL公司、INTEL公 司、日立公司、德州公司等。 8)键盘:选用电话机水晶键盘,工作可靠,永不退色,使用寿命长。 五、常规技术指标 (1)功率-待机:25W 开箱:60W (2)电源电压:AC110V—AC240V 50HZ (3)后备电源(可选):18V 7AH (4)打印速度:35mm/s (5)打印机寿命:50Km (6)柜体冷轧钢板厚度:0.8mm (7)读码速度:<0.4s (8)显示分辨率:3.5寸彩色液晶屏(可定制显示LOGO) 六、条码寄存柜主要功能说明 ?大屏幕液晶显示屏 操作步骤提示、箱门5种状态显示、时钟显示、常规故障指示、设置菜单、管理界面、工作状态显示等。 在现场安装就位后,确认各个部件正常后,即可上电开机,完整的显示: ?全开放式中文设置菜单(可定制多国语言、语音) 寄存柜的各个管理参数全部面向客户开放,客户可选择相应的菜单进行设置。设有三级管理密码,方便不同级别的管理要求。 ?快捷管理员管理菜单 在管理模式下,管理快捷菜单可应急开启箱门、清除箱门ID、查询箱门状态等操作。可设置500位管理员进行管理,每个管理员可根据自己的用户号及管
脐带间充质干细胞的研究进展 间充质干细胞(mesenchymal stem cells,MSC S )是来源于发育早期中胚层 的一类多能干细胞[1-5],MSC S 由于它的自我更新和多项分化潜能,而具有巨大的 治疗价值 ,日益受到关注。MSC S 有以下特点:(1)多向分化潜能,在适当的诱导条件下可分化为肌细胞[2]、成骨细胞[3、4]、脂肪细胞、神经细胞[9]、肝细胞[6]、心肌细胞[10]和表皮细胞[11, 12];(2)通过分泌可溶性因子和转分化促进创面愈合;(3) 免疫调控功能,骨髓源(bone marrow )MSC S 表达MHC-I类分子,不表达MHC-II 类分子,不表达CD80、CD86、CD40等协同刺激分子,体外抑制混合淋巴细胞反应,体内诱导免疫耐受[11, 15],在预防和治疗移植物抗宿主病、诱导器官移植免疫耐受等领域有较好的应用前景;(4)连续传代培养和冷冻保存后仍具有多向分化潜能,可作为理想的种子细胞用于组织工程和细胞替代治疗。1974年Friedenstein [16] 首先证明了骨髓中存在MSC S ,以后的研究证明MSC S 不仅存在于骨髓中,也存在 于其他一些组织与器官的间质中:如外周血[17],脐血[5],松质骨[1, 18],脂肪组织[1],滑膜[18]和脐带。在所有这些来源中,脐血(umbilical cord blood)和脐带(umbilical cord)是MSC S 最理想的来源,因为它们可以通过非侵入性手段容易获 得,并且病毒污染的风险低,还可冷冻保存后行自体移植。然而,脐血MSC的培养成功率不高[19, 23-24],Shetty 的研究认为只有6%,而脐带MSC的培养成功率可 达100%[25]。另外从脐血中分离MSC S ,就浪费了其中的造血干/祖细胞(hematopoietic stem cells/hematopoietic progenitor cells,HSCs/HPCs) [26, 27],因此,脐带MSC S (umbilical cord mesenchymal stem cells, UC-MSC S )就成 为重要来源。 一.概述 人脐带约40 g, 它的长度约60–65 cm, 足月脐带的平均直径约1.5 cm[28, 29]。脐带被覆着鳞状上皮,叫脐带上皮,是单层或复层结构,这层上皮由羊膜延续过来[30, 31]。脐带的内部是两根动脉和一根静脉,血管之间是粘液样的结缔组织,叫做沃顿胶质,充当血管外膜的功能。脐带中无毛细血管和淋巴系统。沃顿胶质的网状系统是糖蛋白微纤维和胶原纤维。沃顿胶质中最多的葡萄糖胺聚糖是透明质酸,它是包绕在成纤维样细胞和胶原纤维周围的并维持脐带形状的水合凝胶,使脐带免受挤压。沃顿胶质的基质细胞是成纤维样细胞[32],这种中间丝蛋白表达于间充质来源的细胞如成纤维细胞的,而不表达于平滑肌细胞。共表达波形蛋白和索蛋白提示这些细胞本质上肌纤维母细胞。 脐带基质细胞也是一种具有多能干细胞特点的细胞,具有多项分化潜能,其 形态和生物学特点与骨髓源性MSC S 相似[5, 20, 21, 38, 46],但脐带MSC S 更原始,是介 于成体干细胞和胚胎干细胞之间的一种干细胞,表达Oct-4, Sox-2和Nanog等多
精神分裂症的病因及发病机理 精神分裂症病因:尚未明,近百年来的研究结果也仅发现一些可能的致病因素。(一)生物学因素1.遗传遗传因素是精神分裂症最可能的一种素质因素。国内家系调查资料表明:精神分裂症患者亲属中的患病率比一般居民高6.2倍,血缘关系愈近,患病率也愈高。双生子研究表明:遗传信息几乎相同的单卵双生子的同病率远较遗传信息不完全相同 的双卵双生子为高,综合近年来11项研究资料:单卵双生子同病率(56.7%),是双卵双生子同病率(12.7%)的4.5倍,是一般人口患难与共病率的35-60倍。说明遗传因素在本病发生中具有重要作用,寄养子研究也证明遗传因素是本症发病的主要因素,而环境因素的重要性较小。以往的研究证明疾病并不按类型进行遗传,目前认为多基因遗传方式的可能性最大,也有人认为是常染色体单基因遗传或多源性遗传。Shields发现病情愈轻,病因愈复杂,愈属多源性遗传。高发家系的前瞻性研究与分子遗传的研究相结合,可能阐明一些问题。国内有报道用人类原癌基因Ha-ras-1为探针,对精神病患者基因组进行限止性片段长度多态性的分析,结果提示11号染色体上可能存在着精神分裂症与双相情感性精神病有关的DNA序列。2.性格特征:约40%患者的病前性格具有孤僻、冷淡、敏感、多疑、富于幻想等特征,即内向
型性格。3.其它:精神分裂症发病与年龄有一定关系,多发生于青壮年,约1/2患者于20~30岁发病。发病年龄与临床类型有关,偏执型发病较晚,有资料提示偏执型平均发病年龄为35岁,其它型为23岁。80年代国内12地区调查资料:女性总患病率(7.07%。)与时点患病率(5.91%。)明显高于男性(4.33%。与3.68%。)。Kretschmer在描述性格与精神分裂症关系时指出:61%患者为瘦长型和运动家型,12.8%为肥胖型,11.3%发育不良型。在躯体疾病或分娩之后发生精神分裂症是很常见的现象,可能是心理性生理性应激的非特异性影响。部分患者在脑外伤后或感染性疾病后发病;有报告在精神分裂症患者的脑脊液中发现病毒性物质;月经期内病情加重等躯体因素都可能是诱发因素,但在精神分裂症发病机理中的价值有待进一步证实。(二)心理社会因素1.环境因素①家庭中父母的性格,言行、举止和教育方式(如放纵、溺爱、过严)等都会影响子女的心身健康或导致个性偏离常态。②家庭成员间的关系及其精神交流的紊乱。③生活不安定、居住拥挤、职业不固定、人际关系不良、噪音干扰、环境污染等均对发病有一定作用。农村精神分裂症发病率明显低于城市。2.心理因素一般认为生活事件可发诱发精神分裂症。诸如失学、失恋、学习紧张、家庭纠纷、夫妻不和、意处事故等均对发病有一定影响,但这些事件的性质均无特殊性。因此,心理因素也仅属诱发因
脐带血造血干细胞库管理办法(试行) 第一章总则 第一条为合理利用我国脐带血造血干细胞资源,促进脐带血造血干细胞移植高新技术的发展,确保脐带血 造血干细胞应用的安全性和有效性,特制定本管理办法。 第二条脐带血造血干细胞库是指以人体造血干细胞移植为目的,具有采集、处理、保存和提供造血干细胞 的能力,并具有相当研究实力的特殊血站。 任何单位和个人不得以营利为目的进行脐带血采供活动。 第三条本办法所指脐带血为与孕妇和新生儿血容量和血循环无关的,由新生儿脐带扎断后的远端所采集的 胎盘血。 第四条对脐带血造血干细胞库实行全国统一规划,统一布局,统一标准,统一规范和统一管理制度。 第二章设置审批 第五条国务院卫生行政部门根据我国人口分布、卫生资源、临床造血干细胞移植需要等实际情况,制订我 国脐带血造血干细胞库设置的总体布局和发展规划。 第六条脐带血造血干细胞库的设置必须经国务院卫生行政部门批准。 第七条国务院卫生行政部门成立由有关方面专家组成的脐带血造血干细胞库专家委员会(以下简称专家委
员会),负责对脐带血造血干细胞库设置的申请、验收和考评提出论证意见。专家委员会负责制订脐带血 造血干细胞库建设、操作、运行等技术标准。 第八条脐带血造血干细胞库设置的申请者除符合国家规划和布局要求,具备设置一般血站基本条件之外, 还需具备下列条件: (一)具有基本的血液学研究基础和造血干细胞研究能力; (二)具有符合储存不低于1 万份脐带血的高清洁度的空间和冷冻设备的设计规划; (三)具有血细胞生物学、HLA 配型、相关病原体检测、遗传学和冷冻生物学、专供脐带血处理等符合GMP、 GLP 标准的实验室、资料保存室; (四)具有流式细胞仪、程控冷冻仪、PCR 仪和细胞冷冻及相关检测及计算机网络管理等仪器设备; (五)具有独立开展实验血液学、免疫学、造血细胞培养、检测、HLA 配型、病原体检测、冷冻生物学、 管理、质量控制和监测、仪器操作、资料保管和共享等方面的技术、管理和服务人员; (六)具有安全可靠的脐带血来源保证; (七)具备多渠道筹集建设资金运转经费的能力。 第九条设置脐带血造血干细胞库应向所在地省级卫生行政部门提交设置可行性研究报告,内容包括:
古代汉语词类活用例句列举 古代汉语词类活用例句列举《郑伯克段于鄢》1、例:壮公生,惊姜氏。P97 惊:用作使动,使。。。惊。2、例:无生民心。P99 生:用作使动,使。。。产生。3、例:若阙地及泉,隧而相见。P101 隧:名词动用。《公孙无知之乱》4、豕立而啼,P109 立:名词作状语,像人一样丫立。〈安之战〉5、皆主?献子。P117 主:名词动用,以。。。为主。6、君无所辱命。P119 辱:动词使动,使。。。受辱。7、从左右,皆肘之。P123 肘:名词使动,表示用胳膊推撞。8、臣辱戎士。123 辱:动词使动。9、人不难以死免其君。P123 免:用作使动,使。。。免于。10、故中御而从齐候。P123 中:方位名词做状语。〈子产说范宣子轻敝〉11、三周华不注。P122 周:
名词动用。12、郑人病之。P129 病:名词用作意动。13、象有齿而焚其身。P130 焚:动词用作使动。14、宣子说,乃轻弊。P130 轻:形容词用作使动,使。。。轻。〈苏秦连横约纵〉15、今先生俨然不运千里而庭教之。P182 远:形容词用作意动。16、明言章理,兵甲愈起。P183 明、章:用作使动。 1 17、辨言伟服。攻战不息。P183 辩、伟:都用作使动,使。。。雄辩,使。。。华美。18、繁称文辞,天下不冶。P183 文:名词用作使动。19、夫徒处而致利,安坐而广地。P183 广:形容词用作使动,使。。。广。20、言语相结,天下为一。P183 言语:名词作状语。21、今欲并天下,凌万乘,诎敌国。制海内,子元元。臣诸候。非兵不可。P183 诎:用作使动,使。。。屈服;子:名词用作使动,使。。。成为子女;臣:名词用作使动,使。。。成为臣子。22、约纵散横,以抑强秦。
存包柜项目方案说明 一、项目基本信息 (一)项目名称 存包柜项目 (二)项目建设单位 xxx有限公司 (三)法定代表人 于xx (四)公司简介 本公司奉行“客户至上,质量保障”的服务宗旨,树立“一切为客户着想” 的经营理念,以高效、优质、优惠的专业精神服务于新老客户。 公司依托集团公司整体优势、发展自身专业化咨询能力,以助力产业提高运营效率为使命,提供全方面的业务咨询服务。 公司将加强人才的引进和培养,尤其是研发及业务方面的高级人才,健全研发、管理和销售等各级人员的薪酬考核体系,完善激励制度,提高公司员工创造力,为公司的持续快速发展提供强大保障。
上一年度,xxx(集团)有限公司实现营业收入7026.99万元,同比增长25.92%(1446.42万元)。其中,主营业业务存包柜生产及销售收入为5749.34万元,占营业总收入的81.82%。 根据初步统计测算,公司实现利润总额1653.53万元,较去年同期相比增长375.52万元,增长率29.38%;实现净利润1240.15万元,较去年同期相比增长139.78万元,增长率12.70%。 (五)项目选址 xx新兴产业示范基地 (六)项目用地规模 项目总用地面积12559.61平方米(折合约18.83亩)。 (七)项目用地控制指标 该工程规划建筑系数71.28%,建筑容积率1.01,建设区域绿化覆盖率7.20%,固定资产投资强度188.57万元/亩。 项目净用地面积12559.61平方米,建筑物基底占地面积8952.49平方米,总建筑面积12685.21平方米,其中:规划建设主体工程7943.44平方米,项目规划绿化面积913.28平方米。 (八)设备选型方案 项目计划购置设备共计91台(套),设备购置费1292.40万元。 (九)节能分析 1、项目年用电量486324.58千瓦时,折合59.77吨标准煤。
古代汉语练习(词类活用) 班级:姓名:学号: 一、简答: 1、什么是古代汉语的词类活用?古代汉语中的词类活用有哪几种? 2、怎样区别使动用法和意动用法?试举例说明。并说明如何翻译。 3、试说明名词做状语主要有哪几种情况。 4、名词、形容词用作动词的情况主要有哪些?应该如何辨认? 二、多项选择题(在每小题的四个备选答案中,选出二个至四个正确的答案,并将其号码分别填在题干后的括号内,多选、少选、错选均无分。每小题1分,共5分) 1.下列各句中加着重号的词,属于词类活用的是() A.斩一首者爵一级B.能富贵将军者,上也 C.曹人凶俱,为其所得者棺而出之 D.夫鼠,昼伏夜动,不穴于寝庙,畏人故也 2.下列各句中加着重号的词属于名词作状语的是() A.裂裳衣疮,手往善药 B.其经承子厚口讲指画为文词者,悉有法度可观 C.范增数目项王D.诸侯宾至 3.下列各句含宾语前置现象的是() A.姜氏何厌之有B.楚君之惠,末之敢忘 C.除君之恶,唯力是视D.昭王南征而不复,寡人是问 4.对下列各句中加着重号的词组分析错误的是() A.子重使太宰伯州犁待于王后(动宾)B.将塞井夷灶而为行也(连动) C.臣之壮也犹不如人(主谓)D.以勇力之所加而治智能之官(偏正) 5.下列句子中有使动用法的是() A.秋九月,晋侯饮赵盾酒,伏甲将攻之 B.是时万石君奋为汉王中涓,受手谒,人见平
C.见灵辄饿,问其病,曰:“不食三日矣。”食之,舍其半 D.仓廪实而知礼节,衣食足而知荣辱 四、指出并具体说明下列文句中的词类活用现象: 1.秦数败赵军,赵军固壁不战。(秦与赵兵相距长平) 2.赵王不听,遂将之。(秦与赵兵相距长平) 3.身所奉饭饮而进食者以十数,所友者以百数。(秦与赵兵相距长平) 4.括军败,数十万之众遂降秦,秦悉阬之。(秦与赵兵相距长平) 5.信数与萧何语,何奇之。(韩信拜将) 6.王必欲长王汉中,无所事信。(韩信拜将) 7.吾亦欲东耳,安能郁郁久居此乎?(韩信拜将) 8.何闻信亡,不及以闻,自追之。(韩信拜将) 9.今大王举而东,三秦可传檄而定也。(韩信拜将) 10.遇有以梦得事白上者,梦得于是改刺连州。(柳子厚墓志铭) 11.自子厚之斥,遵从而家焉,逮其死不去。(柳子厚墓志铭) 12.以如司农治事堂,栖之梁木上。(段太尉逸事状) 13.踔厉风发,率常屈其座人。(柳子厚墓志铭) 14.晞一营大噪,尽甲。(段太尉逸事状) 15.即自取水洗去血,裂裳衣疮,手注善药。(段太尉逸事状) 16.黄罔之地多竹,大者如椽。竹工破之,刳去其节,用代陶瓦。(黄冈竹楼记)17.晋灵公不君。厚敛以彫墙。(晋灵公不君) 18.既而与为公介,倒戟以御公徒而免之。(晋灵公不君) 19.盛服将朝,尚早,坐而假寐。(晋灵公不君) 20.晋侯饮赵盾酒,伏甲将攻之。(晋灵公不君) 五、说明下列文句中的词类活用现象,并将全文译为现代汉语:
精神分裂症的发病原因是什么 精神分裂症是一种精神病,对于我们的影响是很大的,如果不幸患上就要及时做好治疗,不然后果会很严重,无法进行正常的工作和生活,是一件很尴尬的事情。因此为了避免患上这样的疾病,我们就要做好预防,今天我们就请广州协佳的专家张可斌来介绍一下精神分裂症的发病原因。 精神分裂症是严重影响人们身体健康的一种疾病,这种疾病会让我们整体看起来不正常,会出现胡言乱语的情况,甚至还会出现幻想幻听,可见精神分裂症这种病的危害程度。 (1)精神刺激:人的心理与社会因素密切相关,个人与社会环境不相适应,就产生了精神刺激,精神刺激导致大脑功能紊乱,出现精神障碍。不管是令人愉快的良性刺激,还是使人痛苦的恶性刺激,超过一定的限度都会对人的心理造成影响。 (2)遗传因素:精神病中如精神分裂症、情感性精神障碍,家族中精神病的患病率明显高于一般普通人群,而且血缘关系愈近,发病机会愈高。此外,精神发育迟滞、癫痫性精神障碍的遗传性在发病因素中也占相当的比重。这也是精神病的病因之一。 (3)自身:在同样的环境中,承受同样的精神刺激,那些心理素质差、对精神刺激耐受力低的人易发病。通常情况下,性格内向、心胸狭窄、过分自尊的人,不与人交往、孤僻懒散的人受挫折后容易出现精神异常。 (4)躯体因素:感染、中毒、颅脑外伤、肿瘤、内分泌、代谢及营养障碍等均可导致精神障碍,。但应注意,精神障碍伴有的躯体因素,并不完全与精神症状直接相关,有些是由躯体因素直接引起的,有些则是以躯体因素只作为一种诱因而存在。 孕期感染。如果在怀孕期间,孕妇感染了某种病毒,病毒也传染给了胎儿的话,那么,胎儿出生长大后患上精神分裂症的可能性是极其的大。所以怀孕中的女性朋友要注意卫生,尽量不要接触病毒源。 上述就是关于精神分裂症的发病原因,想必大家都已经知道了吧。患上精神分裂症之后,大家也不必过于伤心,现在我国的医疗水平是足以让大家快速恢复过来的,所以说一定要保持良好的情绪。
基于单片机的自动存包系统设计 摘要 近年来,随着生活水平的提高,人们对于社会消费品的质量和数量的要求也在逐渐增加。为了更好的为广大顾客服务,在一些商场、影院、超市等公共场合通常设置有自动存包柜,本次便是针对这一现象进行设计。 本文详细介绍了国内自动存包控制系统的发展现状,发展中所面临的问题。并详细介绍了本系统采用的AT89S52单片机做控制器,可以同时管理四个存包柜。柜门锁是由继电器控制,当顾客需要存包的时候,可以自行到存包柜前按“开门”键,需要顾客向光学指纹识别系统输入个指纹,然后通过继电器进行开门(用亮灯表示),顾客即可存包,并需将柜门关上。当顾客需要取包时,要将只要将之前输入的指纹放置于指纹识别器前方,指纹识别器采集到指纹信息输出相应的高低电平信号传给单片机,系统比较密码一致后,发出开箱信号至继电器将柜门打开,顾客即可将包取出。它具有功能实用、操作简便、安全可靠、抗干扰性强等特点。 关键词:自动存包柜,单片机,指纹识别器
李少鹏:基于单片机的自动存包系统设计 Based on single chip microcomputer automatic package design Abstract In recent years, with the improvement of living standards, people for social consumer goo ds quality and quantity requirements are to increase gradually. In order to better service for the g eneral customers, in some stores, movie theaters, supermarkets public Settings are to be put auto matically usually bag ark, it is functional practical, simple operation, safe and reliable, anti-jamm ing strong sexual characteristics. Domestic deposit automatic control system are introduced in detail in this paper the development of the status quo, problems faced in the development of. And introduces in detail the system adopts single chip microcomputer controller, can simultaneously manage multiple pack ark. Cupboard door lock controlled by relay, when customers need to save package, will be allowed to save package before the ark according to the "open" button, need customer to the system input fingerprint, and then through the relay to open the door (with lighting), customers can save package, and the cupboard door must be closed. When customers need to pick up package, as long as before the input fingerprint should be placed on the fingerprint recognizer, fingerprint recognizer collecting to the fingerprint information and output the corresponding high and low level signal to the microcontroller, the system is password consistent, signal out of the box to the relay Key words: Automatic Storage Bag, Microcontroller, Fingerprint recognizer。
初中所学文言文中的五类常见词类活用现象
古代汉语中的词类活用现象 五种类型:名词用作动词 动词、形容词、名词的使动用法 形容词、名词的意动用法 名词用作状语 动词用作状语 (一)名词用如动词 古代汉语名词可以用如动词的现象相当普遍。如: 从左右,皆肘.之。(左传成公二年) 晋灵公不君.。(左传宣公二年) 孟尝君怪其疾也,衣冠 ..而见之。(战国策·齐策四) 马童面.值,指王翳曰:“此项王也。”(史记·项羽本纪) 夫子式.而听之。(礼记·檀弓下) 曹子手.剑而从之。(公羊传庄公十三年) 假舟楫者,非能水.也,而绝江河。(荀子·劝学) 左右欲刃.相如。(史记·廉颇蔺相如列传) 秦师遂东.。(左传僖公三十二年) 汉败楚,楚以故不能过荥阳而西.。(史记·项羽本纪) 以上所举的例子可以分为两类:前八个例子是普通名词用如动词,后两个例子是方位名词用如动词。 名词用作动词是由上下文决定的。我们鉴别某一个名词是不是用如动词,须要从整个意思来考虑,同时还要注意它在句中的地位,以及它前后有哪些词类的词和它相结合,跟他构成什么样的句法关系。一般情况有如下四种:
①代词前面的名词用如动词(肘之、面之),因为代词不受名词修饰; ②副词尤其是否定副词后面的名词用如动词(“遂东”、“不君”); ③能愿动词后面的名词也用如动词(“能水”、“欲刃”); ④句中所确定的宾语前面的名词用如动词(“脯鄂侯”“手剑”) (二)动词、形容词、名词的使动用法 一、动词的使动用法。 定义:主语所代表的人物并不施行这个动词所表示的动作,而是使宾语所代表的人或事物施行这个动作。例如:《左传隐公元年》:“庄公寤生,惊姜氏。”这不是说庄公本人吃惊,而是说庄公使姜氏吃惊。 在古代汉语里,不及物动词常常有使动用法。不及物动词本来不带宾语,当它带有宾语时,则一定作为使动用法在使用。如: 焉用亡.郑以陪邻?《左传僖公三十年》 晋人归.楚公子榖臣与连尹襄老之尸于楚,以求知罃。(左传成公三年) 大车无輗,小车无杌,其何以行.之哉?《论语·为政》 小子鸣.鼓而攻之可也。《论语·先进》 求也退,故进.之;由也兼人,故退.之。《论语·先进》 故远人不服,则修文德以来.之。《论语·季氏》 有时候不及物动词的后面虽然不带宾语,但是从上下文的意思看,仍是使动用法。例如《论语·季氏》:“远人不服而不能来也”这个“来”字是使远人来的意思。 古代汉语及物动词用如使动的情况比较少见。及物动词本来带有宾语,在形式上和使动用法没有什么区别,区别只在意义上。使动的宾语不是动作的接受者,而是主语所代表的人物使它具有这种动作。例如《孟子·梁惠王上》“朝秦楚”,不食齐宣王朝见秦楚之君,相反的,是齐宣王是秦楚之君朝见自己。 下面各句中的及物动词是使动用法: 问其病,曰:“不食三日矣。”食.之。《左传·宣公二年》
精神分裂症的病因是什么 精神分裂症是一种精神方面的疾病,青壮年发生的概率高,一般 在16~40岁间,没有正常器官的疾病出现,为一种功能性精神病。 精神分裂症大部分的患者是由于在日常的生活和工作当中受到的压力 过大,而患者没有一个良好的疏导的方式所导致。患者在出现该情况 不仅影响本人的正常社会生活,且对家庭和社会也造成很严重的影响。 精神分裂症常见的致病因素: 1、环境因素:工作环境比如经济水平低低收入人群、无职业的人群中,精神分裂症的患病率明显高于经济水平高的职业人群的患病率。还有实际的生活环境生活中的不如意不开心也会诱发该病。 2、心理因素:生活工作中的不开心不满意,导致情绪上的失控,心里长期受到压抑没有办法和没有正确的途径去发泄,如恋爱失败, 婚姻破裂,学习、工作中不愉快都会成为本病的原因。 3、遗传因素:家族中长辈或者亲属中曾经有过这样的病人,后代会出现精神分裂症的机会比正常人要高。 4、精神影响:人的心里与社会要各个方面都有着不可缺少的联系,对社会环境不适应,自己无法融入到社会中去,自己与社会环境不相
适应,精神和心情就会受到一定的影响,大脑控制着人的精神世界, 有可能促发精神分裂症。 5、身体方面:细菌感染、出现中毒情况、大脑外伤、肿瘤、身体的代谢及营养不良等均可能导致使精神分裂症,身体受到外界环境的 影响受到一定程度的伤害,心里受到打击,无法承受伤害造成的痛苦,可能会出现精神的问题。 对于精神分裂症一定要配合治疗,接受全面正确的治疗,最好的 疗法就是中医疗法加心理疗法。早发现并及时治疗并且科学合理的治疗,不要相信迷信,要去正规的医院接受合理的治疗,接受正确的治 疗按照医生的要求对症下药,配合医生和家人,给病人创造一个良好 的治疗环境,对于该病的康复和痊愈会起到意想不到的效果。
自动存包柜的设计与仿真 摘要 本课题是基于单片机的自动存包柜设计。自动存包柜是新一代的存包柜,具有功能实用、操作简单、管理方便、安全可靠等特点,能够更好的服务于不同市场的广大群众,使用者可以根据简明清晰的操作说明自行完成存包取包工作。本系统由MCS-51单片机构成核心控制系统,整个系统由主控部分、键盘显示控制部分、执行部分三部分组成,通过随机密码的产生和核对完成自动存包取包过程。本设计中各元器件便于安装且操作简单,能基本实现存包取包功能。 关键词:自动存包柜;单片机;随机密码
Design and Simulation of Automatic Lockers ABSTRACT This topic is microcontroller-based automatic lockers.Automatic lockers is a new generation of lockers, with a practical, simple operation, easy management, safe and reliable, able to better serve the broad masses of the different markets, users are based on a clear and concise instructions to complete the deposit bags to take the package. The system consists of MCS-51 microcontroller core control system, the entire system from the main section, the keyboard display control part of the implementation of some of the three-part composition, random password generation and check completed automatically save the package to take the package process. Various components of this design is easy to install and easy to operate, can basically save the package to take package function. Key words :Automatic lockers; microcontroller; random password
卫生部办公厅关于印发《脐带血造血干细胞治疗技术管理规 范(试行)》的通知 【法规类别】采供血机构和血液管理 【发文字号】卫办医政发[2009]189号 【失效依据】国家卫生计生委办公厅关于印发造血干细胞移植技术管理规范(2017年版)等15个“限制临床应用”医疗技术管理规范和质量控制指标的通知 【发布部门】卫生部(已撤销) 【发布日期】2009.11.13 【实施日期】2009.11.13 【时效性】失效 【效力级别】部门规范性文件 卫生部办公厅关于印发《脐带血造血干细胞治疗技术管理规范(试行)》的通知 (卫办医政发〔2009〕189号) 各省、自治区、直辖市卫生厅局,新疆生产建设兵团卫生局: 为贯彻落实《医疗技术临床应用管理办法》,做好脐带血造血干细胞治疗技术审核和临床应用管理,保障医疗质量和医疗安全,我部组织制定了《脐带血造血干细胞治疗技术管理规范(试行)》。现印发给你们,请遵照执行。 二〇〇九年十一月十三日
脐带血造血干细胞 治疗技术管理规范(试行) 为规范脐带血造血干细胞治疗技术的临床应用,保证医疗质量和医疗安全,制定本规范。本规范为技术审核机构对医疗机构申请临床应用脐带血造血干细胞治疗技术进行技术审核的依据,是医疗机构及其医师开展脐带血造血干细胞治疗技术的最低要求。 本治疗技术管理规范适用于脐带血造血干细胞移植技术。 一、医疗机构基本要求 (一)开展脐带血造血干细胞治疗技术的医疗机构应当与其功能、任务相适应,有合法脐带血造血干细胞来源。 (二)三级综合医院、血液病医院或儿童医院,具有卫生行政部门核准登记的血液内科或儿科专业诊疗科目。 1.三级综合医院血液内科开展成人脐带血造血干细胞治疗技术的,还应当具备以下条件: (1)近3年内独立开展脐带血造血干细胞和(或)同种异基因造血干细胞移植15例以上。 (2)有4张床位以上的百级层流病房,配备病人呼叫系统、心电监护仪、电动吸引器、供氧设施。 (3)开展儿童脐带血造血干细胞治疗技术的,还应至少有1名具有副主任医师以上专业技术职务任职资格的儿科医师。 2.三级综合医院儿科开展儿童脐带血造血干细胞治疗技术的,还应当具备以下条件:
大件寄存程序 一、大件寄存流程 1、当顾客提出大件寄存需求时,服务中心员工应先查看物品大小,如果可自助存包柜寄存的物品,请顾客优先使用自动存包柜,并提醒顾客不要寄放贵重物品。 2、如果物品过大,自助存包柜寄存不下的物品或顾客坚持要求寄存服务台的商品,服务台员工应与顾客确认是否有贵重物品,如有,请顾客将贵重物品取出自行保管,其它物品则办理寄存手续。 3、员工将顾客物品放置在指定区域,挂好存包牌,将另一块号码牌交予顾客,并告知顾客于当天营业结束前凭此号码牌领取寄存物品,请顾客妥善保管。 4、当顾客前来领取寄存物品时,服务中心员工请顾客出示号码牌,员工收回号码牌,根据号码牌查找物品上对应的号码牌,核对无误后交还顾客物品。存包牌两块互绑在一起放回存放盒内。 5、若核对号码牌时发现物品与存放时不符或丢失,应立即告知客服主管或值班经理前来处理(具体处理方法请参照客诉处理办法) 6、如顾客遗失号码牌时,服务中心员工处理程序: A、通知广播寻找遗失的号码牌。 B、询问顾客具体号码牌号或所寄存物品形状、颜色、特征。 C、询问顾客是否有任何的证件,并请顾客提供有效证件证明所寄存物品确为其所属。 D、请顾客在《寄存物异常登记表》上留下地址、电话、证件号等信息。 E、向顾客适当收取号码牌工本费并在退换货收银机做销售操作,将寄存物品归还于顾客。 7、每日营业结束后盘点号码牌,每块号码牌要求一个号码两块互绑在一起,如已不齐全的号码牌则不再使用。号码牌需定期全部更换。 二、大件寄存顾客告示 1、请您在存包后主动索取存包牌,并保存好自己的存包牌,不要遗失。 2、现金、手机、照相机、存折等超过200元的贵重物品及有效证件如身份证、户口本等物品请随身携带,请勿寄存,由此造成的损失,自行承担。 3、当您领取所存物品时,请出示存包牌,并当面点清您所存的物品,离柜本商场不再负责。 4、如存包牌丢失,应及时持有效证件领取所存物品,并付工本费5元,未及时挂失,导致所存物品丢失,不予负责。
文言实词词类活用 活用为一般动词 (一)名词活用为一般动词 1.两个名词连用,既不是并列关系,又不是修饰关系,便是动宾或主谓,其中一个必然活用为动词。 a .有一老父,衣褐,至良所。 b.籍吏民,封府库。 c.我有嘉宾,鼓瑟吹笙。 d.冬雷震震夏雨雪。 2.名词后紧跟代词,该名词活用为动词。 a.驴不胜怒,蹄之。 b.以其乃华山之阳名之。 c.名余曰正则兮。 3.名词放在副词后,便活用为动词。 a.日将暮,取儿槁葬。 b.太子及宾客知其事者,皆白衣冠以送之。 c.从弟子女十人所,皆衣缯单衣,立大巫后。 4.名词放在“能”“可”“足”“欲”等呢过愿动词后,便活用为动词。 a.假舟楫者,非能水也。 b.云青青兮欲雨。 c.其力尚足以入,火尚足以明。 d.子谓公冶长:“可妻也。” 5.名词带介宾结构做补语,这个名词活用为动词。 a.晋军(于)函陵,秦军(于)氾南。 b.唐浮图慧褒始舍于其址。 6.名词用“而”同动词或动宾词组连接时,活用为动词。 a.三代不同礼而王,五霸不同法而霸。 7.名词在“所”“者”结构中便活用为动词。 a.置人所罾鱼腹中。
a.是以,令吏人完客所馆。 形容词活用为一般动词 1.形容词用在“所”字之后,便活用为动词。 故俗之所贵,主之所贱;吏之所卑,法之所尊也。 (认为宝贵、认为低贱、认为卑下、认为高贵) 2.形容词在能愿动词后,活用为动词。 问其深,则其好游者不能穷也。(走到尽头) 3.形容词在“之”“我”能代词前,活用为动词。 稍出近之。(靠近) 4.形容词后带介宾结构做补语,它活用为动词。 令尹子兰……率使上官大夫短屈原于顷襄王。 (诋毁) 数词活用做一般动词 六王毕,四海一。(统一) 名词做状语 一、普通名词作状语 1.表比喻 a.嫂蛇行匍匐。 b.狐鸣呼曰。 c .赢粮而景从。 d .天下云集响应。 e.常以身翼蔽沛公。 f.一狼径去,其一犬坐于前。 2.表对人的态度 a.君为我呼入,吾得兄事之。 b.人人皆得以隶使之。 3.表动作行为的处所 a.夫以秦王之威,相如廷叱之,辱其群臣廷:在朝廷上 b.童子隅坐而执烛. 隅:在墙角 4.表动作行为的工具、凭借、方式
精神分裂症应该怎么治疗 1、坚持服药治疗 服药治疗是最有效的预防复发措施临床大量统计资料表明,大多数精神分裂症的复发与自行停药有关。坚持维持量服药的病人复发率为40%。而没坚持维持量服药者复发率高达80%。因此,病人和家属要高度重视维持治疗。 2、及时发现复发的先兆,及时处理 精神分裂症的复发是有先兆的,只要及时发现,及时调整药物和剂量,一般都能防止复发,常见的复发先兆为:病人无原因出现睡眠不好、懒散、不愿起床、发呆发愣、情绪不稳、无故发脾气、烦躁易怒、胡思乱想、说话离谱,或病中的想法又露头等。这时就应该及时就医,调整治疗病情波动时的及时处理可免于疾病的复发。 3、坚持定期门诊复查 一定要坚持定期到门诊复查,使医生连续地、动态地了解病情,使病人经常处于精神科医生的医疗监护之下,及时根据病情变化调整药量。通过复查也可使端正人及时得到咨询和心理治疗解除病人在生活、工作和药物治疗中的各种困惑,这对预防精神分裂症的复发也起着重要作用。 4、减少诱发因素 家属及周围人要充分认识到精神分裂症病人病后精神状态的薄弱性,帮助安排好日常的生活、工作、学习。经常与病人谈心,帮助病人正确对待疾病,正确对待现实生活,帮助病人提高心理承受能力,学会对待应激事件的方法,鼓励病人增强信心,指导病人充实生活,使病人在没有心理压力和精神困扰的环境中生活。 首先是性格上的改变,塬本活泼开朗爱玩的人,突然变得沉默寡言,独自发呆,不与人交往,爱干净的人也变的不注意卫生、生活
懒散、纪律松弛、做事注意力不集中,总是和患病之前的性格完全 相悖。 再者就是语言表达异常,在谈话中说一些无关的谈话内容,使人无法理解。连最简单的话语都无法准确称述,与之谈话完全感觉不 到重心。 第三个就是行为的异常,行为怪异让人无法理解,喜欢独处、不适意的追逐异性,不知廉耻,自语自笑、生活懒散、时常发呆、蒙 头大睡、四处乱跑,夜不归宿等。 还有情感上的变化,失去了以往的热情,开始变的冷淡、对亲人不关心、和友人疏远,对周围事情不感兴趣,一点消失都可大动干戈。 最后就是敏感多疑,对任何事情比较敏感,精神分裂症患者,总认为有人针对自己。甚至有时认为有人要害自己,从而不吃不喝。 但是也有的会出现难以入眠、容易被惊醒或睡眠不深,整晚做恶梦或者长睡不醒的现象。这些都有可能是患上了精神分裂症。 1.加强心理护理 心理护理是家庭护理中的重要方面,由于社会上普遍存在对精神病人的歧视和偏见,给病人造成很大的精神压力,常表现为自卑、 抑郁、绝望等,有的病人会因无法承受压力而自杀。家属应多给予 些爱心和理解,满足其心理需求,尽力消除病人的悲观情绪。病人 生活在家庭中,与亲人朝夕相处,接触密切,家属便于对病人的情感、行为进行细致的观察,病人的思想活动也易于向家属暴露。家 属应掌握适当的心理护理方法,随时对病人进行启发与帮助,启发 病人对病态的认识,帮助他们树立自信,以积极的心态好地回归社会。 2.重视服药的依从性 精神分裂症病人家庭护理的关键就在于要让病人按时按量吃药维持治疗。如果不按时服药,精神病尤其是精神分裂症的复发率很高。精神病人在医院经过一系统的治疗痊愈后,一般需要维持2~3年的
卫生部关于印发《脐带血造血干细胞库设置管理规范(试行)》的通知 发文机关:卫生部(已撤销) 发布日期: 2001.01.09 生效日期: 2001.02.01 时效性:现行有效 文号:卫医发(2001)10号 各省、自治区、直辖市卫生厅局: 为贯彻实施《脐带血造血干细胞库管理办法(试行)》,保证脐带血临床使用的安全、有效,我部制定了《脐带血造血干细胞库设计管理规范(试行)》。现印发给你们,请遵照执行。 附件:《脐带血造血干细胞库设置管理规范(试行)》 二○○一年一月九日 附件: 脐带血造血干细胞库设置管理规范(试行) 脐带血造血干细胞库的设置管理必须符合本规范的规定。 一、机构设置 (一)脐带血造血干细胞库(以下简称脐带血库)实行主任负责制。 (二)部门设置 脐带血库设置业务科室至少应涵盖以下功能:脐带血采运、处理、细胞培养、组织配型、微生物、深低温冻存及融化、脐带血档案资料及独立的质量管理部分。 二、人员要求
(一)脐带血库主任应具有医学高级职称。脐带血库可设副主任,应具有临床医学或生物学中、高级职称。 (二)各部门负责人员要求 1.负责脐带血采运的人员应具有医学中专以上学历,2年以上医护工作经验,经专业培训并考核合格者。 2.负责细胞培养、组织配型、微生物、深低温冻存及融化、质量保证的人员应具有医学或相关学科本科以上学历,4年以上专业工作经历,并具有丰富的相关专业技术经验和较高的业务指导水平。 3.负责档案资料的人员应具相关专业中专以上学历,具有计算机基础知识和一定的医学知识,熟悉脐带血库的生产全过程。 4.负责其它业务工作的人员应具有相关专业大学以上学历,熟悉相关业务,具有2年以上相关专业工作经验。 (三)各部门工作人员任职条件 1.脐带血采集人员为经过严格专业培训的护士或助产士职称以上卫生专业技术人员并经考核合格者。 2.脐带血处理技术人员为医学、生物学专业大专以上学历,经培训并考核合格者。 3.脐带血冻存技术人员为大专以上学历、经培训并考核合格者。 4.脐带血库实验室技术人员为相关专业大专以上学历,经培训并考核合格者。 三、建筑和设施 (一)脐带血库建筑选址应保证周围无污染源。 (二)脐带血库建筑设施应符合国家有关规定,总体结构与装修要符合抗震、消防、安全、合理、坚固的要求。 (三)脐带血库要布局合理,建筑面积应达到至少能够储存一万份脐带血的空间;并具有脐带血处理洁净室、深低温冻存室、组织配型室、细菌检测室、病毒检测室、造血干/祖细胞检测室、流式细胞仪室、档案资料室、收/发血室、消毒室等专业房。 (四)业务工作区域应与行政区域分开。