
华北农学报·2010,25(1):136-140
收稿日期:2009-08-08基金项目:教育部博士点基金资助项目(2000065402)作者简介:董桃杏(1982-),女,湖北黄冈人,硕士,主要从事生态学的研究。通讯作者:蔡昆争(1970-),男,云南曲靖人,副教授,博士,主要从事农业生态研究。
干旱胁迫下茉莉酸甲酯对水稻叶片质膜
透性及无机离子含量的影响
董桃杏
1,2
,蔡昆争1,曾任森
1
(1.华南农业大学农业部生态农业重点开放实验室,广东广州 510642;2.湖北第二师范学院化学与生命科学学院,湖北武汉 430205)
摘要:茉莉酸甲酯(M e J A )是植物体内一种具有应答外界刺激,传导逆境信号及启动抗逆基因的天然生理活性物质,提高作物的抗旱性是M e J A 的重要生理功能之一。研究了干旱胁迫下施用不同浓度(250,2.5,0.25μm o l /L )M e J A 对水稻幼苗的生理调控作用。结果表明,干旱处理后水稻叶片水势和叶绿素含量显著下降,叶片M D A 含量、电导率
及无机离子(K +,C a 2+,M g 2+
)含量均显著上升,而喷施不同浓度的M e J A 则显著提高叶片水势和叶绿素含量,降低质
膜透性和叶片无机离子含量,从而增强水稻幼苗的抗旱性。以0.25μm o l /LM e J A 处理抗旱效果最好,2.5μm o l /L 次之,然后是250μm o l /L 。
关键词:茉莉酸甲酯;干旱;水稻;抗旱性
中图分类号:Q 946.81 文献标识码:A 文章编号:1000-7091(2010)01-0136-05
E f f e c t s o f Me t h y l J a s m o n a t e o nMe m b r a n e P e r m e a b i l i t y a n d I n o r g a n i c
I o n s C o n t e n t i nR i c e L e a v e s u n d e r D r o u g h t S t r e s s
D O N GT a o -x i n g 1,2,C A I K u n -z h e n g 1,Z
E N GR e n -s e n
1
(1.K e y L a b o r a t o r y o f E c o l o g i c a l A g r i c u l t u r e o f M i n i s t r y o f A g r i c u l t u r e ,S o u t h C h i n a A g r i c u l t u r a l
U n i v e r s i t y ,G u a n g z h o u 510642,C h i n a ;2.C o l l e g e o f C h e m i s t r y a n d L i f e S c i e n c e ,H u b e i
U n i v e r s i t y o f E d u c a t i o n ,Wu h a n 430205,C h i n a )
A b s t r a c t :M e t h y l J a s m o n a t e (M e J A )i s a n i m p o r t a n t n a t u r a l l y p h y s i o l o g i c a l s u b s t a n c e t h a t r e g u l a t e s t h e e x p r e s -s i o n o f p l a n t d e f e n s e g e n e s i n r e s p o n s e t o v a r i o u s e n v i r o n m e n t a l s t r e s s i n c l u d i n g d r o u g h t .D i f f e r e n t c o n c e n t r a t i o n o f M e J At r e a t m e n t (0.25,2.5a n d 250μm o l /L )u n d e r d r o u g h t s t r e s s w e r e c h o s e n t o s t u d y i t s e f f e c t o n p h y s i o l o g i c a l t r a i t s o f r i c e s e e d l i n g s .U n d e r d r o u g h t s t r e s s c o n d i t i o n ,l e a f w a t e r p o t e n t i a l a n d c h l o r o p h y l l c o n t e n t w e r e s i g n i f i c a n t l y d e c r e a s e d ,w h e r e a s c o n d u c t i v i t y ,t h e c o n t e n t o f M D Aa n d i n o r g a n i c i o n s (K +
,C a 2+
,M g 2+
)i n r i c e l e a v e s w e r e s i g -n i f i c a n t l y i n c r e a s e d ,t h e s e e f f e c t s c o u l d b e r e s t r a i n e da f t e r s p r a y i n g v a r i o u s c o n c e n t r a t i o no f M e J A (250,2.5,0.25μm o l /L ),w h i c h m e a n t M e J Ac o u l d i n c r e a s e d r o u g h t r e s i s t a n c e .T h e r e e x i s t e dc o n c e n t r a t i o ne f f e c t ,0.25μm o l /L M e J Ah a d t h e b e s t e f f e c t ,2.5μm o l /Lt o o k s e c o n d p l a c e ,t h e t h i r d w a s 250μm o l /L .
K e y w o r d s :M e t h y l J a s m o n a t e ;D r o u g h t ;R i c e ;D r o u g h t r e s i s t a n c e 以茉莉酸(J a s m o n i ca c i d ,J A )和茉莉酸甲酯(M e t h y l e J a s m o n a t e ,M e J A )为代表的茉莉酸类物质(J a s m o n a t e s ,J A s )在自然界中广泛存在,对调节植物生长发育、光合特性、抗逆反应等起重要作用。大量研究表明,茉莉酸类物质在环境胁迫反应中是驱动植物防御基因表达和信号传递的重要组成部
分[1-3]
,它能作为内源信号分子参与植物在生物逆境(病害和虫害)和非生物逆境胁迫(干旱、低温、盐胁迫等)中的抗性作用[4-8]
。M e J A 是植物体内一种具有应答外界刺激,传导逆境信号及启动抗逆基因的天然生理活性物质,提高作物的抗旱性是M e J A 的重要生理功能之一。潘瑞炽等[9]
用M e J A 处理花生幼苗之后,其在形态解剖和生理上都发生显著变
1期董桃杏等:干旱胁迫下茉莉酸甲酯对水稻叶片质膜透性及无机离子含量的影响
137
化,由于减少了水分损失,从而加大了叶片中水分的
贮存,提高了幼苗的抗旱性。姚允聪等[10]
用30,70m g /L 的M e J A 处理中度干旱条件下的苹果(M a l u s s i e v e r s i i )幼树,发现能明显增加茎粗,促进地上部和根的干物质积累,显著提高叶片相对含水量(R W C )、游离脯氨酸含量、光合速率及S O D 活性,降低质膜透性、M D A 含量和叶细胞饱和脂肪酸含量,提高不饱和脂肪酸含量和不饱和脂肪酸指数,从而在一定程度上提高了苹果幼树的抗旱性。干旱条件下大豆植株施加J A 后其鲜质量因干旱减少15%,叶片中的J A 水平在2h 内增加约5倍,这个
结果说明J A 在干旱情况下发挥着重要的作用[4]
。在干旱条件下,两种基因型的大麦种子经5,15μm o l /L 的J A 处理24h 后,较低浓度的J A 明显减小干旱诱导的膜伤害[11]
。本研究的目的是通过不同浓度的M e J A 处理对干旱胁迫下水稻幼苗的叶片水势、质膜透性、脂氧合酶(L O X )活性、无机离子(K +
、C a 2+
、M g 2+
)含量的影响,揭示其抗旱生理效应。
1 材料和方法
1.1 试验设计及处理
选用的水稻品种为中二欧6。用0.1%H g C l 2消毒种子30m i n ,浸种24h ,25℃催芽2d 。将已催芽的种子置于装有蛭石的塑料杯中,加入200m L 营养液,放在光照培养箱中,进行培养。培养箱条件
为:光照14h /d ,光强200μm o l /(m 2
·s ),昼夜温度为28℃/24℃,相对湿度为80%,培养40d 后,进行5种处理:①对照;②干旱处理(溶液水势保持在-0.8M P a );③干旱+250μm o l /L M e J A 处理;④干旱+2.5μm o l /LM e J A 处理;⑤干旱+0.25μm o l /L M e J A 处理。每个处理设5个重复。采用20%的P E G 6000模拟干旱处理,此时溶液水势保持在-0.8M P a 左右。处理2d 后采样,选取第2,3,4片叶作为试验材料,测定叶片水势,丙二醛(M D A )含量,电导率,叶绿素含量及无机离子(K +
,C a 2+
,M g 2+
)含量。
1.2 测定指标及方法
叶片水势采用美国Y a m a t o 公司生产的W P 4水势仪进行测定;叶绿素含量采用日本生产S P A D -502型叶绿素计测定;质膜相对透性用电导仪进行测定,丙二醛采用比色法测定
[12]
。
无机渗透调节物质:称取0.3g 鲜叶烘干磨碎样品于消化管中,加入5m L 高氯酸和15m L 浓硝酸
浸泡静置过夜,然后在硝化炉上于180℃下消煮至
透明色,冷却后用去离子水定容至50m L ,用V u r i a n
公司(美国)生产的S p e c t r A A 220F S 火焰原子吸收光谱仪测定C a 2+
,M g 2+
,K +
含量。
2 结果与分析
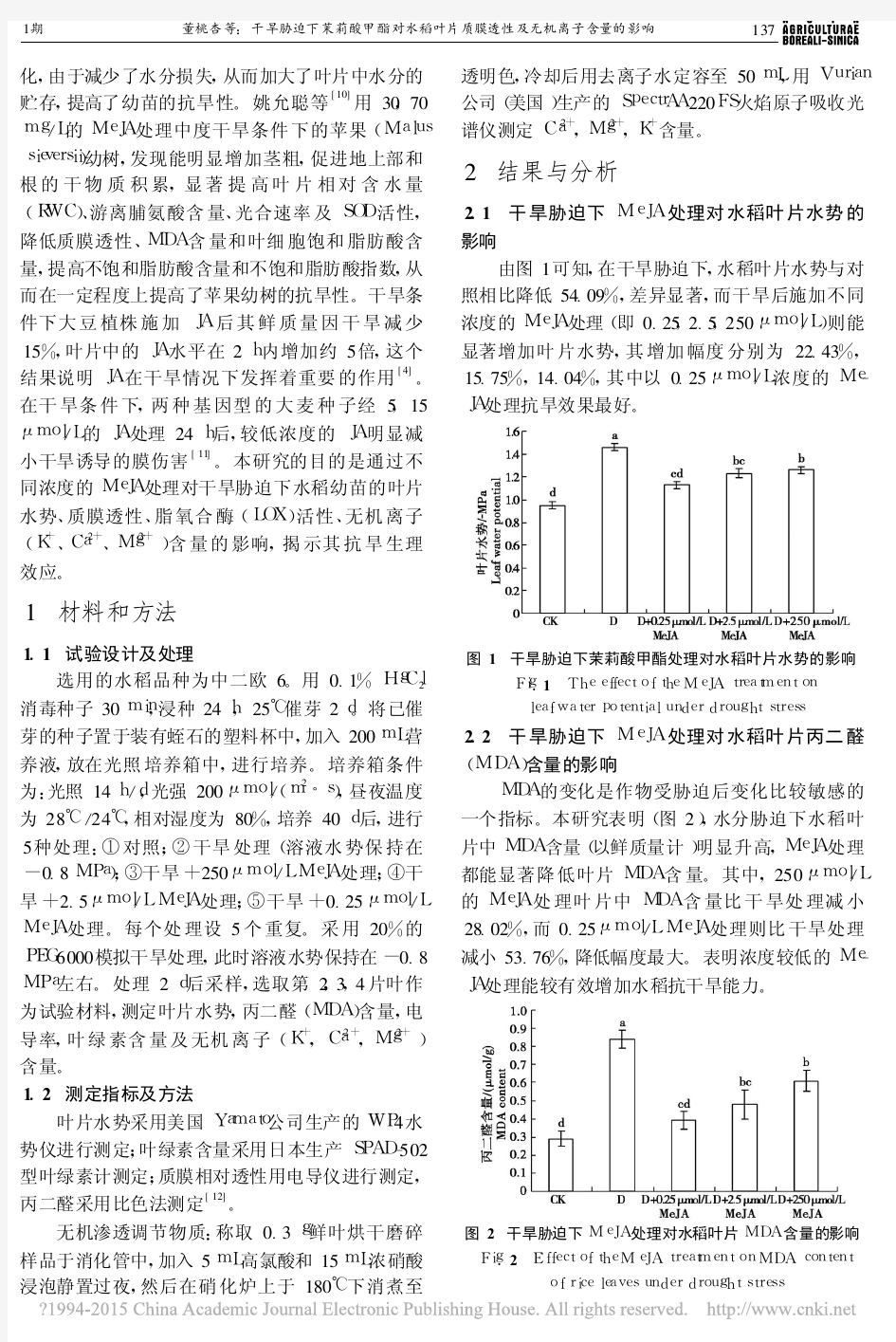
2.1 干旱胁迫下M e J A 处理对水稻叶片水势的影响
由图1可知,在干旱胁迫下,水稻叶片水势与对照相比降低54.09%,差异显著,而干旱后施加不同浓度的M e J A 处理(即0.25,2.5,250μm o l /L )则能显著增加叶片水势,其增加幅度分别为22.43%,15.75%,14.04%,其中以0.25μm o l /L 浓度的M e -J A 处理抗旱效果最好
。
图1 干旱胁迫下茉莉酸甲酯处理对水稻叶片水势的影响
F i g .1 T h e e f f e c t o f t h eMe J A t r e a t m e n t o n l e a f w a t e rp o t e n t i a l u n d e r d r o u g h t s t r e s s
2.2 干旱胁迫下M e J A 处理对水稻叶片丙二醛(MD A )含量的影响
M D A 的变化是作物受胁迫后变化比较敏感的一个指标。本研究表明(图2),水分胁迫下水稻叶片中M D A 含量(以鲜质量计)明显升高,M e J A 处理都能显著降低叶片M D A 含量。其中,250μm o l /L 的M e J A 处理叶片中M D A 含量比干旱处理减小28.02%,而0.25μm o l /LM e J A 处理则比干旱处理减小53.76%,降低幅度最大。表明浓度较低的M e -J A 处理能较有效增加水稻抗干旱能力。
图2 干旱胁迫下Me J A 处理对水稻叶片MD A 含量的影响F i g .2 E f f e c t o f t h e Me J At r e a t m e n t o nMD A c o n t e n t
o f r i c e l e a v e s u n d e r d r o u g h t s t r e s s
138
华 北 农 学 报25卷2.3 干旱胁迫下Me J A处理对水稻叶片电导率的
影响
电导率也可以用伤害率表示,是目前鉴定植物
抗逆性强弱的一个精确而实用的方法,现在已广泛
应用于作物抗性栽培育种。本研究表明,干旱处理
使水稻叶片电导率急剧增加,比例达到25.17%。
加入M e J A后显著降低水稻叶片的电导率,0.25,
2.5及250μm o l/L浓度的M e J A处理与干旱处理相
比分别降低16.62%,11.31%,6.61%。其中0.25
μm o l/L浓度的M e J A处理降低幅度最大(图3)
。
图3 干旱胁迫下Me J A处理对水稻叶片电导率的影响
F i g.3 T h e e f f e c t o f t h e Me J A t r e a t m e n t o nl e a f
c o n
d u c t i v i t y l
e a v e s u n d e r d r o u g h t s t r e s s
2.4 干旱胁迫下Me J A处理对水稻叶片叶绿素含
量的影响
水分胁迫对水稻叶片叶绿素含量也具有明显影
响(图4)。遭受水分胁迫后,水稻叶片叶绿素含量
显著降低,比对照减小21.99%,0.25,2.5和250
μm o l/L 3个浓度的M e J A处理后使叶绿素含量分
别比干旱处理增加17.00%,10.87%和3.68%,其
中,浓度最小的处理(0.25μm o l/L)增加幅度最大
。
图4 干旱胁迫下Me J A处理对水稻叶片叶绿素含量的影响
F i g.4 T h e e f f e c t o f t h e Me J A t r e a t m e n t o nl e a f
c h l o r o p h y l l c o n t e n t u n
d
e rd r o u g h t s t r e s s c h l o r o p h y l l
2.5 干旱胁迫下Me J A处理对水稻叶片无机离子
含量的影响
由图5可知,在干旱胁迫条件下,水稻叶片中钙
离子含量显著增加,与对照相比增加49.87%。干
旱后经过不同浓度的M e J A处理则叶片中钙离子含
量显著下降。其中,0.25,2.5和250μm o l/L的处
理使叶片中钙离子含量相对于单独干旱处理分别降
低29.91%,24.40%和16.21%。
图5 干旱胁迫下Me J A处理对水稻叶片C a2+含量的影响
F i g.5 T h e e f f e c t o f t h e Me J At r e a t me n t o nC a2+c o n t e n t
o f r i c e l e a v e s u n d e r d r o u g h t s t r e s s c h l o r o p h y l l
M e J A处理对镁离子(图6)及钾离子(图7)的
影响与对钙离子的影响也具有相似效应,即干旱胁
迫无机离子含量显著增加,而不同浓度的M e J A处
理则由于缓解干旱胁迫,从而导致离子含量显著下
降,恢复到与对照接近的水平。
图6 干旱胁迫下Me J A处理对叶片Mg2+含量的影响
F i g.6 T h e e f f e c t o f t h e Me J At r e a t m e n t o nMg2+
c o n t e n t o f r i c el e a v e s u n
d
e rd r o u g h t s t r e s s
图7 干旱胁迫下Me J A处理对叶片K+含量的影响
F i g.7 T h e e f f e c t o f t h e Me J At r e a t me n t o nK+
c o n t e n t o f r i c el e a v e s u n
d
e rd r o u g h t s t r e s s
3 讨论
本研究表明,在干旱条件下施加不同浓度的
M e J A均能显著提高水稻幼苗的抗旱性,表现在叶片
水势提高,电导率和丙二醛含量显著下降,而以0.25
1期董桃杏等:干旱胁迫下茉莉酸甲酯对水稻叶片质膜透性及无机离子含量的影响
139
μm o l /LM e J A 处理效果最好。这说明M e J A 处理能
改善叶片的水分状况,降低干旱造成的膜脂过氧化程度,减少细胞膜所受到的伤害。植物细胞对维持细胞的微环境和正常的代谢起着重要的作用,质膜包围在细胞原生质体表面,通常被认为是干旱胁迫伤害的最初和关键部位[13]
。当植物受到逆境影响时,如高温或低温,干旱,盐渍,病原菌浸染后,细胞膜遭到破坏,膜透性增大,从而使细胞内的电解质外渗,以致植物细胞浸提液的电导率增大。膜透性增大的程度与逆境胁迫强度有关,也与植物抗逆性的强弱有关
[14]
。B a n d u r d k a 等
[11]
利用两种基因型的
大麦作为试验材料,在干旱条件下施加茉莉酸后发
现能显著降低大麦细胞膜受的伤害。陈培琴等[15]
发现水分胁迫下电解质渗透率升高,这是膜透性增大的缘故,经J A 处理葡萄幼苗在一定程度上缓解了膜损伤程度,提高了葡萄幼苗的耐热性能。本研究表明,在干旱胁迫下水稻叶片M D A 含量显著增加,施加M e J A 后由于减少了细胞的膜脂过氧化作用,从而减轻植物细胞膜受到伤害的程度。有关研究也表明,M e J A 增强水稻幼苗的抗旱性与水稻幼苗自身清除活性氧的能力有关
[16]
。
渗透调节是植物适应干旱的一种重要生理机制。在干旱胁迫条件下植物体内的有机(如脯氨酸、可溶性糖、游离氨基酸等)和无机渗透调节物质(如K +
、C a 2+
、M g 2+
等)会大量积累。李春香等
[17]
通过小麦试验表明随着水分胁迫的加重,叶片中无机渗透调节物质(K +
、C a 2+
、M g 2+
)的浓度均有增加的趋势,不同生育期及不同品种间差异不显著。蔡昆争等
[18,19]
研究表明,不同生育期干旱胁迫后水稻
根系和叶片的无机渗透调节物质(K +
、C a 2+
和M g 2+
)的含量均大幅度上升,干旱越严重,上升幅度越大,反映了作物通过渗透调节适应干旱的生理机
制。C a 2+
离子作为植物细胞的第二信使传递干旱信号,调节干旱胁迫导致的生理反应
[20]
。有研究表明
干旱抑制根系对钙的吸收,而钙能提高植株的抗旱性,抑制干旱胁迫下活性氧物质的生成,保护细胞质膜的结构,维持正常的光合作用,以及调节激素和一些重要的生化物质代谢
[21,22]
。目前关于K +,M g
2+
等无机渗透调节物质的报道比较少,关军峰等[20]
研
究表明,K +
对抗氧化酶活性具有促进作用。N i c o l a
等
[23]
通过细胞膜片钳试验,以蚕豆(V i c i a f a b a )为
材料,证实M e J A 对保卫细胞K +
通道有浓度效应,通过调节原生质膜K +
的流量来促进气孔关闭。本研究表明,干旱条件下叶片中K +
,C a 2+
,M g 2+
含量大幅度上升,而不同浓度的M e J A 处理均显著降低
了这些无机离子的浓度,这说明茉莉酸甲酯处理后缓解了干旱胁迫的压力,改善了植株体内的水分状况,因此无机离子的含量因而相对干旱来说就显著降低。参考文献:
[1] H a r m sK ,A t z o r nR ,B r a s hA ,e t a l .E x p r e s s i o no f a f l a x
a l l e n eo x i d es y n t h a s ec D N A l e a d s t oi n c r e a s e de n d o g e -n o u s j a s m o n i c a c i dl e v e l s i nt r a n s g e n i cp o t a t o p l a n t s
b u t n o t t oa
c o r r e s p o n
d i n ga c t i v a t i o no f J A -r
e s p o n d i n gg e n e s [J ].P l a n t C e l l ,1995,7:1645-1654.
[2] L e h m a n nJ ,A t z o r nR ,B r u c k n e r C ,e t a l .A c c u m u l a t i o no f
j a s m o n a t e ,a b s c i s i ca c i d ,s p e c i f i ct r a n s c r i p t s a n dp r o t e i n s i no s m o t i c a l l y s t r e s s e db a r l e yl e a f s e g m e n t s [J ].P l a n t a ,1995,197:156-162.
[3] 蔡昆争,董桃杏,徐 涛.茉莉酸类物质(J A s )的生理
特性及其在逆境胁迫中的抗性作用[J ].生态环境,2006,15(2):397-404.
[4] C r e e l m n RA ,M u l e n t J E .J a s m o n i c a c i dd i s t r i b u t i o n a n d
a c t i o ni np l a n t s :R e g u l a t i o nd u r i n gd e v e l o p m e n t a n dr e -s p o n s e t o
b i o t i
c a n da b i o t i c s t r e s s [J ].P r o c e e
d i n g s o f t h
e N a t i o n a l A c a d e m y o
f S c i e n c e s o f t h e U S A ,1995,92:4114-4119.
[5] M o o n s A ,P r i n s e n E ,B a u wG ,e t a l .A n t a g o n i s t i c e f f e c t s o f
a b s c i s i c a c i d a n dj a s m o n a t e s o n s a l t s t r e s s -i n d u c i b l e t r a n -s c r i p t s i nr i c e r o o t s [J ].T h e P l a n t C e l l ,1997,9:2243-2259.
[6] G a oXP ,Wa n gXF ,L uY F ,e t a l .J a s m o n i c a c i di s i n -v o l v e d i n t h e w a t e r -s t r e s s -i n d u c e d b e t a i n e a c c u m u l a t i o n i n p e a r l e a v e s [J ].P l a n t C e l l a n dE n v i r o n m e n t ,2004,27:497-507.
[7] G o n z a l i e zA ,T i z n a d oH M ,Z a v a l e t aG R ,e t a l .M e t h y l
j a s m o n a t e t r e a t m e n t s r e d u c ec h i l l i n gi n j u r ya n da c t i v a t e t h e d e f e n s e r e s p o n s e o f g u a v a f r u i t s [J ].B i o c h e m i c a l a n d B i o p h y s i c a l R e s e a r c hC o m m u n i c a t i o n s ,2004,313:694-701.
[8] T h a l e rJ S ,O w e nB ,H i g g n sV J .T h eR o l eo f t h eJ a s -m o n a t e r e s p o n s ei np l a n t s u s c e p t i b i l i t yt od i v e r s ep a t h o -g e n s w i t har a n g eo fl i f e s t y l e s[J ].P l a n tP h y s i o l o g y ,2004,135(1):530-538.
[9] 潘瑞炽,古焕庆.茉莉酸甲酯对花生幼苗生长合抗旱
性的影响[J ].植物学报,1998,40(3):256-262.[10] 姚允聪,张大鹏,王有年.M J 提高苹果幼树抗旱性的
研究初报[J ].园艺学报,1999,26(5):333-334.
[11] B a n d u r s k a H ,S t r o n s k i T r o i c s k i A ,J a nK .T h ee f f e c t o f
j a s m o n a t eo nt h ea c c u m u l a t i o no f A B A ,p r o l i n ea n di t s i n f l u e n c e o nm e m b r a n ei n j u r yu n d e r w a t e r d e f i c i e n t i n t w ob a r l e yg e n o t y p e s [J ].A C T A P h y s i o l o g i a eP l a n t a -r u m ,2003,25(3):279-285.
140
华 北 农 学 报25卷
[12] 李合生.植物生理生化实验原理与技术[M].北京:
高等教育出版社,2000.
[13] 王洪春.植物抗逆生理[J].植物生理学通讯,1981,
(6):72-81.
[14] 陈少裕.膜脂过氧化与植物逆境胁迫[J].植物学通
报,1989,6(4):211-217.
[15] 陈培琴,郁松林,詹妍妮,等.茉莉酸对葡萄幼苗耐热
性的影响[J].石河子大学学报,2006,24(1):87-
91.
[16] 董桃杏,蔡昆争,张景欣,等.茉莉酸甲酯对水稻幼苗
的抗旱生理效应[J].生态环境,2007,16(4):1261-
1265.
[17] 李春香,王 玮,李德全.长期水分胁迫对小麦生育
中后期根叶渗透调节物质的影响[J].西北植物学
报.2001,21(5):924-930.
[18] 蔡昆争,吴学祝,骆世明.不同生育期水分胁迫对水
稻根叶渗透调节物质变化的影响[J].植物生态学
报,2008,32(2):491-500.
[19] 蔡昆争,吴学祝,骆世明.抽穗期不同程度水分胁迫
对水稻产量和根叶渗透调节物质的影响[J].生态学
报,2008,28(12):6148-6158.
[20] 关军锋,李广敏.C a2+与植物抗旱性的关系[J].植物
学通报,2001,18(4):473-478.
[21] 蒋明义,郭少川.水分亏缺诱导的氧化胁迫和植物的
抗氧化作用[J].植物生理学通讯,1996,32(2):17-
23.
[22] 宗 会.干旱胁迫下C a2+·C a M信使系统对稻苗保护
酶活性的影响[J].西北植物学报,2004,24(5):843
-849.
[23] N i c o l a H,E v a n s.M o d u l a t i o n o f g u a r dc e l l p l a s m a m e m-
b r a n e p o t a s s i u m
c u r r e n t s b y m e t h y l j a s m o n a t e[J].P l a n t
P h y s i o l o g y,2003,131:8-11.
寒害生理与植物抗冷性 摘要:植物在长期进化过程中,形成了各种适应冬季低温的生长习性。寒害指由低温引起植物伤害的现象,包括冷害和冻害。植物对低温的适应性和抵抗能力称为抗寒性。 关键词:冷害冻害抗寒性冷驯化 1. 冷害生理与植物抗冷性 1.1 冷害、抗冷性的概念及分类 零上低温时,虽无结冰现象,但能引起喜温植物的生理障碍,是植物受伤甚至死亡,0℃以上低温对植物造成的危害称为冷害(chilling injury)。而植物对零上低温的适应能力称为抗冷性(chilling resistance)。在我国,冷害常发生于早春和晚秋季节,主要危害发生在作物的苗期和子粒或果实成熟期,处于开花期的果树遇冷害是会引起大量落花,使结实率降低。根据植物对冷害的反应速度,冷害分为:一、直接伤害。植物受低温影响数小时,最多在一天内即出现伤斑及坏死,直接破坏了原生质活性。二、间接伤害。植物受低温危害后,短时间无异常表现,至少在几天后才出现组织柔软、萎焉,因低温引起代谢失常而造成细胞伤害。 1.2 影响冷害因素 冷害对植物的伤害不仅与低温的程度和持续时间直接有关,还与植物组织的生理年龄、生理状况及对冷害的敏感性有关。温度低,持续时间长,植物受害严重,反之则轻。在同等冷害条件下,幼嫩组织和器官比老的组织和器官受害严重;同一植株生殖生长期比营养生长期对冷害敏感,其中花粉母细胞减数分裂期前后最敏感。 1.3 冷害机制
冷害对植物的伤害大致分为两个步骤:第一步是膜相改变,第二步是由于膜损坏而引起代谢紊乱,导致死亡。 1)膜脂发生相变。在低温冷害下,生物膜的脂类由液晶态变化凝胶态,从而引起与膜相结合的酶解离或使酶亚基分解失去活性。因为酶蛋白质是通过疏水键与膜脂相结合的,而低温使二者结合脆弱,易于分离。相变温度随脂肪酸链的长度而增加,而随不饱和脂肪酸所占比例增加而降低。温带植物比热带植物耐低温的原因之一是构成膜脂不饱和脂肪酸的含量较高。膜不饱和脂肪酸指数,即不饱和脂肪酸在总脂肪酸中的相对比值,可成为衡量植物抗冷性的重要生理指标。 2)膜的结构改变。在缓慢降温条件下,由于膜脂的固化使得膜结构紧缩,降低了膜对水和溶质的透性;在寒流突然来临的情况下,由于膜体紧缩不匀而出现断裂,因而会造成膜的破损渗漏,胞内溶质外流。 3)代谢紊乱。低温使得生物膜结构发生显著变化,进而导致植物体内新陈代谢的有序性被打破,特别是光合与呼吸速率改变,植物处于饥饿状态,而且还积累有毒的中间物质。 1.4 提高植物抗冷措施及原理 ○1低温锻炼。例如,春季采用温室、温床育苗,在露天移栽前,先降低室温或床温至10℃左右保持1~2天,移入大田后即可抵抗3~5℃的低温。凡经过低温锻炼的植物,膜不饱和脂肪酸含量增加,变相温度降低,透性稳定;细胞内[NADPH]/[NADP+]增高,A TP增加,这些都有助于抗冷性的形成与增强。 ○2化学诱导。使用ABA、CTK、2,4-D、油菜素内酯等提高植物抗冷性。 ○3合理施肥。在低温到来之前,合理调整施肥种类,适当增施磷钾肥,少施或不施速效氮肥。 ○4选育抗冷品种。通过基因工程、细胞工程及杂交育种技术选育抗冷性强的新品种。 2. 冻害与植物抗冻性 2.1 冻害概念、类型及危害 冰点以下的低温使植物组织内结冰引起的伤害称冻害。植物对零下低温的适应能力称抗冻性。植物受冻害时,叶片犹如烫伤,细胞失去膨压,组织变软,叶片颜色变为褐色,严重时导致死亡。主要有两种类型:一是胞外结冰。当环境温度缓慢降低,植物组织内温度降到冰点以下时,细胞间隙的水分开始结冰。大多数植物胞间结冰后,经缓慢解冻后仍能恢复正常生长。二是胞内结冰。当环境温度骤降,植物组织同时还会发生胞内结冰,一般先在原生质内结冰,后在液泡内结冰。胞内结冰一般不发生,一旦发生植物很难存活。
拟南芥 中文学名:阿拉伯芥(拟南芥) 拉丁学名:Arabidopsis thaliana 别称:阿拉伯鼠耳芥、拟南芥、鼠耳芥、阿拉伯草 界:植物界 门:被子植物门 纲:双子叶植物纲 目:十字花目 科:十字花科 属:鼠耳芥属(拟南芥属) 英文名称:Thale Cress 个体特征 阿拉伯芥是一种十字花科植物,广泛用于植物遗传学、发育生物学和分子生物学的研究,已成为一种典型的模式植物,其原因主要基于该植物具有以下特点:阿拉伯芥1 (1)植株形态个体小,高度只有30cm左右,1个茶杯可种植好几棵;(2)生长周期快,每代时间短,从播种到收获种子一般只需6周左右;(3)种子多,每株每代可产生数千粒种子;(4)形态特征简单,生命力强,用普通培养基就可作人工培养;(5)基因组小,只有5对染色体。虽然阿拉伯芥在许多方面“简单”,但它的大多数基因与其他“复杂”的植物基因具有很高的同源性,另外,由于这种植物的全部基因组测序已经完成,因此可以预测,阿拉伯芥在植物学所有领域的研究中将发挥更大的作用。阿拉伯芥2 阿拉伯芥的基因组是目前已知植物基因组中最小的。每个单倍染色体组(n=5)的总长只有7000万个碱基对,即只有小麦染色体组长的1/80,这就使克隆它的有关基因相对说来比较容易。阿拉伯芥是自花受粉植物,基因高度纯合,用理化因素处理突变率很高,容易获得各种代谢功能的缺陷型。例如用含杀草剂的培养基来筛选,一般获得抗杀草剂的突变率是1/100000。由于有上述这些优点,所以阿拉伯芥是进行遗传学研究的好材料,被科学家誉为“植物中的果蝇”。 繁殖培育 概述 阿拉伯芥 在一般的温室或人工气候室条件下,从阿拉伯芥种子的春化至第一批角果成熟大约需8周左右时间。当然,也可以通过改变生长条件以达到使阿拉伯芥提前或推后开花结实的目的。如延长每天的光照时间,可使阿拉伯芥明显地提前开花结实,利用每天接近24小时的光照条件培养,甚至在6周左右即可收获第一批成熟角果。阿拉伯芥的这一特性使实验工作周期大大缩短,特别是对于许多遗传分析工作,比利用一般的高等植物材料(如麦类、豆类作物)可以成倍地节约时间。阿拉伯芥是野生的双子叶草本植物,在野外自然条件下一般春季萌发,到夏季即种子成熟。应该说阿拉伯芥的生长发育对环境条件并无十分特殊的要求,因此在实验室条件下的种植及管理也相对比较简单。但中国许多实验室反映,阿拉伯芥的培养较为困难,幼苗时发生死亡、或抽薹后植株生长瘦弱等现象经常发生;而且经常发生较严重的虫害等。
植物学通报1997,14(2):1~8 Ch inese Bulletin of Bo tany 植物抗寒机理研究进展Ξ 沈 漫 王明庥 黄敏仁 (南京林业大学林木遗传和基因工程重点实验室,南京210037) 摘 要 本文综合概述了国内外有关植物抗寒机理研究的动态,主要讨论了植物抗寒性与细 胞膜系、酶系多态性及抗寒基因表达与调控之间的相关性。此外,亦提出了有关植物抗寒机制 研究领域值得深入研讨的问题。 关键词 植物抗寒性;细胞膜系;酶系;抗寒基因 AD VANCES IN RESEARCH ON CH I LL ING- RESISTANCE M ECHAN IS M S OF PLANTS Shen M an W ang M ing2x iu H uang M in2ren (F orest T ree and Genetic E ng ineering Op en ing L abora tory,N anj ing F orestry U n iversity,N an jing210037) Abstract T h is paper gives a general statem en t abou t the p resen t developm en t of ch illing2re2 sistan t m echan is m s of p lan ts at hom e and ab road,and it deals m ain ly w ith the relati on s be2 tw een the ch illing2resistance of p lan ts and cell m em b rane system,enzym e system diverisity and ch illing2resistan t gene exp ressi on and con tro l.In additi on,the paper po in ts ou t som e p rob lem s w o rth delib rating deep ly in the research field of p lan t ch illing2resistance. Key words Ch illing2resistance of p lan ts,Cell m em b rane system,Enzym e system diversity, Ch illing2resistan t gene 植物对环境变迁及不良环境有足够的适应性和抗抵能力,这种抗逆性既受系统发育的遗传基因所控制,又受个体发育中生理生态所制约。温度作为重要的环境因子之一,在植物遗传背景限制的前提下,对植物某些生长发育过程起着决定性作用。至今低温寒害对植物的伤害还没有找到根本的解决途径。因此,探索植物抗寒性的生理机制及其遗传因素,不仅在基础理论上具有重要意义,在解决生产实际问题上也具有广泛的应用价植。 近年来,国内外从细胞和分子生物学方面来研究植物的抗寒性,取得了一些重要结果,似乎找到了深入研究的突破口。为便于对植物低温反应和抗寒机制有一个较全面的了解,本文对这一领域的研究概况和进展作一个综述。 Ξ“八五”国家科技攻关课题《美洲黑扬胶合板林纸浆材新品种选育研究》一部分。
大同市常见四种绿化灌木抗寒性比较 摘要:植物的正常生长发育需要一个适宜的温度环境,低于一定的温度,植物就会受到低温的胁迫或者冻害,甚至引起植株死亡。但在低温的胁迫下,植物体内也会发生一系列的生理生化反应来消除或降低低温的伤害作用。本文综述了寒害和冻害发生的生理机理,从生理指标方面论述了植物与抗寒性的关系、各项生理指标的测定方法以及对未来研究的展望。 关键词:抗寒性、生理指标 1 绿化灌木简介 水蜡、紫叶小檗、小叶黄杨、小叶丁香是大同市绿化中常见的四种灌木。 水蜡是木犀科女贞属的植物,是一种落叶灌木,在北方各地广泛栽培,适应性较强,喜光照,稍耐荫,耐寒,对土壤要求不严格,适合做盆景或造型树等。 紫叶小檗是小檗科小檗属的植物,落叶灌木,适应性强,喜阳,耐半阴,但在光线稍差或密度过大时部分叶片会返绿。耐寒,但不畏炎热高温,耐修剪。园林常用与常绿树种作块面色彩布置,可用来布置花坛、花镜,是园林绿化中色块组合的重要树种。 小叶黄杨是黄杨科黄杨属的植物,常绿灌木,我国各地均有分布,小叶黄杨属于生长缓慢的植物,其耐寒性很弱,在零下10摄氏度,既能冻伤。而且很怕水淹,但是耐旱性很好,生长浅根性,根系密集发达。养护管理简单方便,寿命漫长,通过不断的引种,现在在北方北京等地方冬天也能很好的生活。 小叶丁香是木犀科丁香属的植物,落叶灌木,主要分布于中国河北、河南、山西、陕西等省,喜充足阳光,也耐半荫。适应性较强,耐寒、耐旱、耐瘠薄,病虫害较少。以排水良好、疏松的中性土壤为宜,忌酸性土。忌积涝、湿热。 2 植物抗寒性研究 2.1植物寒害和冻害的生理机理 2.1.1寒害机理 喜温植物在零下低温条件下,原生质流动减慢或停止,水分平衡失调,光合速率减弱,呼吸速率大起大落。零下低温对组织的伤害,大致分为两个步骤:第一步是膜相的改变,第二步是由于膜损坏而引起的代谢紊乱,导致死亡。抗寒性弱的植物,由于生物膜的不饱和脂肪酸含量少,膜的液化程度较差,伸缩性小,在低温来临的时候,膜从液晶态转变为凝胶态,膜收缩,出现裂缝。
冠菌素和茉莉酸甲酯对玉米种子萌发和幼苗生长的影响 摘要玉米种子经COR和MeJA处理后发现,低浓度的COR(0.01μmol/L)和MeJA(1.0μmol/L)促进种子萌发;提高了种子中可溶性糖含量、α-淀粉酶活性、增加侧根数和株高,而高浓度COR(1.0μmol/L)和MeJA(100.0μmol/L)则相反;COR和MeJA诱导的根系表面积的增加主要是由于须根数增加导致的;在促进种子萌发方面,COR的活性比MeJA高,大约是MeJA的100倍,初步认为低浓度COR促进种子萌发与诱导的α-淀粉酶有关。 关键词冠菌素(COR);茉莉酸甲酯(MeJA);玉米;萌发 玉米是重要的粮食作物、饲料作物和能源作物。培育齐苗壮苗是玉米田间生产的重要目标之一。实际生产中,一些玉米种子因为仓储老化或者萌发环境恶劣导致出苗率低、幼苗不健壮,还有一些基因型玉米种子田间制种过程中也出现类似的问题,严重影响繁殖制种速度及田间生产应用[1]。 利用植物生长调节物质处理种子,可以促进或抑制种子的生长发育,使生长发育发生质的变化[2]。玉米是使用化控调节剂较多的作物品种之一。一些研究发现[3-4],采用化学调控物质如赤霉素(GA)、6-苄基氨基嘌呤(6-BA)处理种子,可以打破玉米种子休眠,促进发育、发根,培育壮苗,提高幼苗的抗逆性。 冠菌素(Coronatine,COR)是丁香假单胞菌属的代谢产物[5],研究发现,COR能影响植物生长发育、诱导防御相关的次生物质代谢和调节逆境胁迫等[6],COR和茉莉酸/茉莉酸甲酯(JA/MeJA)都有环五烷结构,两者的许多生理功能相似[7-10]。有很多报道涉及MeJA调节水稻和花生种子的萌发[11-12],但尚未见COR和MeJA对玉米种子萌发的影响。本文以玉米为材料,比较COR与MeJA 在种子萌发和幼苗生长方面的异同,明确COR和MeJA调控玉米种子萌发的效应,为种子萌发调控提供一条新的可行途径。 1材料与方法 1.1玉米种子和处理药剂 玉米(Zea may L.)品种为农大3138,种子由中国农业大学玉米中心提供。 COR由中国农业大学化控中心发酵生产,菌种为PG4180,浓度已经HPLC 检测标定好,直接用蒸馏水配制成0.001 μmol/L、0.01μmol/L、0.1μmol/L、
植物的抗寒性锻炼与冻害预防 低温下植物的适应性生理生化变化在冬季严寒来临之前,随着日照的缩短和气温的降低,植物体内会发生一系列适应低温的生理生化变化,从而提高了植物的抗寒性. 这种逐步提高抗寒能力的适应过程称为抗寒锻炼(cold hardening)或低温训化(cold acclimation)。 晚秋或早春寒潮突然袭击植物就易受害经适当的抗寒锻炼过程,植物逐渐完成适应低温的一系列代谢变化,获得较强的抗寒性。我国北方晚秋时,植物内部的抗寒锻炼还未完成,抗寒力差;在早春,温度已回升,植物的抗寒力逐渐下降。 植物抗寒锻炼过程中体内发生的适应性生理变化: (1)组织的含水量降低,而束缚水的相对含量增高。 (2)呼吸减弱消耗减少.有利于糖分等的积累,植物的整个代谢强度减弱,抗逆性增强。 (3) ABA(天然脱落酸)含量增多,生长停止,进入休眠 冬小麦的核膜口逐渐关闭,细胞核与细胞质之间物质交流停止,细胞分裂和生长活动受到抑制,植物进入休眠。 植物进入深度休眠后,其抗寒性能力显著增强。 ABA(天然脱落酸)含量保护物质积累可溶性糖含量增加,对细胞的生命物质和生物膜起保护作用。可增加细胞液浓度,降低冰点,提高原生质保水能力,保护蛋白质胶体不致遇冷变性凝聚;可进一步转化为其它保护物质(如磷脂、氨基酸等)和能源. 在抗寒锻炼中,氨基酸的含量也增多. 脯氨酸的含量增加更为明显,是防冻剂或膜的稳定剂,对植物适应多种逆境具有重要作用。 2.低温诱导蛋白(Cold acclimation protein) 植物经低温诱导能使某些特定的基因活化,并得以表达合成一组新蛋白。 近年来,已有近百种植物低温诱导蛋白被发现和研究,但还不清楚它们在提高植物抗寒性过程中的机理。 抗冻蛋白(antifreeze protein AFP) 是生活在两极冰水中的鱼类血液中含有的糖蛋白.能降低细胞间隙体液冰点。植物本身也可能具有与动物中类似的抗冻蛋白和基于相似原理的抗冻能力。拟南芥冷调节蛋白(coldyreguated protein.COR) COR 6.6蛋白油菜的BN28蛋白拟南芥叶绿体的COR15蛋白胚胎发育晚期丰富蛋白(late embryogenesis abunndant protein,LEA) 植物在胚胎发育晚期,种子脱水时大量产生的蛋白质。多数是高度亲水、沸水中稳定的可溶性蛋白. 植物在低温诱导下也能表达多种LEA蛋白。有助于提高植物在冰
茉莉酸甲酯与二氢茉莉酮酸甲酯对悬浮培养的甘草细胞生长和黄酮积累的影响 杨英, 郑辉, 李赟, 季家兴, 余龙江* 华中科技大学生命科学与技术学院, 武汉430074 提要: 建立了稳定的甘草细胞悬浮培养体系, 在一个培养周期内, 细胞的生长曲线呈“S”型, 培养21 d 的干重、鲜重和黄酮产量都达到最高值。甘草细胞悬浮培养体系中分别添加100 μmol·L -1二氢茉莉酮酸甲酯和茉莉酸甲酯时, 虽然对细胞生长有一定程度的抑制, 但细胞中甘草黄酮产量仍有提高。添加二氢茉莉酮酸甲酯和茉莉酸甲酯的最适时间分别为细胞培养后的第5天和第10天。 关键词: 胀果甘草悬浮细胞; 甘草黄酮; 茉莉酸甲酯; 二氢茉莉酮酸甲酯 Effects of Methyl Jasmonate and Methyl Dihydrojasmonate on the Cell Growth and Flavonoids Accumulation in Cell Suspension Culture of Glycyrrhiza inflata Bat. YANG Ying, ZHENG Hui, LI Yun, JI Jia-Xing, YU Long-Jiang * College of Life Science and Technology, Huazhong University of Science and Technology, Wuhan 430074, China Abstract: The effects of methyl jasmonate and methyl dihydrojasmonate on cell growth and flavonoids produc-tion in suspension cultures of Glycyrrhiza inflata Batal were investigated. Biomass accumulation and flavonoids production both exhibited “S” curve in one culture cycle, with the greatest values obtained on day 21. In addition, although the cell growth was restrained to some degree by the methyl dihydrojasmonate and methyl jasmonate treatment with the concentration of 100 μmol·L -1, the flavonoids accumulation was increased. Moreover,the optimum time for the addition of methyl dihydrojasmonate and methyl jasmonate was the 5th day and the 10th day during the cell culture, respectively. Key words: cell suspension cultures of Glycyrrhiza inflata ; flavonoids production; methyl jasmonate; methyl dihydrojasmonate 收稿2008-06-16修定2008-07-22 资助新疆建设兵团新疆昆仑神农股份有限公司提供经费。 * 通讯作者(E -m a i l : Y u l j @h u s t.e d u.c n ; T e l : 027-********)。 采用细胞培养大规模生产甘草的有效成分已有报道, 如梁玉玲等(2000)用胀果甘草(Glycyrrhiza inflata )愈伤组织培养生产甘草酸; 杨世海等(2005)的乌拉尔甘草(G. uralensis )愈伤组织培养条件的探索以及杜旻等(2001)乌拉尔甘草毛状根培养体系的建立等。但有关采用甘草细胞培养生产总黄酮的研究还未见有报道。茉莉酸甲酯(methyl jasmonate,MJ)是一种小的信号分子, 具有广泛的生理作用(刘燕等2007), 它可以作为细胞信使, 参与到植物细胞的代谢过程中, 激活特定的防御相关基因表达, 引起细胞抗逆反应产物表达(宾金华和潘瑞炽1999)。张进杰和徐茂军(2006)研究NO 和茉莉酸甲酯影响黄芩悬浮细胞生长及黄芩苷合成的结果表明, MJ 能增加黄芩苷的合成。余龙江等(1999)的研究表明,茉莉酸甲酯有诱导紫杉醇生物合成的作用。茉莉酸甲酯对植物培养物的次级代谢的诱导也引起人们 的注意(于树宏和李玲2002; 徐亮胜等2005; 罗建平等2006), 但其对胀果甘草悬浮细胞合成甘草黄酮的研究尚未见报道。二氢茉莉酮酸甲酯是MJ 的类似物, 在工业中可以作为一种香料, 但价格比MJ 便宜很多。为此, 本文研究MJ 及其类似物二氢茉莉酮酸甲酯(methyl dihydrojasmonate)对甘草细胞生长和甘草黄酮积累的影响, 以期为甘草细胞规模化生产黄酮类化合物提供参考。 材料与方法 实验材料为我们实验室保存的高产甘草黄酮
植物怎样过冬 冬天天气寒冷,各种植物仍能渡过严寒的冬季,来年继续生长、开花、结果。奥秘在哪里呢? 原来植物在寒冷到来之前,在生理上相应地做出各种适应性反应:如可溶性糖渡度的提高,就可以提高细胞溶液浓渡,使水点降低。还可以缓冲原生质过度脱水,保护原生质胶体不致遇冷凝固。另外糖分子还有巨大的表面活动能力,可以吸附在细胞器的表面之上,减弱它们的生命能力。细胞内糖多,渗透压加大,保留水分多,减少外出结冰。还有的植物通过降低自身含水量,以适应低温条件,安全渡过寒冷的冬季。 当初冬温度降到5度左右,冬小麦的地上生长基本停止,但光合作用仍继续缓慢进行,这时所合成的产物并不转化成淀粉或其他非溶性物质,而是以可溶性糖类(主要是葡萄糖)积存于细胞中。由于冬季麦苗叶绿素形成少,细胞呈中性或微酸性,此时,麦苗颜色开始变红,这才是麦苗抗寒能力强,生长正常的一种标志。 果树花芽也能安全越冬,才能使来年花开满树,结出丰收的果实。这主要靠得是花芽内部含水量的变化。当气温下降时,花芽迅速排出内部的水,使芽内的汁液达到高度渡缩的程度。这种高渡度汁液具有极强的抗冻能力,它在严寒时也不会结冰因此,防止了细胞膜由于冰冻而引起破裂,即使气温下降到零下30度时,花芽内细胞仍能安然无恙。 植物的抗寒性 1. 低温下植物的适应性生理生化变化 在冬季严寒来临之前,随着日照的缩短和气温的降低,植物体内会发生一系列适应低温的生理生化变化,从而提 高了植物的抗寒性. 这种逐步提高抗寒能力的适应过程称为抗寒锻炼(cold hardening)或低温 训化(cold acclimation)。 ·晚秋或早春寒潮突然袭击植物就易受害 经适当的抗寒锻炼过程,植物逐渐完成适应低温的一系列代谢变化,获得较强的抗寒性。 我国北方晚秋时,植物内部的抗寒锻炼还未完成,抗寒力差;在早春,温度已回升,植物的抗寒力逐渐下降。 植物抗寒锻炼过程中体内发生的适应性生理变化 (1)组织的含水量降低,而束缚水的相对含量增高。 (2)呼吸减弱消耗减少.有利于糖分等的积累,植物的整个代谢强度减弱,抗逆性增强。 (3) ABA含量增多,生长停止,进入休眠 ·冬小麦的核膜口逐渐关闭,细胞核与细胞质之间物质交流停止,细胞分裂和生长活动受到抑制,植物进入休眠。
植物抗寒性鉴定 我国植物种类繁多,分布区域广,在晚秋和早春时期发生的冻害和冷害两种低温危害,常常给越冬作物和果木造成严重伤害。冻害由0℃以下低温造成,冷害由0℃以上低温引起,冷害对植物的伤害程度,除取决于低温外,还取决于低温维持时间的长短。植物抗寒性的强弱决定其生长季节,因此蔬菜作物利用抗寒品种,可以将露地栽培提前,提早供应市场;而选育抗寒性强的果树品种,不仅是寒带果树育种者的主攻方向,而且也是温带甚至热带果树育种者重要育种目标之一。 本实验重点学习实验室间接鉴定果树抗寒性的方法和步骤。 一、试材及用具 1.试材及处理:植物枝条或花朵,将采回的枝条剪成40cm左右的长度,用自来水冲洗数遍(洗掉泥土、灰尘、虫卵),再用蒸馏水冲洗三次,然后用吸水纸吸干水分,最后将枝条末端进行蜡封。将每个品种蜡封后的枝条分成相等的6份,其中一份作为对照,其余每份作为一个低温处理,放于冰箱中(0℃~4℃)保存备用。每次处理时,各取参试品种的一份枝条放于超低温冰箱或程控冰箱内进行低温处理,处理温度梯度为:CK(0℃),-20℃,-25℃,-30℃,-35℃,-40℃。降温速度为4℃/h,达到目的处理温度后维持12h,然后逐步升温,升温速度亦为4℃/h。花朵的处理温度梯度为:CK(0℃),-1℃,-3℃,-5℃,-6℃,-7℃,-8℃。 2.仪器烘箱,发芽箱,培养皿,标牌,电导仪,具塞刻度试管(20ml),恒温水浴锅,温度计,玻璃棒,天平,研钵,石英砂,高速台式离心机,分光光度计,微量进样器,荧光灯(4000lx),容量瓶(250ml、25ml),聚丙烯酰胺凝胶垂直板电泳槽,刻度吸管(10ml、5ml),离心管等。 二、内容说明 植物的抗寒性鉴定可分为田间鉴定和实验室间接鉴定两种方法。 1.田间鉴定田间自然鉴定就是在冻害发生期(早春及晚秋)对受冻的田间植株一定器官、组织以一定的标准进行评价、比较,然后根据冻害情况评价抗寒性。陈学森等(2001)对山东省春季“倒春寒”发生后,受害的核果类果树的花器官的抗寒性进行了调查,选出了当地花期抗寒性较强的红荷包、红丰等杏品种,证明
论植物的抗寒性和抗热性 姓名毛丹宁班级市场营销1101 学号20116512摘要:地球上的植物种类繁多,形态结构和生活习性各不相同,一定地区植物群落构成热带亚热带温带寒带四种植被类型【1】,植物的生长受各方面因素的影响,如温度气候等因素,在一定的温度范围内,温度与植物生长成正相关【2】。而周围环境对植物生长也有很大的影响,对不同环境的适应体现了植物的抗逆性,如抗寒性,抗热性等。 关键词:抗热性,抗寒性,机理,措施 引言:植物的抗逆性是指植物对逆境的适应性反应,任何植物的抗逆性都不是突然形成的,而是逐步适应形成的【3】,植物的抗寒性与抗热性就是经过一段时间的锻炼而形成的。 1 植物的抗寒性 1·1寒害与植物抗寒性 由低温引起的植物伤害现象统称为寒害,包括冻害和冷害,植物对低温的适应性和抵抗能力统称为抗寒性【4】,植物寒害机制和抗寒性是同一个问题的两个方面,植物在冬季来临之前,随着温度的逐渐降低,体内发生一系列的变化,增强抗寒力,在气温回暖的过程中抗寒力又逐渐消失【3】。 1·2植物对抗寒的生理适应 植物在长期的进化过程中,对冬季低温在生长习性方面有各种特殊的适应方式,在冬季来临之前,随着气温的逐渐降低,体内发生了一系列的适应低温的生理变化,抗寒力逐渐加强,这个提高抗寒能力的过程称为抗寒锻炼,即使抗寒性很强的植物,在未进行过抗寒锻炼之前,对寒冷的抵抗能力还是很弱的【5】。 植物在生理生长方面对低温的适应性变化有:随着温度的下降,植物吸水较少,含水量逐渐下降,抗寒锻炼过程中细胞内吸水性胶体加强,束缚水含量提高,自由水含量减少,总含水量减少,使植物抗寒性得以加强;随着温度下降,呼吸逐渐减弱,消耗的糖分减少,增加糖的积累,代谢活动有利于对不良环境的抵抗;随着温度的降低,脱落酸含量增多;随着温度的降低,植物生长逐渐停止,并逐渐进入休眠期;当温度逐渐下降时,淀粉水解成糖比较旺盛,保护物质增多【5】。 1·3内外条件对抗寒性的影响 内部因素:各种植物原产地不同,生长期长短不同,对温度要求也不一样,同一作物不同品种之间抗寒性差别也很明显,而同一植物不同生长期的抗寒性也不相同,一般来说,各种较强的品种抗寒性较强,各种较弱的品种抗寒性弱。 外部因素:抗寒性强弱与植物所处休眠状态及抗寒锻炼情况有关,所以影响休眠和抗寒锻炼的
植物抗寒性及其相关基因工程研究进展 摘要:低温是影响植物分布、产量及品质的重要非生物胁迫因素,提高植物的抗寒性是作物育种的重要任务。近年来,植物抗寒分子机理研究不断深入,植物抗寒基因工程研究获得了长足的进展。本文从膜稳定性、抗氧化酶活性、抗冻蛋白、低温信号转录因子和渗透调节物质等方面对植物耐冷性基因工程研究进展进行了分析、归纳与总结,旨在为植物抗寒机理研究及植物抗寒育种提供参考。 关键词:植物;抗寒性;基因工程;抗寒育种。 Research Advance in the Cold-resistance and the related Genetic Engineering in Plants Abstract: Low-temperature is a abiological intimidation affecting plant's distribution、output and quality, and the breedingof cold-resistant plants is one of the most important tasks in the development of agriculture's.In recent years,plant's cold-resistant genetic engineering has acquired great progress, with the mechanism of cold- resistance is studied deeper and deeper.This paper gives a generalstatement about the recent development of plant genetic engineering for cold-resistance in composition of membrane lipids,the activity of antioxidang enzymes,the antif reeze protein,the transcription factors of lowtemperature cell signaling and osmolytes,aiming to provide some useful information and ideas to research-ers who work on plant breeding and the mechanism of cold-resistance. Key words:plants; cold-resistance; genetic engineering; plant breeding of cold-resistance. 温度是气候因素中影响植物地理分布的主导因素,低温胁迫是植物栽培中经常遇到的一种灾害,涉及到粮食作物、园艺植物及其它许多经济植物。低温影响植物的生长代谢,引起植物相关生理指标变化,导致植物的生长受损,严重时导致死亡。 低温不仅限制植物的种植范围,也会造成减产和品质下降,严重时甚至绝收。因此,有关植物抗寒性的研究一直是植物学研究领域的热点之一。早期研究主要集中在植物遭寒害后的细胞形态结构、生理、生物化学变化等方面。对植物的抗寒分子机理认识不够,提高植物抗寒性主要依靠传统育种方法,虽然也取得一些进展,但由于其周期长且工作繁琐,已不适应现代农业的发展要求[1]。近年来,随着对植物抗寒分子机理研究的不断深入和分子生物学技术的迅猛发展,抗寒研究已经从生理水平深入到分子水平,促进了植物抗寒基因工程的发展。 1植物抗寒生理研究进展 1.1 生物膜系统与抗寒性的关系 1973年,Lyons提出了著名的“膜脂相变”假说[2],认为温度下降时,生物膜首先发生膜脂的物相变化,从液晶相变为凝胶相,膜的外型和厚度也发生变化,