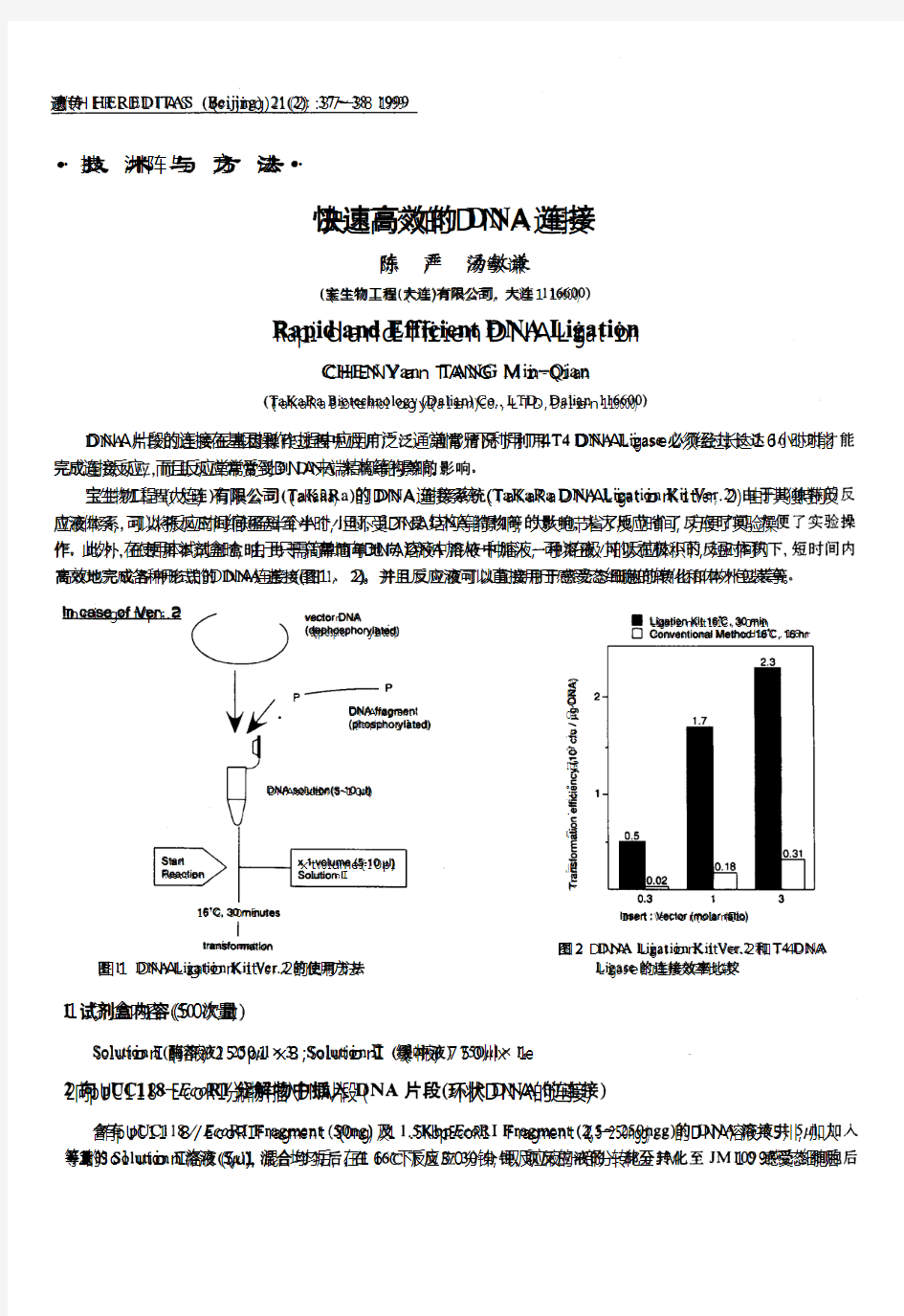
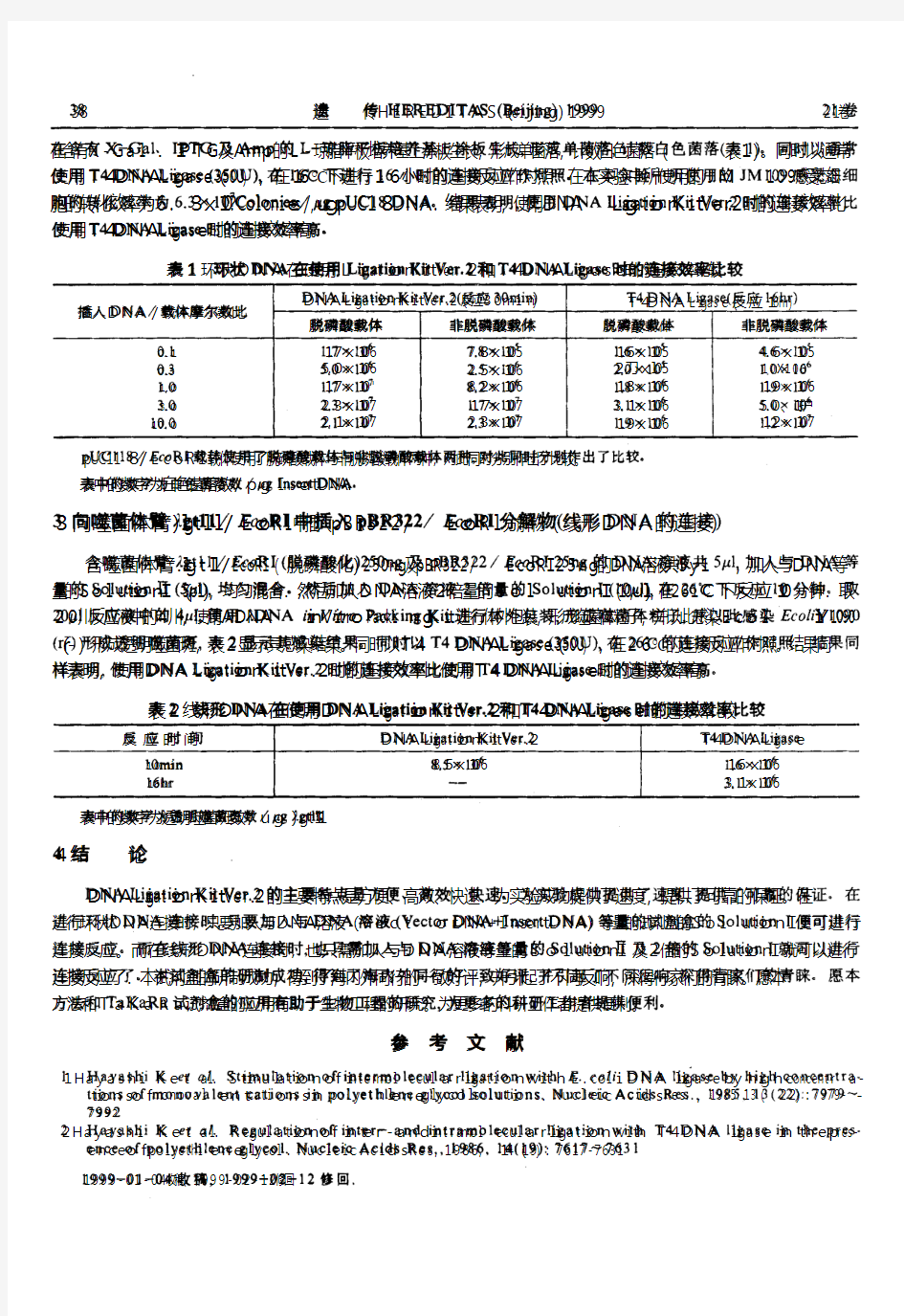
一. 重组质粒的构建 T质粒载体 重组的DNA分子是在DNA连接酶的作用下,有Mg2 、ATP存在的连接缓冲系统中,将分别经酶切的载体分子与外源DNA分子进行连接。 DNA连接酶有两种:T4噬菌体DNA连接酶和大肠杆菌DNA连接酶。两种DNA连接酶都有将两个带有相同粘性末端的DNA分子连在一起的功能,而且T4噬菌体DNA连接酶还有一种大肠杆菌DNA连接酶没有的特性,即能使两个平末端的双链DNA分子连接起来。但这种连接的效率比粘性末端的连接率低,一般可通过提高T4噬菌体DNA 连接酶浓度或增加DNA浓度来提高平末端的连接效率。 T4噬菌体DNA 连接酶催化DNA 连接反应分为3 步:首先,T4 DNA 连接酶与辅因子ATP形成酶-ATP复合物;然后,酶-ATP复合物再结合到具有5’磷酸基和3’羟基切口的DNA上,使DNA腺苷化;最后产生一个新的磷酸二酯键,把切口封起来。连接反应通常将两个不同大小的片断相连。 很多DNA聚合酶在进行PCR扩增时会在PCR产物双链DNA每条链的3’端加上一个突出的碱基A。pUCm-T载体是一种已经线性化的载体,载体每条链的3’端带有一个突出的T。这样,pUCm-T载体的两端就可以和PCR产物的两端进行正确的AT配对,在连接酶的催化下,就可以把PCR产物连接到pUCm-T载体中,形成含有目的片断的重组载体。 连接反应的温度在37℃时有利于连接酶的活性。但是在这个温度下粘末端的氢键结合是不稳定的。因此采取折中的温度,即12-16℃,连接12-16h(过夜),这样既可最大限度地发挥连接酶的活性,又兼顾到短暂配对结构的稳定。 二. 感受态制备原理 细菌在0°C CaCl 低渗溶液中胀成球形,丢失部分膜蛋白,成为容易 2 吸收外源DNA的感受态。 三. β-半乳糖甘酶显色反应选择法 LacZ基因是大肠杆菌乳糖操纵子中的一个基因,可以编码β—半乳糖核苷酶。β—半乳糖核苷酶是由4个亚基组成的四聚体,可催化乳糖的水解.用X-Gal为底物进行染色时,呈蓝色。 现在一些特定的质粒(比如pUC/pBS等),常带有β—半乳糖核苷酶的调控序列和β—半乳糖核苷酶N端146个氨基酸(α肽段)的编码序列,在这个编码序列里还插入一个多克隆位点(MCS),它并不影响lacZ的表达。另外,常用的大肠杆菌带有β—半乳糖核苷酶C端部分序列(β肽段),的编码序列。在各自独立的情况下,这些质粒与大肠杆菌各自编码的β—半乳糖核苷酶片段都没有酶的活性。只有当携带α
限制性核酸内切酶(以下简称限制酶):限制酶主要存在于微生物(细菌、霉菌等)中。一种限制酶只能识别一种特定的核苷酸序列,并且能在特定的切点上切割DNA分子。是特异性地切断DNA链中磷酸二酯键的核酸酶(“分子手术刀”)。发现于原核生物体内,现已分离出100多种,几乎所有的原核生物都含有这种酶。是重组DNA技术和基因诊断中重要的一类工具酶。例如,从大肠杆菌中发现的一种限制酶只能识别GAATTC序列,并在G和A 之间将这段序列切开。目前已经发现了200多种限制酶,它们的切点各不相同。苏云金芽孢杆菌中的抗虫基因,就能被某种限制酶切割下来。在基因工程中起作用。 DNA连接酶:主要是连接DNA片段之间的磷酸二酯键,起连接作用,在基因工程中起作用。 DNA聚合酶:催化脱氧核苷酸之间的聚合反应。主要是连接DNA片段与单个脱氧核苷酸之间的磷酸二酯键,在DNA复制中起做用。 DNA聚合酶只能将单个核苷酸加到已有的核酸片段的3′末端的羟基上,形成磷酸二酯键;而DNA连接酶是在两个DNA片段之间形成磷酸二酯键,不是在单个核苷酸与DNA片段之间形成磷酸二酯键。 DNA聚合酶是以一条DNA链为模板,将单个核苷酸通过磷酸二酯键形成一条与模板链互补的DNA链;而DNA连接酶是将DNA双链上的两个缺口同时连接起来。因此DNA连接酶不需要模板。 RNA聚合酶(又称RNA复制酶、RNA合成酶)的催化活性:RNA聚合酶以完整的双链DNA为模板,转录时DNA的双链结构部分解开,转录后DNA仍然保持双链的结构。真核生物RNA聚合酶:真核生物的转录机制要复杂得多,有三种细胞核内的RNA聚合酶:RNA 聚合酶I转录rRNA,RNA聚合酶II转录mRNA,RNA聚合酶III转录tRNA和其它小分子RNA。在RNA复制和转录中起作用。 反转录酶:属RNA指导的DNA聚合酶,具有三种酶活性,即RNA指导的DNA聚合酶,RNA酶,DNA指导的DNA聚合酶。在分子生物学技术中,作为重要的工具酶被广泛用于建立基因文库、获得目的基因等工作。在基因工程中起作用。
1. T4DNA连接酶: 本酶催化相邻DNA链的5’-P末端和3’-OH末端以磷酸二酯键结合的反应,(将已有的两个DNA片段连接成为一条DNA链的酶,常用于基因工程,将目的基因连在质粒载体上,作用于两脱氧核糖核苷酸间的磷酸二脂键),需ATP作辅酶。本酶不仅可以催化粘性末端之间或平滑末端之间的DNA的连接,也可以催化DNA与RNA之间以及少数RNA之间的连接。 T4DNA连接酶可连接DNA-DNA,DNA-RNA,RNA-RNA和双链DNA粘性末端或平头末端。无论是T4DNA连接酶,还是大肠杆菌DNA 连接酶都不能催化两条游离的DNA链相连接。T4 DNA连接酶常用于催化双链DNA平末端或互补粘性末端之间的连接反应,也能催化双链RNA 5'-磷酸末端和3'-羟基末端间的连接。还可以修复双链DNA、RNA 或DNA/RNA杂交双链中的单链切口。以上反应均需消耗ATP。 粘末端的连接反应: 插入片段和载体的摩尔浓度比特别重要,此比例在2-6之间最好,低于2:1就会导致较低的连接效率,高于6:1则会导致多个插入。摩尔比请按载体与插入片段 DNA浓度及分子大小来计算。 平滑末端的连接反应:平滑末端的连接反应与突出末端相比反应较慢(其Km值约为突出末端的100倍)。进行平滑末端的连接反应时,可提高DNA浓度,将使用酶量增加到突出末端量的2~5倍左右。 与粘粒或噬菌体进行连接反应:可使载体和插入DNA的摩尔比调整为1:1,同时增大DNA浓度以便取得良好效果。 (0.05-0.1 μg/ul以上)。 反应温度:该酶的最适温度为37℃,由于热稳定性较差,因此长时间反应时通常需在16℃下进行。若反应1-2小时左右的话也可在室温下进行反应。 抑制剂:T4 DNA连接酶要求Mg 2+,因此螯合Mg 2+的EDTA 的存在会阻碍反应。将溶解于含有高浓度EDTA缓冲液中的 DNA准备作为样品使用时,最好先用灭菌蒸馏水或TE缓冲液进行置换。 2. T4 DNA聚合酶: T4DNAPolymerase,即T4DNA聚合酶,是一种模板依赖的DNA聚合酶,可以依赖于DNA模板对5' 端突出末端进行补平;同时可对3' 端突出末端进行削平。也可以在结合有引物的单链DNA模板上,从5'→3'方向催化DNA合成反应。 特点:T4DNAPolymerase由于同时具有5'→3' DNA聚合酶活性和
实验一载体与目的基因的连接与转化以及 重组DNA的提取与酶切鉴定 一、实验目的 1.CaCl2法制备感受态细胞 2.目的基因与载体连接(c-myc+pSV2;粘端连接) 3.重组质粒转化大肠杆菌并筛选转化体(HB101;Amp r) 4.质粒DNA的小量快速制备 5.质粒DNA的限制性内切酶酶切 6.DNA的琼脂糖凝胶电泳 二、实验原理 通过粘端连接法将具有相同粘性末端的DNA分子连接在一起,通过碱基配对氢键形成一个相对稳定的结构,利用连接酶发挥间断修复的功能,从而获得重组的DNA分子。 受体细胞经处理后(电击或CaCl2等处理),细胞膜通透性发生变化,从而使外源的载体分子通过感受态细胞,并使受体细胞获得新的稳定遗传的性状,该过程称为转化。由于本实验种pSV带有抗氨苄青霉素的基因,因而转化后的细胞在含氨苄青霉素的平板上培养可以筛选出转化成功的受体细胞。 分离质粒DNA的步骤包括:培养细菌使质粒扩增、收集和裂解细菌以及分离和纯化质粒DNA。SDS可以使细胞壁裂解,碱变性抽提质粒DNA的原理是利用染色体DNA与质粒DNA的变性复性的差异达到分离目的,当pH>12.6时,染色体DNA氢键断裂,双螺旋结构解开而变性,质粒DNA由于超螺旋共价闭合环状结构,两条互补链不会完全分离。当采用pH 4.8的NaAc高盐缓冲液调节pH至中性时,质粒DNA恢复原有的构型,而染色体DNA则不能复性而缠绕形成网状结构。通过离心可将染色体DNA及大分子RNA、蛋白质等去除。 三、实验器材和试剂 1.器材 恒温摇床、电热恒温培养箱、电热恒温水浴、台式离心机、低温离心机、涡旋振荡器、移液枪及枪头、1.5 ml离心管、制冰机、三角推棒、酒精灯、细菌培
DNA聚合酶和DNA连接酶的比较 “红细胞”专题复习 山东省青岛市第九中学辛建福 红细胞不仅在动物体内起着非常重要的生理作用,还作为生物科学某些领域研究的好材料, 且课本涉及的地方有多处。因此,有关红细胞知识点常成为高考命题的切入点。现将有关的考点归纳总结如下:
考点1.组成细胞的分子 例1.“朴雪”乳酸亚铁口服液可以有效地治疗人类缺铁性贫血症,这是因为其中的Fe2+进入人体后能() A.调节血液的酸碱平衡 B.调节血液的渗透压 C.构成中的血红蛋白 D.促使更多红细胞的产生 解析:此题考查红细胞内血红蛋白特有的无机盐组成和无机盐的生理作用。某些无机盐可以用来构造细胞内某些复杂化合物的重要组成部分,如Fe2+进入人体后构成血红蛋白的主要成分,Mg2+是叶绿素分子必需的成分。故答案选C。 答案: C 考点2.细胞的基本结构 例2.若从成熟的红细胞上获取细胞膜,可用来处理细胞的试剂是() A.10%盐酸 B.蛋白酶 C.磷脂酶 D.清水 解析:此题考查成熟红细胞膜的化学物质组成和分离出纯细胞膜的方法。获取细胞膜就是让其破裂,让内部物质释放出。10%盐酸浓度高,使红细胞皱缩甚至杀死;脂类和蛋白质是细胞膜的主要组成物质,故蛋白酶、磷脂酶均使膜结构遭破坏;清水使细胞渗透吸水胀破,内部物质流出只剩细胞膜。故选D。
答案: D 例3.人的红细胞和精子的寿命都很短,这一事实体现了() A.环境因素的影响 B.功能对寿命的决定 C.核质相互依存的关系 D.遗传因素的作用 解析:此题考查红细胞的寿命和细胞的完整性。细胞的各个部分不是彼此孤立的,而是互相紧密联系、协调一致的,实际上一个细胞就是一个有机的统一整体,细胞只有保持完整性,才能够正常地完成各项生命活动。例如人体成熟的红细胞、人工去核的细胞和丢弃大部分细胞质的精子细胞,一般不能存活多久,有力地说明细胞完整性的重要意义。故答案选C。 答案:C 考点3.细胞的生命历程 例4.青蛙红细胞的分裂方式是() A.二分裂 B.无丝分裂 C.有丝分裂 D.减数分裂 解析:此题考查非哺乳类动物红细胞的结构和蛙的红细胞独特的分裂方式。无丝分裂的特点是在分裂开过程中核膜、核仁并不消失,也无染色体变化和纺锤体丝出现,它是真核细胞的一种分裂方式,如蛙的红细胞分裂方式就是这样。二分裂是指单细胞生物(如细菌)一种常见的繁殖方式,进行分裂生殖时,先是核逐渐延长,然后逐渐分成两个新个体。虽然两者都要“一分为二”,但分裂的机理和本质有所不同。人和哺乳动物
T4 DNA Ligase即T4 DNA连接酶,可以催化粘端或平端双链DNA或RNA的5’-P末端和3’-OH末端之间以磷酸二酯键结合,该催化反应需A TP作为辅助因子。同时T4 DNA连接酶可以修补双链DNA、双链RNA 或DNA/RNA杂合物上的单链缺刻(single-strand nicks)。 用途:T4 DNA Ligase常用于DNA片段和载体、linker或adaptor等的连接。也可以用于缺刻修复及Ligase 介导的RNA检测。 来源:本T4 DNA Ligase由大肠杆菌表达,表达基因的来源为T4嗜菌体。 活性定义:One unit is defined as the amount of enzyme required to give 50% ligation of HindIII fragments of lambda DNA in 30 min at 16℃in 20 μl of the assay mixture containing 50 mM Tris, pH 7.5, 10 mM MgCl2, 10 mM DTT, 1 mM ATP, 25 μg/ml BSA and a 5'-DNA termini concentration of 0.12 μM(300 μg/ml)。200U等于1个Weiss unit,以Weiss unit计,本产品共1000单位。 纯度:不含DNA内切酶、外切酶和磷酸酯酶,不含RNA酶,满足常规连接反应要求。 酶储存溶液:20 mM Tris, pH 7.5, 50 mM KCl, 1 mM DTT, 0.1 mM EDTA and 50%(v/v)glycerol。 10X Ligation Buffer:400 mM Tris, pH 7.8, 100 mM MgCl2, 100 mM DTT, 5 mM A TP。 失活或抑制:65℃孵育10分钟可以导致T4 DNA Ligase失活;NaCl或KCl浓度大于200mM时强烈抑制T4 DNA Ligase。 包装清单: 产品编号产品名称包装 D7008-1 T4 DNA Ligase(1000U/μl) 200,000U D7008-2 10X Ligation Buffer 1.5ml -说明书1份 保存条件: -20℃保存。 注意事项: 对于普通的转化大肠杆菌的操作,不必对连接产物进行纯化,连接产物可以直接用于转化。但用电转方法转化大肠杆菌时,通常宜先用DNA纯化试剂盒或酚氯仿抽提方法等纯化DNA,然后再进行电转。 需进行平端连接或快速连接时,推荐使用碧云天的快速DNA连接试剂盒(D7002/D7003)。T4 DNA Ligase 可以进行平端连接,但效率较低。 普通的连接反应不必进行凝胶电泳观察。如果需要对于连接产物进行凝胶电泳观察,推荐先在65℃孵育10分钟使T4 DNA Ligase失活,以避免T4 DNA Ligase和DNA结合导致的条带位置迁移(band shift)。 为了您的安全和健康,请穿实验服并戴一次性手套操作。
试验一、DNA连接和转化 质粒的转化是指将质粒或以它为载体构建的重组子导入细菌的过程。将连接产物转化到感受态细胞中,实现重组克隆的增殖,便于后续分子操作。可以采用多种方法筛选和鉴定目的克隆。 一、实验目的 掌握DNA的连接方法和热激法转化大肠杆菌感受态细胞及转化子的鉴定原理和实验方法。 二、实验原理 1)限制性内切酶可识别特定位点并切割DNA 产生粘性末端或平端的外源片段,经DNA 的纯化处理后用于连接反应;选择克隆载体多克隆位点上相应的限制性内切酶切割,并用碱性磷酸酶处理防止载体自连;在连接酶的作用下将外源片段连接到载体上,实现外源片段的克隆。 2)TA克隆:克隆载体用限制性内切酶酶切后,再在两侧的3端添加“T”。大部分耐热性DNA聚合酶进行PCR反应时都有在PCR产物的3末端添加“A”的特性。二者在DNA连接酶的作用下利用粘性末端进行连接。 3)热激法:大肠杆菌在0 ℃CaCl2低渗溶液中,细菌细胞膨胀成球形,转化混合物中的DNA 形成抗DNase 的羟基-钙磷酸复合物粘附于细胞表面,
经42℃短时间热冲击处理,促进细胞吸收DNA 复合物,在丰富培养基上生长数小时后,球状细胞复原并分裂增殖。在被转化的细胞中,重组子基因得到表达,在选择性培养基平板上可挑选所需的转化子。 三、仪器和材料 超净工作台、恒温培养箱、移液枪、冷冻循环水浴锅、恒温摇床。 pMD18-T Vector、DNA片段、Solution I、感受态细胞DH5α、Amp+ LB液体培养基、Amp-LB液体培养基、Amp-LB固体培养基、X-gal (20mg/ml)、IPTG(200mg/ml)。 四、实验步骤 1、连接反应步骤: 1)在离心管中配制下列混合溶液,全量为5μl。(冰上操作) pMD18-T Vector 1μl DNA 1μl H2O 3μl 2)加入5μl的solution I。(冰上操作) 3)16℃反应30min。 2、转化反应步骤: 1) 将连接产物10μl加入到已制备好的感受态细胞中,冰上孵育30min。 2) 42℃水浴中热休克90s,立即冰上冷却2min。 3)加入200μl不含氨苄(amp)的LB液体培养基,37℃,150rpm,振荡培养40min。 4) 加40μl的X-Gal (20mg/ml)和8μl IPTG(200mg/ml)。 5) 约200μl转化产物涂布于培养平板上。放置于37℃培养箱,待溶液被琼 脂吸干,倒置平板,37℃,培养16h。 6)第二天上午观察蓝白斑筛选结果。 五、作业 1、实验结果。(平板图片) 2、在热激以后进行活化培养,这时的培养基中为什么不加入氨苄青霉素?
目的基因与载体连接、感受态制备及转化 【实验原理】 1;酶促生物化学反应过程 在一定的条件下,由DNA连接酶催化目的基因与载体相邻的5’端磷酸与3’端羟基之间形成磷酸二酯键的过程。相同或不同的限制性内切酶产生相同的粘性末端,在降至退火温度时,能重新互补结合,在DNA连接酶的催化下,目的基因与载体相连接。 2;DNA连接酶的分类: T4 DNA连接酶:催化dsDNA粘末端连接及平端连接 大肠杆菌DNA连接酶:不能催化平末端连接,其底物只能是带缺口的双链DNA分子和具同源互补粘末端的不同DNA分子 3;T4 DNA连接酶:来源T4噬菌体感染的大肠杆菌 最佳pH值7.2~7.8,常用的反应液为pH7.6 的Tris-HCl缓冲液 需ATP,Mg2+参加反应 二硫苏糖醇等巯基化合物可促进连接酶的连接 作用;高浓度的Na+、K+等抑制酶的活性。 4;受体分类:受体细胞也称为宿主,是重组子扩增及表达的场所,分为原核细胞和真核细胞两类。 5;应用:原核细胞:重组子复制扩增,外源基因表达系统 真核细胞:主要用于外源基因的表达 6;转化:特指以质粒DNA活以它作为载体构建的重组子导入细菌的过程。 转染:指噬菌体、病毒或以它们作为载体构建的重组子导入细胞的过程。 7;感受态细胞:受体细胞经过一些特殊方法处理后,细胞膜的通透性发生变化,成为最适摄取和容纳外源DNA的生理状态。 常用方法:0.1mol/L CaCl2 特点:a.重组酶缺陷,限制修饰系统缺陷 b.不存在载体的筛选标记 c.接受DNA的位点暴露 d.细胞膜通透性增加 8;不同层次,不同水平上进行筛选,以区别转化子与非转化子、重组子与非重组子,以及鉴定所需的特异性重组子。 直接筛选:针对载体携带的标记和插入DNA片段 1.抗性筛选(抗生素平板,ampR , tetR , neoR) 2.标志补救(α-互补,蓝白斑筛选) 3.PCR 4.限制性内切酶消化 4.DNA测序 间接筛选:针对插入片段的蛋白产物,免疫学筛选 9;蓝白斑筛选是根据载体的遗传特征筛选重组子,如α-互补、抗生素基因等。现在使用的许多载体都带有一个大肠杆菌DNA的短区段,其中有β-半乳糖苷酶基因(lacZ)的调控序列和前146个氨基酸的编码信息。在这个编码区中插入了一个多克隆位点(MCS),它并不破坏读框,但可使少数几个氨基酸插入到β-半乳糖苷酶的氨基端而不影响功能,这种载体适用于可编码β-半乳糖苷酶C端部分序列的宿主细胞。因此,宿主和质粒编码的片段虽都
植物基因工程中目的基因的转化方法 张亚欢 20102170231 目的基因的直接转化法和间接转化法 摘要: 植物基因工程中目的基因的转化方法有间接转化法和直接转化法,主要阐述各种转化方法的原理、特点,以期为植物转基因方法的选择提供帮助. 关键词:植物基因工程; 转化方法; 间接转化法; 直接转化法 植物基因工程是通过导入外源基因对植物进行遗传转化,使植物获得抗虫、抗病、抗逆境、优质高产以及生产药物等诸多能力,从而使人们最大限度地利用植物资源。在植物基因工程中最为关键的是植物基因工程技术,该技术是以纯化的外源DNA导入植物细胞以期获得转基因植物的方法,其内容包括:目的基因的分离和鉴定;植物表达载体的构建;植物细胞的遗传转化;转化值物细胞的筛选;转基因植物的鉴定;外源基因表达的检测等[1]。自20 世纪90 年代以来,人们对外源基因导入植物细胞进行了大量的研究,先后采用了多种方法对目的基因进行遗传转化,迄今为止,已经建立了10 余种基因转化方法,这些方法可以分为两大类: 即间接转化法和直接转化法. 下面就这两类转化方法从其原理、特点作一简单介绍. 1 间接转化法 间接转化法是以生物体为媒介的植物转基因方法,有农杆菌介导法和病毒介导法. 1.1 农杆菌介导法 农杆菌介导法是以农杆菌为媒介对植物进行遗传转化的方法,该方法广泛地应用于愈伤组织、悬浮细胞、叶圆盘、茎切段、子叶切片、下胚轴切段、大田植株花茎的切段和薄层细胞等离体材料的转化,是目前双子叶植物常用的基因转移方法. 它是通过根癌农杆菌(Agrobacterim tumefaciens)的Ti 质粒(Tumer inducing plasmid) 和发根农杆菌(Agrobacterim rhizogenis )的Ri质粒(Root inducing Plasmid) 上的一段T一DNA区在农杆菌侵染植物形成肿瘤的过程中,T一DNA可以被转移到植物细胞并插入到染色体基因中[2]。由于T一DNA能够进行高频率的转移,而且Ti质粒和Ri质粒上可以插入到50kb 的外源基因,因此Ti 质粒和Ri质粒就成为植物基因转化的理想载体系统. 1.1.1 Ti 质粒载体系统 Ti 质粒是Lenent 等从根癌农杆菌中分离到的一种巨大质粒,可将外源基因置换T一DNA中的非必需序列使得外源基因整合到受体染色体上而获得稳定的表达,并能使植物细胞转化为肿瘤状态,大量合成冠瘿碱。冠瘿碱是根癌农杆菌的唯一碳源,有利于根癌农杆菌的繁殖和Ti质粒的转移,并进一步扩大侵染领域。 随着根癌农杆菌介导法基因转化技术的逐渐成熟与完善,它已成为常规育种的重要辅助手段,但在植物基因工程的实际操作中使用野生型的Ti 质粒直接作为基因克隆的载体仍然还有一些困难:①Ti质粒分子量很大,很难用常规的方法操作,另外在T一DNA 区段上不可能找到单一的DNA限制内切酶位点,不能插入外源DNA片段; ②野生型Ti 质粒对感染的植物具有毒性并能诱发冠瘿病,导入植物组织使其不能再生出健康的正常植株[3],因此野生型的Ti 质粒必须经过
DNA连接反应 (一)外源DNA和质粒载体的连接反应 外源DNA片段和线状质粒载体的连接,也就是在双链DNA5'磷酸和相邻的3'羟基之间形成的新的共价链。如质粒载体的两条链都带5'磷酸,可生成4个新的磷酸二酯链。但如果质粒DNA已去磷酸化,则吸能形成2个新的磷酸二酯链。在这种情况下产生的两个杂交体分子带有2个单链切口,当杂本导入感受态细胞后可被修复。相邻的5'磷酸和3'羟基间磷酸二酯键的形成可在体外由两种不同的DNA连接酶催化,这两种酶就是大肠杆菌DNA连接酶和T4噬菌体DNA连接酶。实际上在有克隆用途中,T4噬菌体DNA连接酶都是首选的用酶。这是因为在下沉反应条件下,它就能有效地将平端DNA片段连接起来。 DNA一端与另一端的连接可认为是双分子反应,在标准条件下,其反应速度完全由互相匹配的DNA末端的浓度决定。不论末端位于同一DNA分子(分子内连接)还是位于不同分子(分子间连接),都是如此。现考虑一种简单的情况,即连接混合物中只含有一种DNA,也就是用可产生粘端的单个限制酶切割制备的磷酸化载体DNA。在瓜作用的底物。如果反应中DNA浓度低,则配对的两个末端同一DNA分子的机会较大(因为DNA分子的一个末端找到同一分子的另一末端的概率要高于找到不同DNA分子的末端的概率)。这倦,在DNA浓度低时,质粒DNA重新环化将卓有成效。如果连接反应中DNA浓度有所增高,则在分子内连接反应发生以前,某一个DNA分子的末端碰到另一DNA分子末端的可能性也有所增大。因此在DNA浓度高时,连接反的初产物将是质粒二聚体和更大一些的寡聚体。Dugaiczyk等(1975;同时参见Bethesda Res,Lab.出版的Focus第2卷,第2、3期合刊)从理论上探讨了DNA浓度对连接产物性质的影响。简而言之,环化的连接产物与多联体连接产物的比取决于两个参数:j和i。j是DNA分子的一个末端在同一分子的另一末端附近的有效浓度,j的数值是根据如下一种假设作出的:沉吟液中的DNA呈随机卷曲。这样,j与DNA分子的长度成反比(因为DNA越长,某一给定分子的两末端的越不可能相互作用),因此j对给定长度的DNA分子来说是一个常数,与DNA深度无关。j=[3/(3πlb0)]3/2其中l是DNA长度,以cm计,b是随机卷曲的DNA区段的长度。b的值
快速DNA连接试剂盒 概述: 克隆中两个重要步骤包括外源DNA与载体的连接和重组DNA的转化。连接反应是通过连接酶完成的,连接反应需要ATP和镁离子催化双链DNA的3’-OH 和5’-P形成磷酸二酯键。DNA末端可以是粘末端,也可以是平末端。粘末端连接反应效率高于平末端连接效率。因此,在连接平末端分子时DNA 浓度要高于粘末端分子连接反应的DNA浓度。PEG可以使T4 DNA连接酶对平末端和粘末端连接效率得到提高[Pheiffer, B.H., Zimmerman, S.B., Nucleic Acids Res., 11, 7853-7871, 1983]。由于PEG可以辅助提高连接效率,该试剂盒对于粘末端和平末端的连接只需十分钟即可完成。试剂盒中提供独特的5×Rapid Ligation Buffe,专为提高DNA连接效率设计。高的PEG浓度可以提高连接反应效率但却会使转化效率降低,因此应该对PEG浓度进行优化。该试剂盒中优化的PEG浓度同时保证了高的连接效率和转化效率。 目录号规格价格 LINK-3030次180元 保存: 试剂盒中所有组分需-20℃保存。请不要用其它试剂替代该试剂盒中的反应缓冲液。 试剂盒包装组成: 该试剂盒中提供的试剂可以完成30×20μl DNA连接反应。 试剂盒组成规格( 30 reactions ) T4 DNA ligase 30μl 5×Rapid Ligation Buffer 200μl (5x RLB) 特点: 室温(20-25℃)连接10-30 min。 存储缓冲液: 10 mM Tris-HCl (pH 7.5) 50 mM KCl 1 mM Dithiothreitol 50% Glycerol 质量控制: 试剂盒中提供的T4 DNA连接酶没有检测到外切核酸酶或内切核酸酶活性。另外每一批连接酶都通过SDS聚丙烯酰胺凝胶检测蛋白污染(低于5%)。 ·核酸外切酶污染检验 T4 DNA连接酶与1mg经超声波处理的3H标记的E.coli DNA (105cpm/mg)在50ml反应体系中,采用随酶提供的Rapid Ligation Buffer,
一. 重组质粒的构建T质粒载体 重组的DNA分子是在DNA连接酶的作用下,有Mg2 、ATP存在的连接缓冲系统中,将分别经酶切的载体分子与外源DNA分子进行连接。 DNA连接酶有两种:T4噬菌体DNA连接酶和大肠杆菌DNA连接酶。两种DNA连接酶都有将两个带有相同粘性末端的DNA分子连在一起的功能,而且T4噬菌体DNA连接酶还有一种大肠杆菌DNA连接酶没有的特性,即能使两个平末端的双链DNA分子连接起来。但这种连接的效率比粘性末端的连接率低,一般可通过提高T4噬菌体DNA 连接酶浓度或增加DNA浓度来提高平末端的连接效率。 T4噬菌体DNA 连接酶催化DNA 连接反应分为3 步:首先,T4 DNA 连接酶与辅因子ATP形成酶-ATP复合物;然后,酶-ATP复合物再结合到具有5’磷酸基和3’羟基切口的DNA上,使DNA腺苷化;最后产生一个新的磷酸二酯键,把切口封起来。连接反应通常将两个不同大小的片断相连。 很多DNA聚合酶在进行PCR扩增时会在PCR产物双链DNA每条链的3’端加上一个突出的碱基A。pUCm-T载体是一种已经线性化的载体,载体每条链的3’端带有一个突出的T。这样,pUCm-T载体的两端就可以和PCR产物的两端进行正确的AT配对,在连接酶的催化下,就可以把PCR产物连接到pUCm-T载体中,形成含有目的片断的重组载体。 连接反应的温度在37℃时有利于连接酶的活性。但是在这个温度下粘末端的氢键结合是不稳定的。因此采取折中的温度,即12-16℃,连接12-16h(过夜),这样既可最大限度地发挥连接酶的活性,又兼顾到短暂配对结构的稳定。 二. 感受态制备原理 低渗溶液中胀成球形,丢失部分膜蛋白,成为容易细菌在0°C CaCl 2 吸收外源DNA的感受态。 三. β-半乳糖甘酶显色反应选择法 LacZ基因是大肠杆菌乳糖操纵子中的一个基因,可以编码β—半乳糖核苷酶。β—半乳糖核苷酶是由4个亚基组成的四聚体,可催化乳糖的水解.用X-Gal为底物进行染色时,呈蓝色。 现在一些特定的质粒(比如pUC/pBS等),常带有β—半乳糖核苷酶的调控序列和β—半乳糖核苷酶N端146个氨基酸(α肽段)的编码序列,在这个编码序列里还插入一个多克隆位点(MCS),它并不影响lacZ的表达。另外,常用的大肠杆菌带有β—半乳糖核苷酶C端部分序列(β肽段),的编码序列。在各自独立的情况下,这些质粒与大肠杆菌各自编码的β—半乳糖核苷酶片段都没有酶的活性。只有当携带α肽编码信息的克隆载体成功进入宿主细胞,在培养基诱导物IPTG的诱导下,载体质粒能够合成β—半乳糖核苷酶N端(α肽段),这样就与宿主细胞合成的β—半乳糖核苷酶C端部分序列(β肽段)互补,形成完整的β—半乳糖核苷酶活性蛋白。